
Effect of Copper on Expression of Functional Genes and Proteins Associated with Bradyrhizobium diazoefficiens Denitrification

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
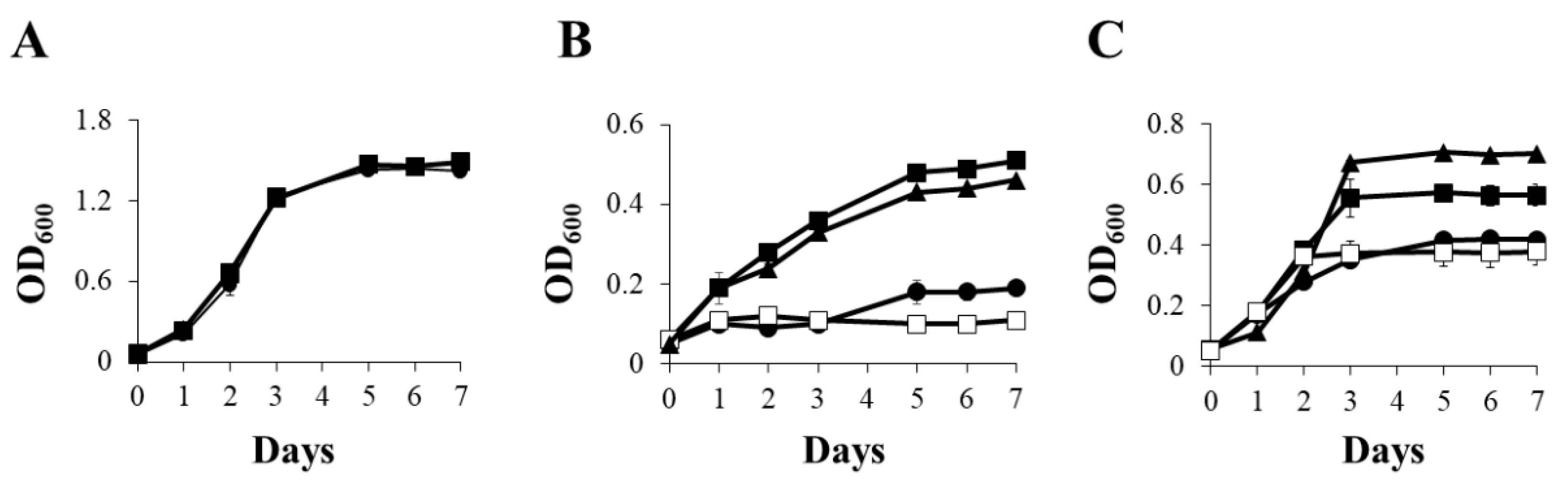
2.1. Copper Effect on B. diazoefficiens 110spc4 Growth under Different Oxygen Conditions
2.2. Disparate Response of Denitrification Gene Expression to Copper
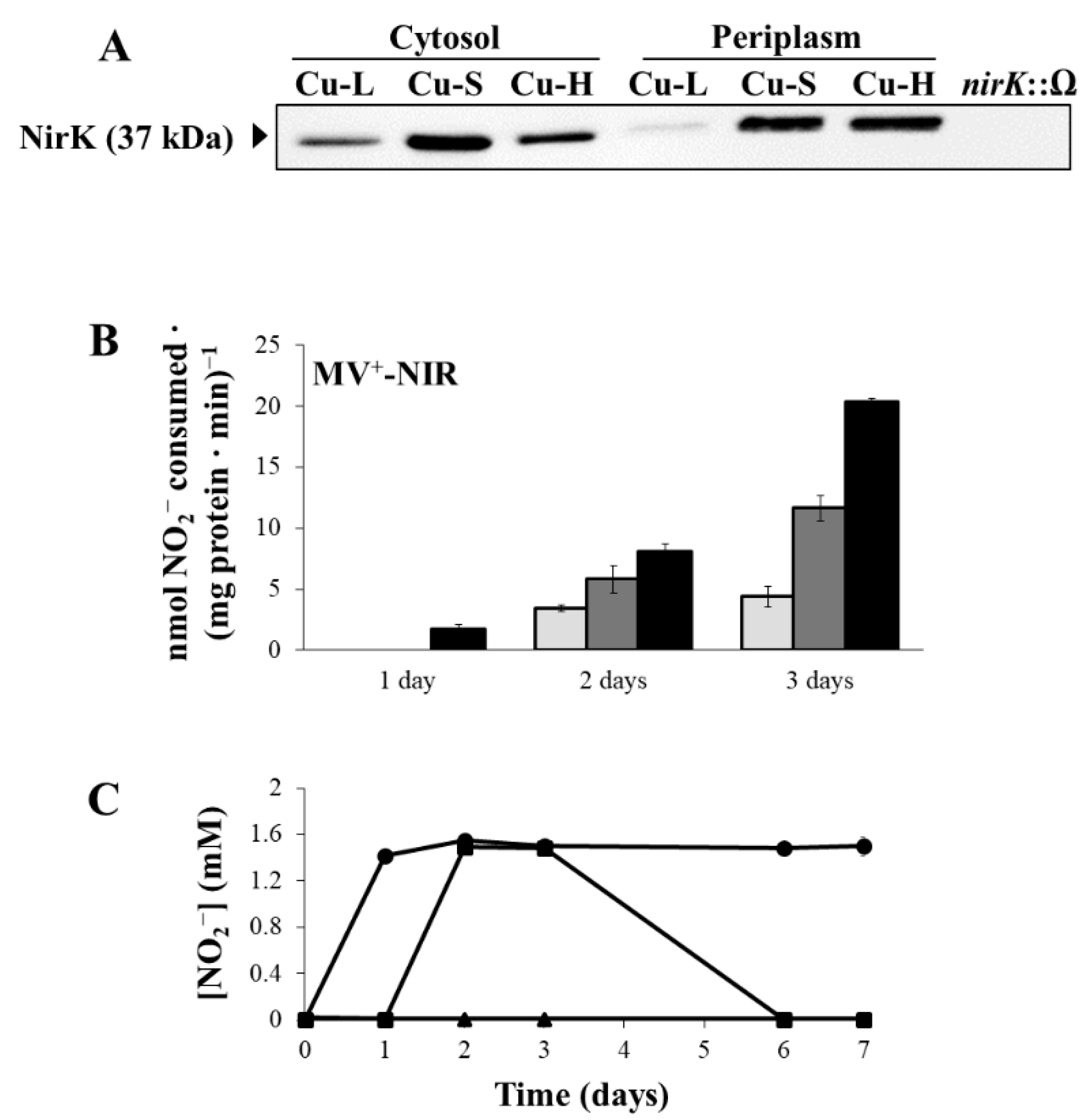
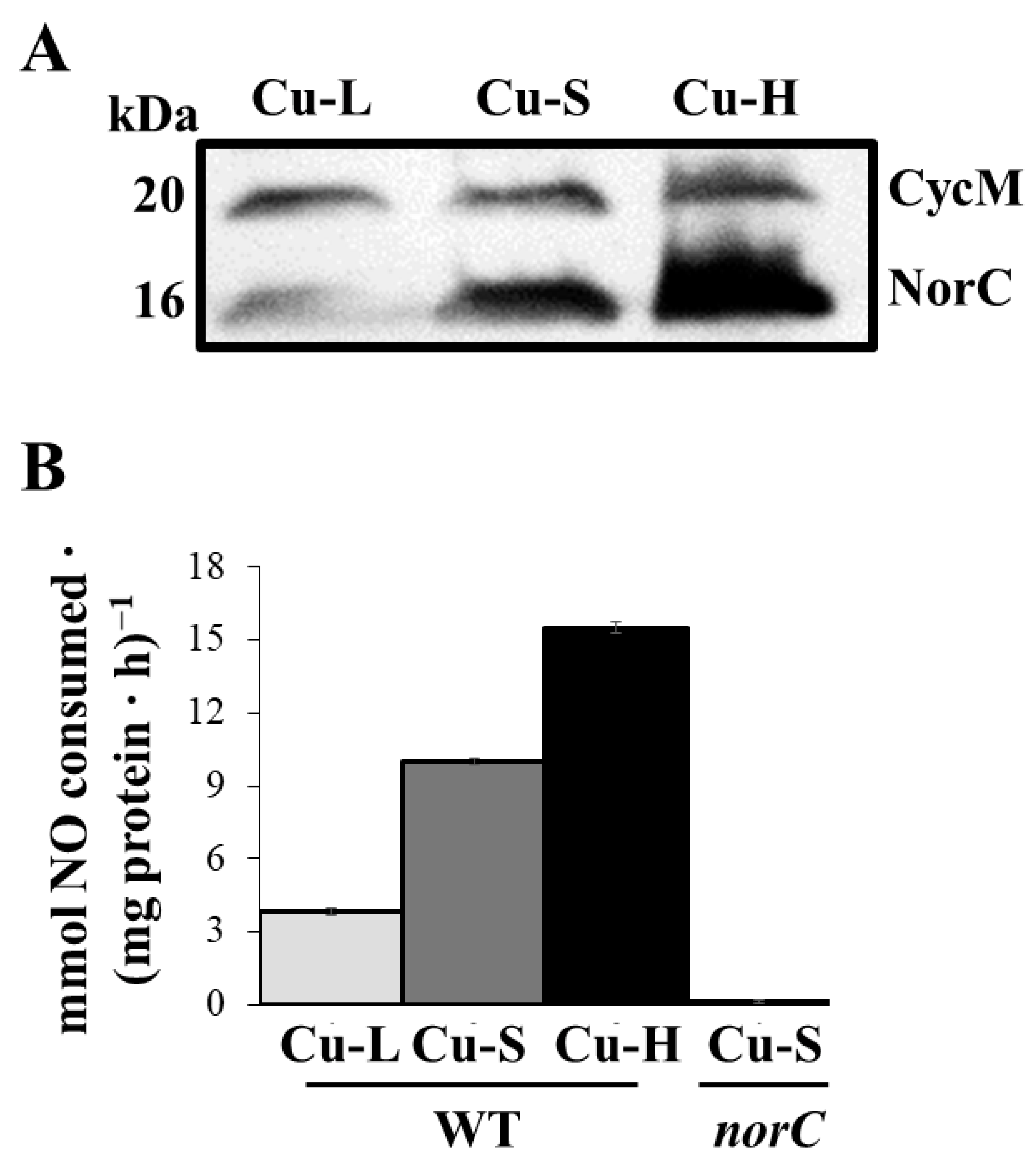
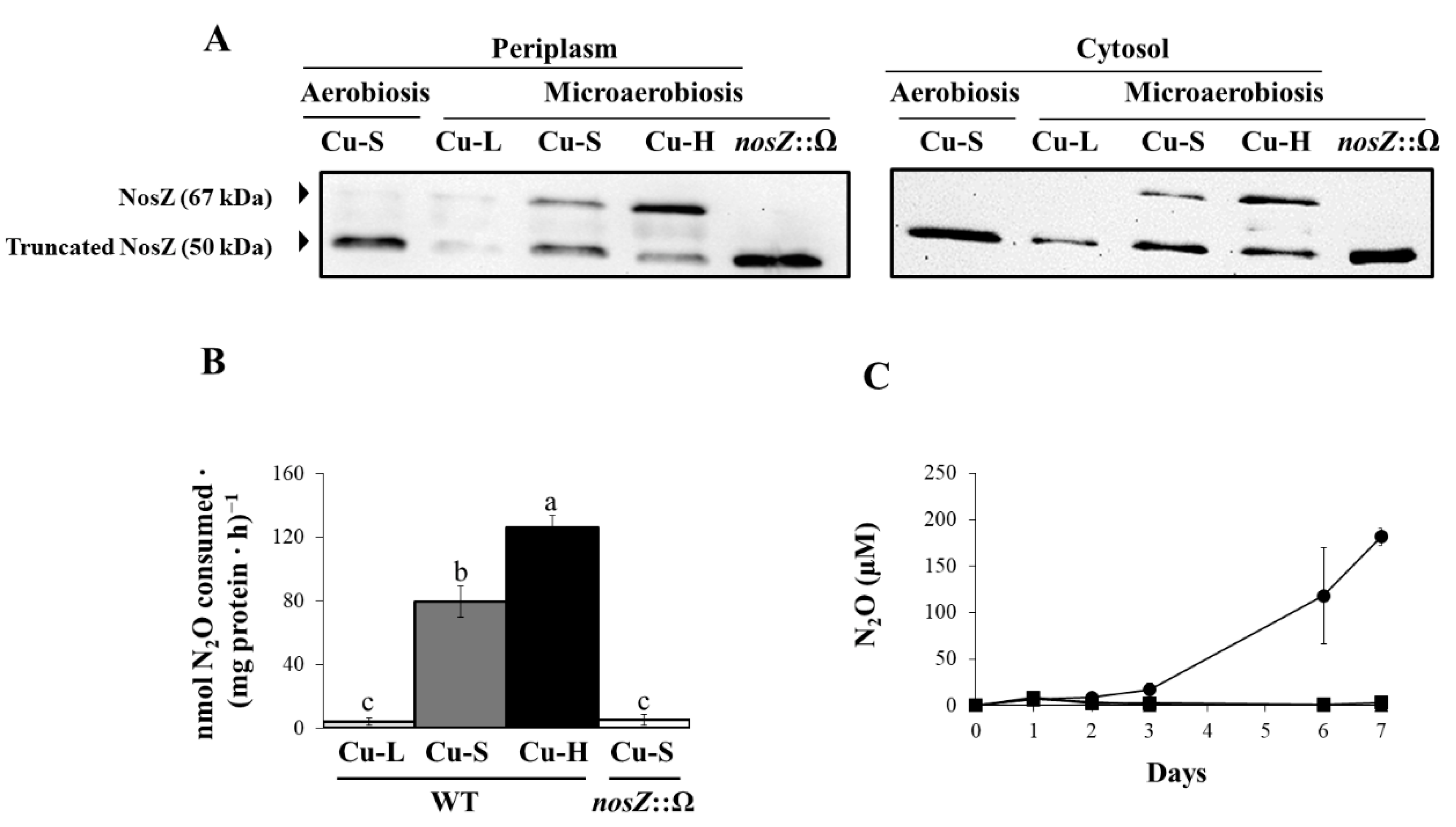
2.3. Influence of Copper on Expression and Activity of Denitrification Enzymes
2.4. Investigating the Possible Role of NosR in Nos Gene Expression and Nos Activity
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Analysis of Gene Expression by qRT-PCR
4.3. Analytical Methods
4.4. Determination of β-Galactosidase Activity
4.5. Determination of N2O Production
4.6. Determination of Nitrate (NR, EC 1.7.99.4) and Nitrite Reductase (NIR, EC 1.7.2.1) Activities
4.7. Determination of Nitric Oxide Reductase (NOR, EC 1.7.2.5) Activity
4.8. Determination of Nitrous Oxide Reductase (N2OR, EC 1.7.2.4) Activity
4.9. Detection of Membrane-Bound NorC by Heme-Staining
4.10. Detection of NapA, NirK and NosZ by Immunoblot Analyses
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ravishankara, A.R.; Daniel, J.S.; Portmann, R.W. Nitrous oxide (N2O): The dominant ozone-depleting substance emitted in the 21st century. Science 2009, 326, 123–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IPCC. Summary for policymakers. In Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; Shukla, P.R., Skea, J., Calvo Buendía, E., Masson-Delmotte, V., Pörtner, H.-O., Roberts, D.C., Zhai, P., Slade, R., Connors, S., van Diemen, R., et al., Eds.; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2019. [Google Scholar]
- Galloway, J.N.; Aber, J.D.; Erisman, J.W.; Seitzinger, S.P.; Howarth, R.W.; Cowling, E.B.; Cosby, B.J. The nitrogen cascade. Bioscience 2003, 53, 341–356. [Google Scholar] [CrossRef]
- Richardson, D.; Felgate, H.; Watmough, N.; Thomson, A.; Baggs, E. Mitigating release of the potent greenhouse gas N2O from the nitrogen cycle—could enzymic regulation hold the key? Trends. Biotechnol. 2009, 27, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.G.; Townsend, A.R. Stoichiometric control of organic carbon-nitrate relationships from soils to the sea. Nature 2010, 464, 1178–1181. [Google Scholar] [CrossRef] [PubMed]
