
Nanoscale Calculation of Proton-Induced DNA Damage Using a Chromatin Geometry Model with Geant4-DNA

 ,
,
Abstract
:1. Introduction
2. Results
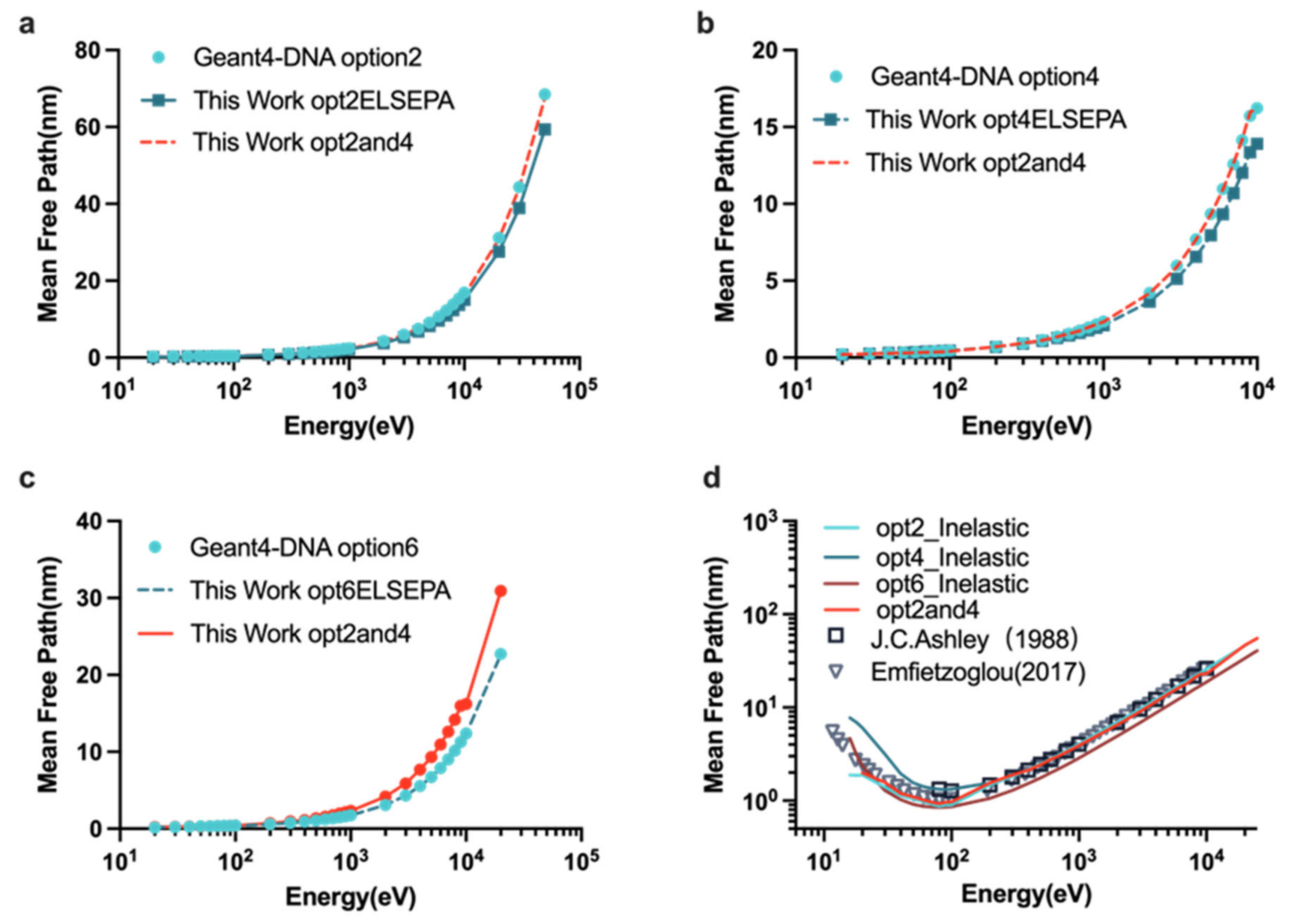
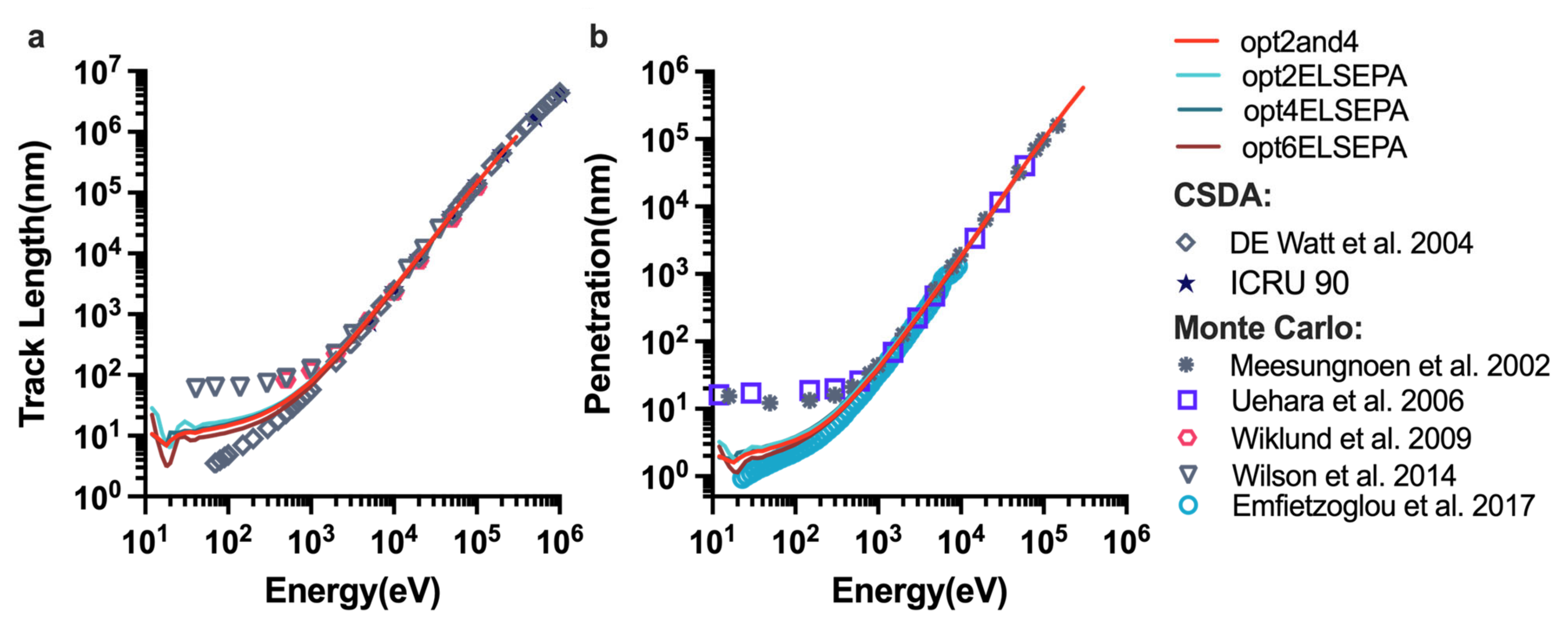
2.1. Benchmarking the Physics Constructors
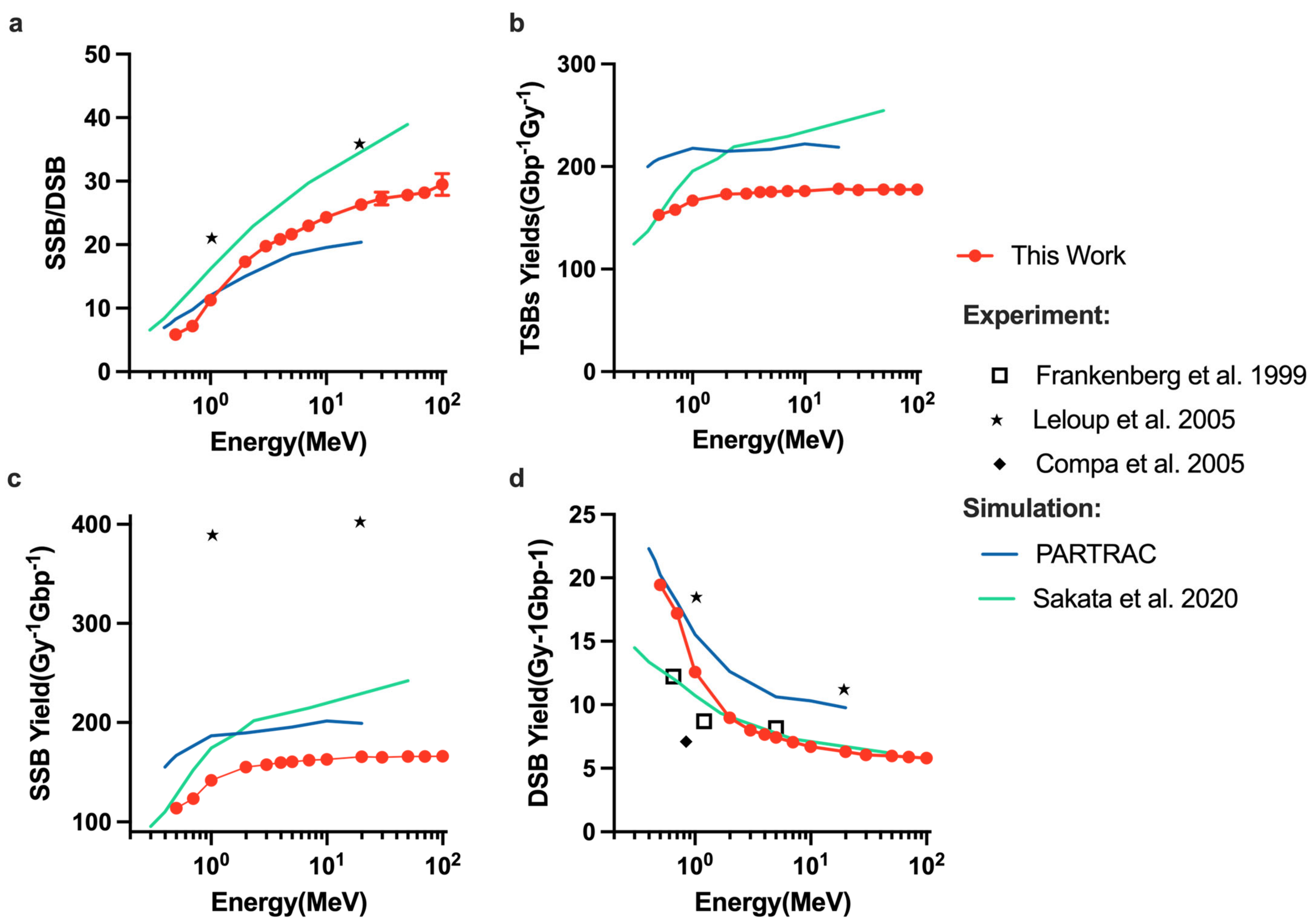
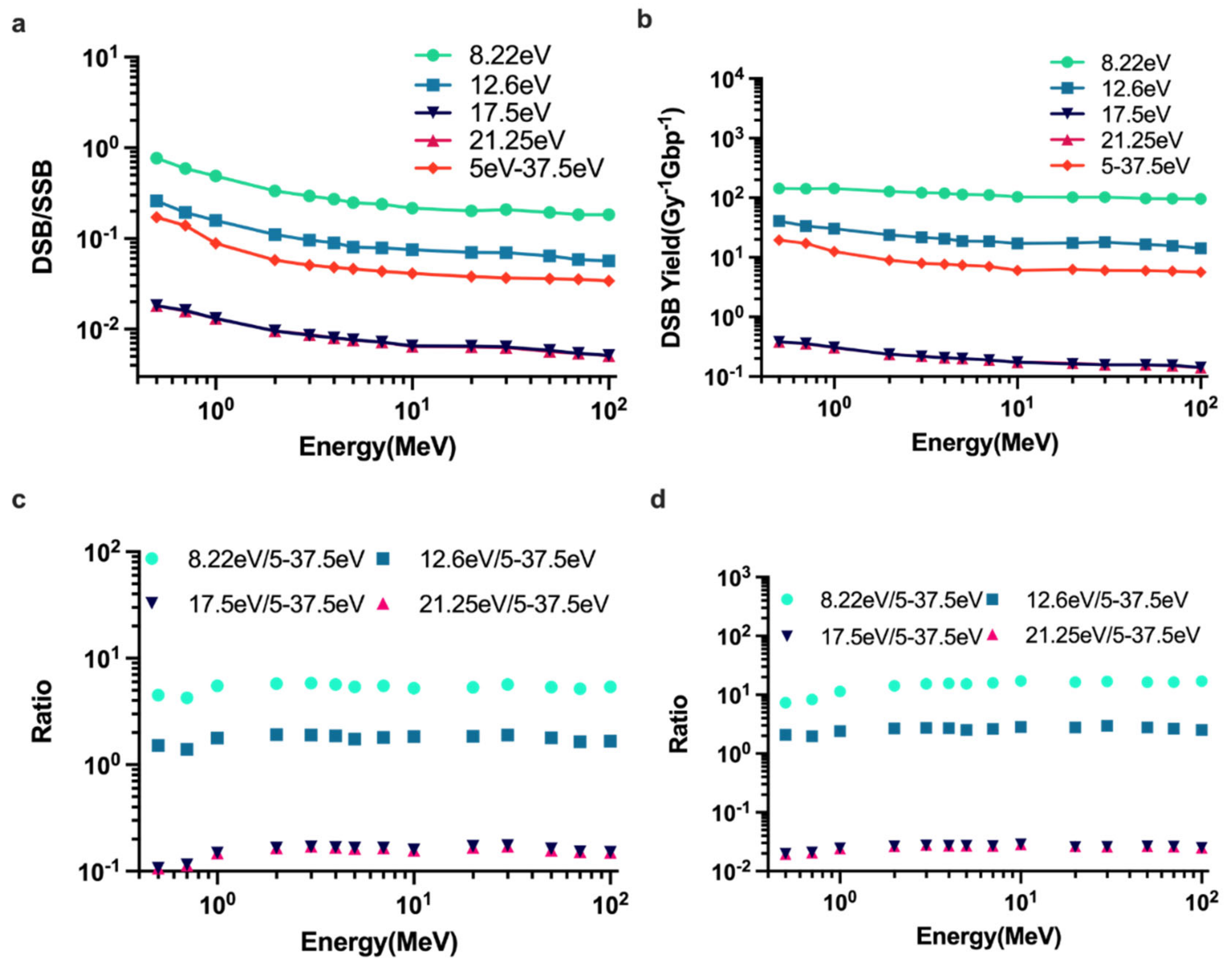
2.2. DNA Damage Simulations
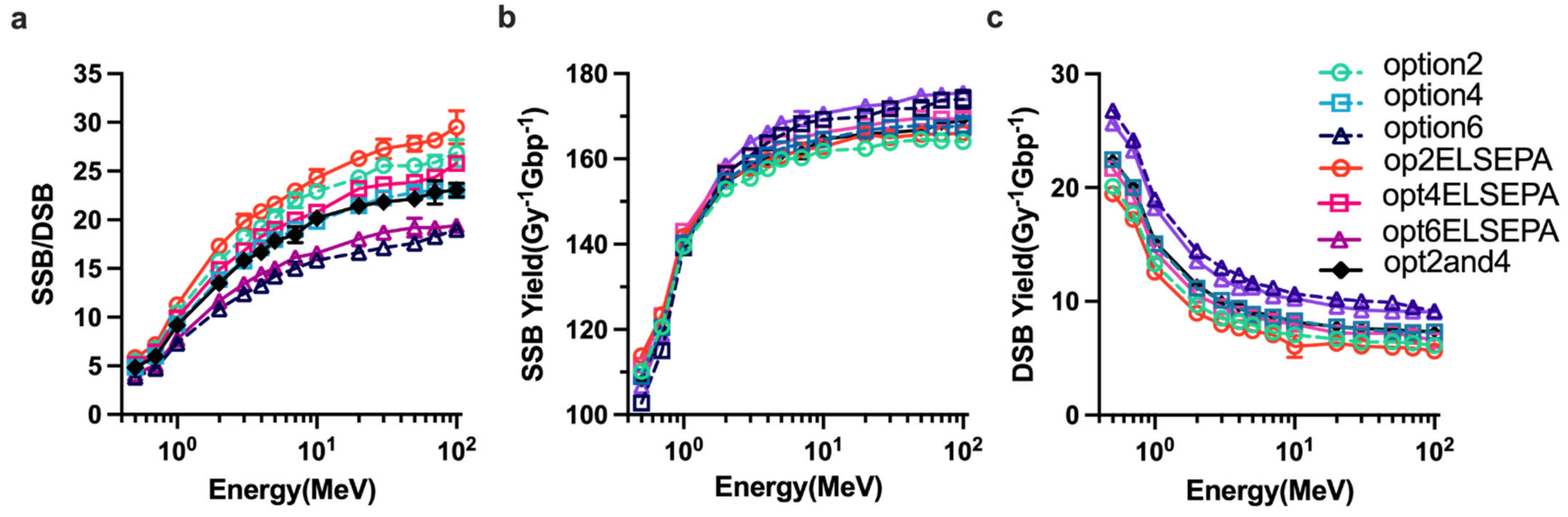
2.3. Parameter Sensitivity Simulations of DNA Damage
3. Discussion
4. Materials and Methods
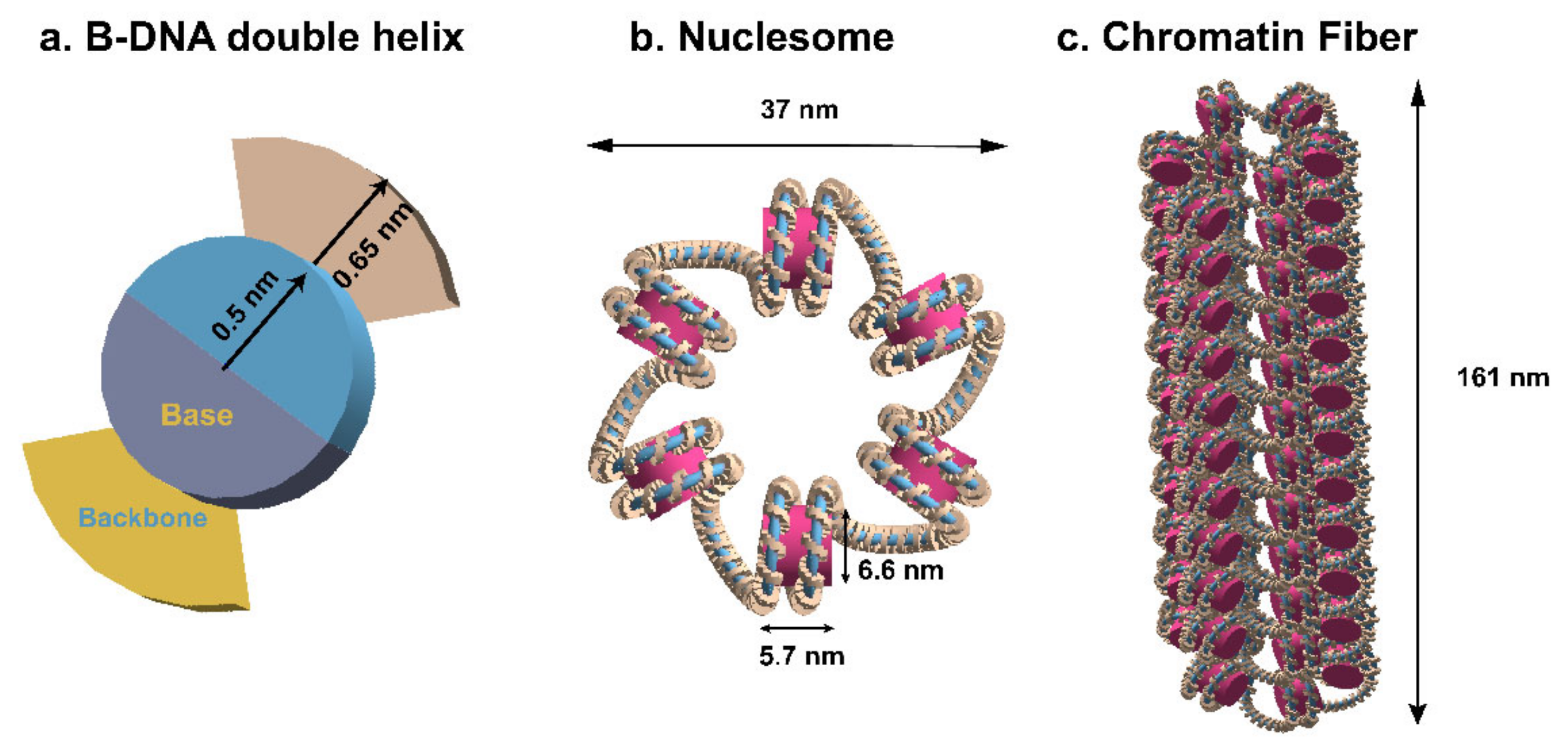
4.1. Models of DNA Geometry
4.2. Physics Settings
4.3. The Examples of Geant4-DNA
4.3.1. The “mfp” Example of Geant4-DNA
4.3.2. The “Range” Example of Geant4-DNA
4.3.3. The “Microyz” Example of Geant4-DNA
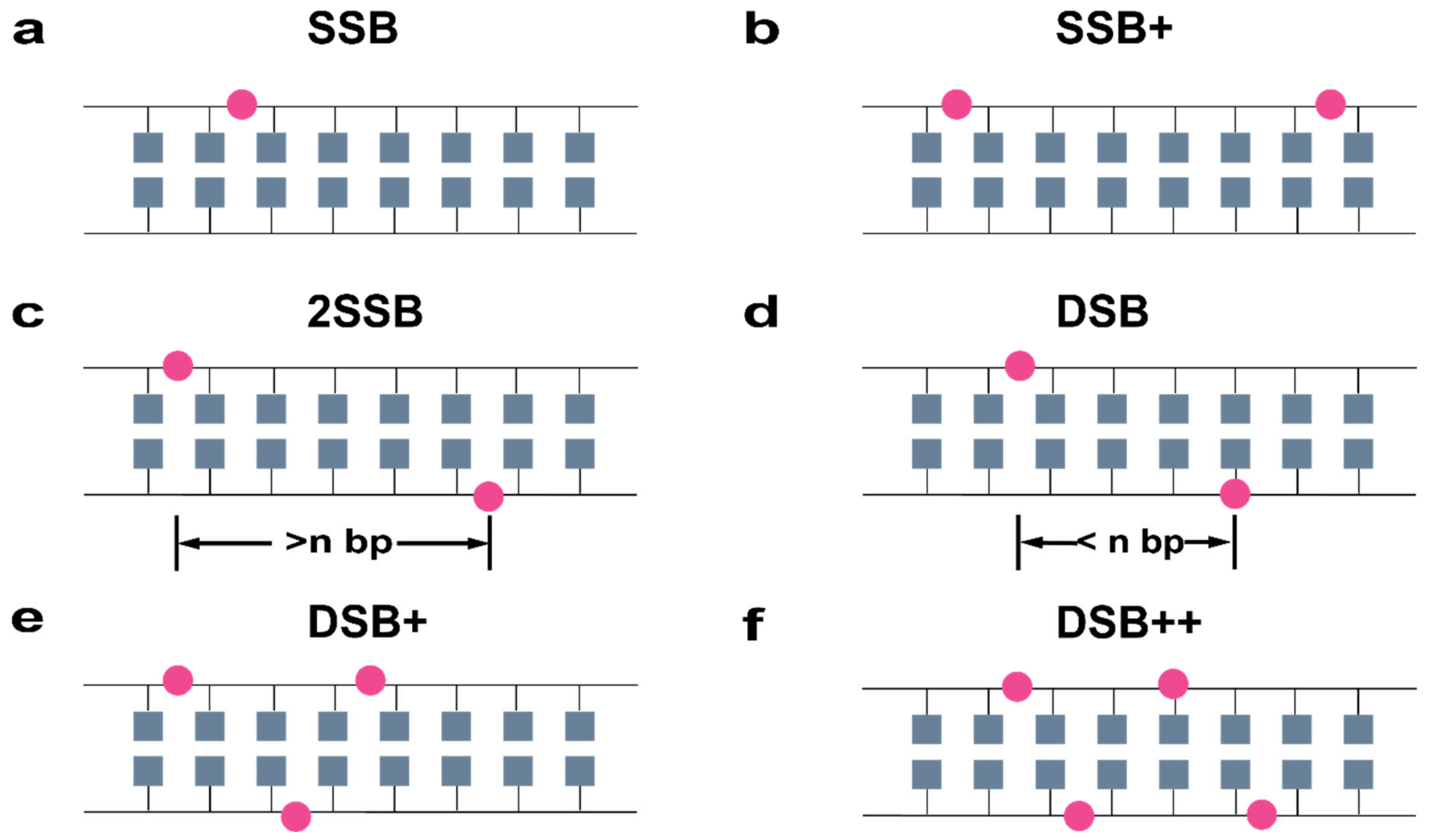
4.4. DNA Damage Model
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IAEAICRU. Relative Biological Effectiveness in Ion Beam Therapy; Technical Reports Series No. 461; International Atomic Energy Agency Vienna International Centre: Vienna, Austria, 2008; pp. 1–165. [Google Scholar]