- Thomson, A.J.; Giannopulos, G.; Pretty, J.; Baggs, E.M.; Richardson, D.J. Biological sources and sinks of nitrous oxide and strategies to mitigate emissions. Philos. Trans. R. Soc. 2012, 367, 1157–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapleigh, J.P. The denitrifying prokaryotes. In The Prokaryotes; Dworkin, M., Ed.; Springer Science: New York, NY, USA, 2006; Volume 2, pp. 769–793. [Google Scholar] [CrossRef]
- Zumft, W.G. Cell biology and molecular basis of denitrification. Microbiol. Mol. Biol. Rev. 1997, 61, 533–616. [Google Scholar] [CrossRef]
- van Spanning, R.J.M.; Delgado, M.J.; Richardson, D.J. The nitrogen cycle: Denitrification and its relationship to N2 fixation. In Nitrogen Fixation in Agriculture, Forestry, Ecology, and the Environment; Werner, D., Newton, W.E., Eds.; Spring Science: Dordrecht, The Netherlands, 2005; Volume 4, pp. 277–327. [Google Scholar]
- van Spanning, R.J.M.; Richardson, D.J.; Ferguson, S.J. Introduction to the biochemistry and molecular biology of denitrification. In Biology of the Nitrogen Cycle, 1st ed.; Bothe, H., Ferguson, S.J., Newton, W.E., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 3–20. [Google Scholar]
- Kraft, B.; Strous, M.; Tegetmeyer, H.E. Microbial nitrate respiration--genes, enzymes and environmental distribution. J. Biotechnol. 2011, 155, 104–117. [Google Scholar] [CrossRef]
- Richardson, D.J. Redox complexes of the nitrogen cycle. In Nitrogen Cycling in Bacteria: Molecular Analysis; Moir, J.W.B., Ed.; Caister Academic Press: Norfolk, UK, 2011; pp. 23–39. [Google Scholar] [CrossRef]
- Bueno, E.; Mesa, S.; Bedmar, E.J.; Richardson, D.J.; Delgado, M.J. Bacterial adaptation of respiration from oxic to microoxic and anoxic conditions: Redox control. Antioxid. Redox. Signal 2012, 16, 819–852. [Google Scholar] [CrossRef]
- Torres, M.J.; Simon, J.; Rowley, G.; Bedmar, E.J.; Richardson, D.J.; Gates, A.J.; Delgado, M.J. Nitrous Oxide Metabolism in Nitrate-Reducing Bacteria: Physiology and Regulatory Mechanisms. Adv. Microb. Physiol. 2016, 68, 353–432. [Google Scholar] [CrossRef] [Green Version]
- Bedmar, E.J.; Robles, E.F.; Delgado, M.J. The complete denitrification pathway of the symbiotic, nitrogen-fixing bacterium Bradyrhizobium japonicum. Biochem. Soc. Trans. 2005, 33, 141–144. [Google Scholar] [CrossRef] [Green Version]
- Bedmar, E.J.; Bueno, E.; Correa, D.; Torres, M.J.; Delgado, M.J.; Mesa, S. Ecology of denitrification in soils and plant-associated bacteria. In Beneficial Plant-Microbial Interactions: Ecology and Applications, 1st ed.; Rodelas, B., González-López, J., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 164–182. [Google Scholar] [CrossRef]