- Shin, W.; Sakata, D.; Lampe, N.; Belov, O.; Tran, N.H.; Petrovic, I.; Ristic-Fira, A.; Dordevic, M.; Bernal, M.A.; Bordage, M.; et al. A Geant4-DNA evaluation of radiation-induced DNA damage on a human fibroblast. Cancers 2021, 13, 4940. [Google Scholar] [CrossRef] [PubMed]
- Nikjoo, H.; Emfietzoglou, D.; Liamsuwan, T.; Taleei, R.; Liljequist, D.; Uehara, S. Radiation track, DNA damage and response—A review. Rep. Prog. Phys. 2016, 79, 116601. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; McNamara, A.L.; McMahon, S.J.; Mendez, J.R.; Henthorn, N.T.; Faddegon, B.; Held, K.D.; Perl, J.; Li, J.; Paganetti, H.; et al. Cellular response to proton irradiation: A simulation study with TOPAS-nBio. Radiat. Res. 2020, 4, 9–21. [Google Scholar] [CrossRef]
- Nikjoo, H.; Taleei, R.; Liamsuwan, T.; Liljequist, D.; Emfietzoglou, D. Perspectives in radiation biophysics: From radiation track structure simulation to mechanistic models of DNA damage and repair. Radiat. Phys. Chem. 2016, 128, 3–10. [Google Scholar] [CrossRef]