- Salas, A.; Cabrera, J.J.; Jiménez-Leiva, A.; Mesa, S.; Bedmar, E.J.; Richardson, D.J.; Gates, A.J.; Delgado, M.J. Bacterial nitric oxide metabolism: Recent insights in rhizobia. Adv. Microb. Physiol. 2021, 78, 259–315. [Google Scholar] [CrossRef] [PubMed]
- Poole, P.; Ramachandran, V.; Terpolilli, J. Rhizobia: From saprophytes to endosymbionts. Nat. Rev. Microbiol. 2018, 16, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.J.; Bonnard, N.; Tresierra-Ayala, A.; Bedmar, E.J.; Müller, P. The Bradyrhizobium japonicum napEDABC genes encoding the periplasmic nitrate reductase are essential for nitrate respiration. Microbiology 2003, 149, 3395–3403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velasco, L.; Mesa, S.; Delgado, M.J.; Bedmar, E.J. Characterization of the nirK gene encoding the respiratory, Cu-containing nitrite reductase of Bradyrhizobium japonicum. Biochim. Biophys. Acta 2001, 1521, 130–134. [Google Scholar] [CrossRef]
- Mesa, S.; Velasco, L.; Manzanera, M.E.; Delgado, M.J.; Bedmar, E.J. Characterization of the norCBQD genes, encoding nitric oxide reductase, in the nitrogen fixing bacterium Bradyrhizobium japonicum. Microbiology 2002, 148, 3553–3560. [Google Scholar] [CrossRef] [Green Version]
- Velasco, L.; Mesa, S.; Xu, C.; Delgado, M.J.; Bedmar, E.J. Molecular characterization of nosRZDFYLX genes coding for denitrifying nitrous oxide reductase of Bradyrhizobium japonicum. Antonie Leeuwenhoek 2004, 85, 229–235. [Google Scholar] [CrossRef]
- Mesa, S.; Bedmar, E.J.; Chanfon, A.; Hennecke, H.; Fischer, H.-M. Bradyrhizobium japonicum NnrR, a denitrification regulator, expands the FixLJ-FixK2 regulatory cascade. J. Bacteriol. 2003, 185, 3978–3982. [Google Scholar] [CrossRef] [Green Version]
- Mesa, S.; Hauser, F.; Friberg, M.; Malaguti, E.; Fischer, H.-M.; Hennecke, H. Comprehensive assessment of the regulons controlled by the FixLJ-FixK2-FixK1 cascade in Bradyrhizobium japonicum. J. Bacteriol. 2008, 190, 6568–6579. [Google Scholar] [CrossRef] [Green Version]
- Bueno, E.; Robles, E.F.; Torres, M.J.; Krell, T.; Bedmar, E.J.; Delgado, M.J.; Mesa, S. Disparate response to microoxia and nitrogen oxides of the Bradyrhizobium japonicum napEDABC, nirK and norCBQD denitrification genes. Nitric Oxide 2017, 68, 137–149. [Google Scholar] [CrossRef] [Green Version]