- Vyšín, L.; Brabcová, K.P.; Štěpán, V.; Moretto-Capelle, P.; Bugler, B.; Legube, G.; Cafarelli, P.; Casta, R.; Champeaux, J.P.; Sence, M.; et al. Proton-induced direct and indirect damage of plasmid DNA. Radiat. Environ. Biophys. 2015, 54, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Leloup, C.; Garty, G.; Assaf, G.; Cristovão, A.; Breskin, A.; Chechik, R.; Shchemelinin, S.; Paz-Elizur, T.; Livneh, Z.; Schulte, R.W.; et al. Evaluation of lesion clustering in irradiated plasmid DNA. Int. J. Radiat. Biol. 2005, 81, 41–54. [Google Scholar] [CrossRef] [PubMed]
- Rogakou, E.P.; Pilch, D.R.; Orr, A.H.; Ivanova, V.S.; Bonner, W.M. DNA double-stranded breaks induce histone H2AX phosphorylation on Serine 139. J. Biol. Chem. 1998, 273, 5858–5868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedelnikova, O.A.; Rogakou, E.P.; Panyutin, I.G.; Bonner, W.M. Quantitative detection of 125IdU-induced DNA double-strand breaks with γ-H2AX antibody. Radiat. Res. 2002, 158, 486–492. [Google Scholar] [CrossRef]