- Torres, M.J.; Bueno, E.; Jiménez-Leiva, A.; Cabrera, J.J.; Bedmar, E.J.; Mesa, S.; Delgado, M.J. FixK2 Is the main transcriptional activator of Bradyrhizobium diazoefficiens nosRZDYFLX genes in response to low oxygen. Front. Microbiol. 2017, 8, 1621. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Leiva, A.; Cabrera, J.J.; Bueno, E.; Torres, M.J.; Salazar, S.; Bedmar, E.J.; Delgado, M.J.; Mesa, S. Expanding the regulon of the Bradyrhizobium diazoefficiens NnrR transcription factor: New insights into the denitrification pathway. Front. Microbiol. 2019, 10, 1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera, J.J.; Jiménez-Leiva, A.; Tomás-Gallardo, L.; Parejo, S.; Casado, S.; Torres, M.J.; Bedmar, E.J.; Delgado, M.J.; Mesa, S. Dissection of FixK2 protein-DNA interaction unveils new insights into Bradyrhizobium diazoefficiens lifestyles control. Environ. Microbiol. 2021, 23, 6194–6209. [Google Scholar] [CrossRef] [PubMed]
- Bakken, L.R.; Frostegard, A. Sources and sinks for N2O, can microbiologist help to mitigate N2O emissions? Environ. Microbiol. 2017, 19, 4801–4805. [Google Scholar] [CrossRef]
- Carreira, C.; Nunes, R.F.; Mestre, O.; Moura, I.; Pauleta, S.R. The effect of pH on Marinobacter hydrocarbonoclasticus denitrification pathway and nitrous oxide reductase. J. Biol. Inorg. Chem. 2020, 25, 927–940. [Google Scholar] [CrossRef] [PubMed]
- Olaya-Abril, A.; Hidalgo-Carrillo, J.; Luque-Almagro, V.M.; Fuentes-Almagro, C.; Urbano, F.J.; Moreno-Vivián, C.; Richardson, D.J.; Roldán, M.D. Effect of pH on the denitrification proteome of the soil bacterium Paracoccus denitrificans PD1222. Sci. Rep. 2021, 11, 17276. [Google Scholar] [CrossRef] [PubMed]
- Black, A.; Hsu, P.-C.L.; Hamonts, K.E.; Clough, T.J.; Condron, L.M. Influence of copper on expression of nirS, norB and nosZ and the transcription and activity of NIR, NOR and N2OR in the denitrifying soil bacteria Pseudomonas stutzeri. Microb. Biotechnol. 2016, 9, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Matsubara, T.; Frunzke, K.; Zumft, W.G. Modulation by copper of the products of nitrite respiration in Pseudomonas perfectomarinus. J. Bacteriol. 1982, 149, 816–823. [Google Scholar] [CrossRef] [Green Version]
- Felgate, H.; Giannopulos, G.; Sullivan, M.J.; Gates, A.J.; Clarke, T.A.; Baggs, E.; Rowley, G.; Richardson, D.J. The impact of copper, nitrate and carbon status on the emission of nitrous oxide by two species of bacteria with biochemically distinct denitrification pathways. Environ. Microbiol. 2012, 14, 1788–1800. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Gates, A.J.; Appia-Ayme, C.; Rowley, G.; Richardson, D.J. Copper control of bacterial nitrous oxide emission and its impact on vitamin B12-dependent metabolism. Proc. Natl. Acad. Sci. USA 2013, 110, 19926–19931. [Google Scholar] [CrossRef] [Green Version]
- Serventi, F.; Youard, Z.A.; Murset, V.; Huwiler, S.; Bühler, D.; Richter, M.; Luchsinger, R.; Fischer, H.-M.; Brogioli, R.; Niederer, M.; et al. Copper Starvation-inducible Protein for Cytochrome Oxidase Biogenesis in Bradyrhizobium japonicum. J. Biol. Chem. 2012, 287, 38812–38823. [Google Scholar] [CrossRef] [Green Version]
- Richardson, D.J.; Wehrfritz, J.-M.; Keech, A.; Crossman, L.C.; Roldan, M.D.; Sears, H.J.; Butler, C.S.; Reilly, A.; Moir, J.W.B.; Berks, B.C.; et al. The diversity of redox proteins involved in bacterial heterotrophic nitrification and aerobic denitrification. Biochem. Soc. Trans. 1998, 26, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Bueno, E.; Bedmar, E.J.; Richardson, D.J.; Delgado, M.J. Role of Bradyrhizobium japonicum cytochrome c550 in nitrite and nitrate respiration. FEMS Microbiol. Lett. 2008, 279, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Delgado, M.J.; Bedmar, E.J.; Downie, J.A. Genes involved in the formation and assembly of rhizobial cytochromes and their role in symbiotic nitrogen fixation. Adv. Microb. Physiol. 1998, 40, 191–231. [Google Scholar] [CrossRef] [PubMed]
- Bühler, D.; Rossmann, R.; Landolt, S.; Balsiger, S.; Fischer, H.-M.; Hennecke, H. Disparate pathways for the biogenesis of cytochrome oxidases in Bradyrhizobium japonicum. J. Biol. Chem. 2010, 285, 15704–15713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jünemann, S. Cytochrome bd terminal oxidase. Biochim. Biophys. Acta 1997, 1321, 107–127. [Google Scholar] [CrossRef] [Green Version]
- Marcus, S.A.; Sidiropulos, S.W.; Steinberg, H.; Talaat, A.M. CsoR Is Essential for Maintaining Copper Homeostasis in Mycobacterium tuberculosis. PLoS ONE 2016, 11, e0151816. [Google Scholar] [CrossRef]
- Liu, T.; Ramesh, A.; Ma, Z.; Ward, S.K.; Zhang, L.; George, G.N.; Talaat, A.M.; Sacchettini, J.C.; Giedroc, D.P. CsoR is a novel Mycobacterium tuberculosis copper-sensing transcriptional regulator. Nat. Chem. Biol. 2007, 3, 60–68. [Google Scholar] [CrossRef]
- Rademacher, C.; Masepohl, B. Copper-responsive gene regulation in bacteria. Microbiology 2012, 158, 2451–2464. [Google Scholar] [CrossRef]
- Liang, J.; Zhang, M.; Lu, M.; Li, Z.; Shen, X.; Chou, M.; Wei, G. Functional characterization of a csoR-cueA divergon in Bradyrhizobium liaoningense CCNWSX0360, involved in copper, zinc and cadmium cotolerance. Sci. Rep. 2016, 6, 35155. [Google Scholar] [CrossRef] [Green Version]
- Fraústo da Silva, J.J.R.; Williams, R.J.P. The Biological Chemistry of the Elements: The Inorganic Chemistry of Life, 2nd ed.; Oxford University Press Inc.: New York, NY, USA, 2001. [Google Scholar] [CrossRef] [Green Version]