- Schipler, A.; Mladenova, V.; Soni, A.; Nikolov, V.; Saha, J.; Mladenov, E.; Iliakis, G. Chromosome thripsis by DNA double strand break clusters causes enhanced cell lethality, chromosomal translocations and 53BP1-recruitment. Nucleic Acids Res. 2016, 44, 7673–7690. [Google Scholar] [CrossRef] [Green Version]
- Ostling, O.; Johanson, K.J. Microelectrophoretic study of radiation-induced DNA damages in individual mammalian cells. Biochem. Biophys. Res. Commun. 1984, 123, 291–298. [Google Scholar] [CrossRef]
- Uehara, S.; Nikjoo, H.; Goodhead, D.T. Cross-sections for water vapour for the Monte Carlo electron track structure code from 10 eV to the MeV region. Phys. Med. Biol. 1993, 38, 1841. [Google Scholar] [CrossRef]
- Nikjoo, H.; Uehara, S.; Emfietzoglou, D.; Pinsky, L. A database of frequency distributions of energy depositions in small-size targets by electrons and ions. Radiat. Prot. Dosim. 2011, 143, 145–151. [Google Scholar] [CrossRef]
- Liamsuwan, T.; Emfietzoglou, D.; Uehara, S.; Nikjoo, H. Microdosimetry of low-energy electrons. Int. J. Radiat. Biol. 2012, 88, 899–907. [Google Scholar] [CrossRef]
- Taleei, R.; Girard, P.M.; Sankaranarayanan, K.; Nikjoo, H. The non-homologous end-joining (NHEJ) mathematical model for the repair of double-strand breaks: II. Application to damage induced by ultrasoft X rays and low-energy electrons. Radiat. Res. 2013, 179, 540–548. [Google Scholar] [CrossRef]
- Watanabe, R.; Rahmanian, S.; Nikjoo, H. Spectrum of radiation-induced clustered non-DSB damage—A Monte Carlo track structure modeling and calculations. Radiat. Res. 2015, 183, 525–540. [Google Scholar] [CrossRef]
- Friedland, W.; Jacob, P.; Paretzke, H.G.; Stork, T. Monte Carlo simulation of the production of short DNA fragments by low-linear energy transfer radiation using higher-order DNA models. Radiat. Res. 1998, 150, 170–182. [Google Scholar] [CrossRef]