- Maillard, J.; Spronk, C.A.E.M.; Buchanan, G.; Lyall, V.; Richardson, D.J.; Palmer, T.; Vuister, G.W.; Sargent, F. Structural diversity in twin-arginine signal peptide-binding proteins. Proc. Natl. Acad. Sci. USA 2007, 104, 15641–15646. [Google Scholar] [CrossRef] [Green Version]
- Sargent, F. The twin-arginine transport system: Moving folded proteins across membranes. Biochem. Soc. T 2007, 35, 835–847. [Google Scholar] [CrossRef] [PubMed]
- Esclapez, J.; Zafrilla, J.; Martínez-Espinosa, R.M.; Bonete, M.J. Cu-NirK from Haloferax mediterranei as an example of metalloprotein maturation and exportation via Tat system. Biochim. Biophys. Acta 2013, 1834, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Cristaldi, J.C.; Ferroni, F.; Duré, A.B.; Ramírez, C.S.; Dalosto, S.D.; Rizzi, A.C.; González, P.J.; Rivas, M.G.; Brondino, C.D. Heterologous production and functional characterization of Bradyrhizobium japonicum copper-containing nitrite reductase and its physiological redox partner cytochrome c550. Metallomics 2020, 12, 2084–2097. [Google Scholar] [CrossRef] [PubMed]
- Granger, J.; Ward, B.B. Accumulation of nitrogen oxides in copper-limited cultures of denitrifying bacteria. Limnol. Oceanogr. 2003, 48, 313–318. [Google Scholar] [CrossRef]
- Wüst, A.; Schneider, L.; Pomowski, A.; Zumft, W.G.; Kroneck, P.M.H.; Einsle, O. Nature’s way of handling a greenhouse gas: The copper-sulfur cluster of purple nitrous oxide reductase. Biol. Chem. 2012, 393, 1067–1077. [Google Scholar] [CrossRef]
- Wunsch, P.; Zumft, W.G. Functional domains of NosR, a novel transmembrane iron-sulfur flavoprotein necessary for nitrous oxide respiration. J. Bacteriol. 2005, 187, 1992–2001. [Google Scholar] [CrossRef] [Green Version]
- Zumft, W.G.; Kroneck, P.M.H. Respiratory transformation of nitrous oxide (N2O) to dinitrogen by Bacteria and Archaea. Adv. Microb. Physiol. 2007, 52, 107–227. [Google Scholar] [CrossRef]
- Wunsch, P.; Körner, H.; Neese, F.; van Spanning, R.J.M.; Kroneck, P.M.H.; Zumft, W.G. NosX function connects to nitrous oxide (N2O) reduction by affecting the Cuz center of NosZ and its activity in vivo. FEBS Lett. 2005, 579, 4605–4609. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Trncik, C.; Andrade, S.L.A.; Einsle, O. The flavinyl transferase ApbE of Pseudomonas stutzeri matures the NosR protein required for nitrous oxide reduction. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 95–102. [Google Scholar] [CrossRef]
- Honisch, U.; Zumft, W.G. Operon structure and regulation of the nos gene region of Pseudomonas stutzeri, encoding an ABC-Type ATPase for maturation of nitrous oxide reductase. J. Bacteriol. 2003, 185, 1895–1902. [Google Scholar] [CrossRef] [Green Version]