- Friedland, W.; Jacob, P.; Bernhardt, P.; Paretzke, H.G.; Dingfelder, M. Simulation of DNA damage after proton irradiation. Radiat. Res. 2003, 159, 401–410. [Google Scholar] [CrossRef]
- Friedland, W.; Jacob, P.; Kundrát, P. Stochastic simulation of DNA double-strand break repair by non-homologous end joining based on track structure calculations. Radiat. Res. 2010, 73, 677–688. [Google Scholar] [CrossRef]
- Friedland, W.; Dingfelder, M.; Kundrát, P.; Jacob, P. Track structures, DNA targets and radiation effects in the biophysical Monte Carlo simulation code PARTRAC. Mutat. Res. Mol. Mech. Mutagen. 2011, 711, 28–40. [Google Scholar] [CrossRef]
- Friedland, W.; Kundrát, P.; Jacob, P. Stochastic modelling of DSB repair after photon and ion irradiation. Int. J. Radiat. Biol. 2012, 88, 129–136. [Google Scholar] [CrossRef]
- Friedland, W.; Schmitt, E.; Kundrát, P.; Dingfelder, M.; Baiocco, G.; Barbieri, S.; Ottolenghi, A. Comprehensive track-structure based evaluation of DNA damage by light ions from radiotherapy-relevant energies down to stopping. Sci. Rep. 2017, 7, 45161. [Google Scholar] [CrossRef]