- Vincent, J.M. A Manual for Practical Study of Root Nodule Bacteria; Blackwell Scientific Publications: Oxford, UK, 1970. [Google Scholar]
- Becker, A.; Bergès, H.; Krol, E.; Bruand, C.; Rüberg, S.; Capela, D.; Lauber, E.; Meilhoc, E.; Ampe, F.; de Bruijn, F.J.; et al. Global changes in gene expression in Sinorhizobium meliloti 1021 under microoxic and symbiotic conditions. Mol. Plant Microbe Interact. 2004, 17, 292–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishop, P.E.; Guevara, J.G.; Engelke, J.A.; Evans, H.J. Relation between Glutamine-Synthetase and Nitrogenase Activities in Symbiotic Association between Rhizobium japonicum and Glycine max. Plant Physiol. 1976, 57, 542–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regensburger, B.; Hennecke, H. RNA polymerase from Rhizobium japonicum. Arch. Microbiol. 1983, 135, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Robles, E.F.; Sánchez, C.; Bonnard, N.; Delgado, M.J.; Bedmar, E.J. The Bradyrhizobium japonicum napEDABC genes are controlled by the FixLJ-FixK2-NnrR regulatory cascade. Biochem. Soc. Trans. 2006, 34, 108–110. [Google Scholar] [CrossRef]
- Hauser, F.; Pessi, G.; Friberg, M.; Weber, C.; Rusca, N.; Lindemann, A.; Fischer, H.-M.; Hennecke, H. Dissection of the Bradyrhizobium japonicum NifA+σ54 regulon, and identification of a ferredoxin gene (fdxN) for symbiotic nitrogen fixation. Mol. Genet. Genom. 2007, 278, 255–271. [Google Scholar] [CrossRef]
- Lindemann, A.; Moser, A.; Pessi, G.; Hauser, F.; Friberg, M.; Hennecke, H.; Fischer, H.-M. New target genes controlled by the Bradyrhizobium japonicum two-component regulatory system RegSR. J. Bacteriol. 2007, 189, 8928–8943. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef]
- Hageman, R.H.; Hucklesby, D.P. Nitrate reductase from higher plants. In Methods in Enzymology; San Pietro, A., Ed.; Academic Press: London, UK, 1971; Volume 23, pp. 491–503. [Google Scholar] [CrossRef]
- Miranda, K.M.; Espey, M.G.; Wink, D.A. A rapid, simple spectrophotometric method for simultaneous detection of nitrate and nitrite. Nitric Oxide 2001, 5, 62–71. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for Quantitation of Microgram Quantities of Protein Utilizing Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cabrera, J.J.; Salas, A.; Torres, M.J.; Bedmar, E.J.; Richardson, D.J.; Gates, A.J.; Delgado, M.J. An integrated biochemical system for nitrate assimilation and nitric oxide detoxification in Bradyrhizobium japonicum. Biochem. J. 2016, 473, 297–309. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.H. Experiments in Molecular Genetics, 1st ed.; Cold Spring Harbor: Long Island, NY, USA, 1972. [Google Scholar]
- Bricio, C.; Alvarez, L.; San Martin, M.; Schurig-Briccio, L.A.; Gennis, R.B.; Berenguer, J. A Third Subunit in Ancestral Cytochrome c-Dependent Nitric Oxide Reductases. Appl. Environ. Microbiol. 2014, 80, 4871–4878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, M.J.; Rubia, M.I.; de la Peña, T.C.; Pueyo, J.J.; Bedmar, E.J.; Delgado, M.J. Genetic basis for denitrification in Ensifer meliloti. BMC Microbiol. 2014, 14, 142–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Relevant Description | Source of Reference |
|---|---|---|
| 110spc4 | Cmr Spcr wild-type, a spectinomycin resistant derivative of USDA110 | [61] |
| GRPA1 | Cmr Spcr Smr napA::Ω | [19] |
| GRK308 | Cmr Spcr Smr nirK::Ω | [20] |
| GRC131 | Cmr Kmr norC::aphII-PSP | [21] |
| GRZ3035 | Cmr Spcr Smr nosZ::Ω | [22] |
| ΔnosR | Cmr Spcr B. diazoefficiens 110spc4 markerless deletion mutant | Laboratory collection |
| BG0602 | Cmr Tcr napE-lacZ chromosomally integrated into USDA110 | [62] |
| RJ2498 | Cmr Spcr Tcr nirK-lacZ chromosomally integrated into 110spc4 | [23] |
| RJ2499 | Cmr Spcr Tcr norC-lacZ chromosomally integrated into 110spc4 | [23] |
| BG0301 | Cmr Spcr Tcr nosR-lacZ chromosomally integrated into 110spc4 | [26] |
| ΔnosR-BG0301 | Cmr Spcr Tcr nosR-lacZ chromosomally integrated into ΔnosR | Laboratory collection |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacheco, P.J.; Cabrera, J.J.; Jiménez-Leiva, A.; Bedmar, E.J.; Mesa, S.; Tortosa, G.; Delgado, M.J. Effect of Copper on Expression of Functional Genes and Proteins Associated with Bradyrhizobium diazoefficiens Denitrification. Int. J. Mol. Sci. 2022, 23, 3386. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063386
Pacheco PJ, Cabrera JJ, Jiménez-Leiva A, Bedmar EJ, Mesa S, Tortosa G, Delgado MJ. Effect of Copper on Expression of Functional Genes and Proteins Associated with Bradyrhizobium diazoefficiens Denitrification. International Journal of Molecular Sciences. 2022; 23(6):3386. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063386
Chicago/Turabian StylePacheco, Pedro J., Juan J. Cabrera, Andrea Jiménez-Leiva, Eulogio J. Bedmar, Socorro Mesa, Germán Tortosa, and María J. Delgado. 2022. "Effect of Copper on Expression of Functional Genes and Proteins Associated with Bradyrhizobium diazoefficiens Denitrification" International Journal of Molecular Sciences 23, no. 6: 3386. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063386