- Li, J.; Li, C.; Qiu, R.; Yan, C.; Xie, W.; Zeng, Z.; Tung, C. Comparison of direct DNA strand breaks induced by low energy electrons with different inelastic cross sections. Nucl. Instrum. Methods Phys. Res. Sect. B 2013, 311, 27–36. [Google Scholar] [CrossRef]
- Li, J.; Li, C.; Qiu, R.; Yan, C.; Xie, W.; Wu, Z.; Zeng, Z.; Tung, C. DNA strand breaks induced by electrons simulated with Nanodosimetry Monte Carlo Simulation Code: NASIC. Radiat. Prot. Dosim. 2015, 166, 38–43. [Google Scholar] [CrossRef]
- Chow, J.C.L. Dose enhancement effect in radiotherapy: Adding gold nanoparticles to tumor in cancer treatment. In Nanostructures for Cancer Therapy; Ficai, A., Grumezescu, A.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 383–403. [Google Scholar]
- Plante, I.; Cucinotta, F.A. Ionization and excitation cross sections for the interaction of HZE particles in liquid water and application to Monte Carlo simulation of radiation tracks. New J. Phys. 2008, 10, 125020. [Google Scholar] [CrossRef] [Green Version]
- Plante, I.; Cucinotta, F.A. Cross sections for the interactions of 1 eV–100 MeV electrons in liquid water and application to Monte-Carlo simulation of HZE radiation tracks. New J. Phys. 2009, 11, 063047. [Google Scholar] [CrossRef]
- Plante, I.; Devroye, L. Considerations for the independent reaction times and step-by-step methods for radiation chemistry simulations. Radiat. Phys. Chem. 2017, 139, 157–172. [Google Scholar] [CrossRef]
- Plante, I.; Ponomarev, A.; Patel, Z.; Slaba, T.; Hada, M. RITCARD: Radiation-Induced Tracks, Chromosome Aberrations, Repair and Damage. Radiat. Res. 2019, 192, 282–298. [Google Scholar] [CrossRef]
- Incerti, S.; Baldacchino, G.; Bernal, M.; Capra, R.; Champion, C.; Francis, Z. The geant4-dna project. Int. J. Model. Simul. Sci. Comput. 2010, 1, 157–178. [Google Scholar] [CrossRef]
- Incerti, S.; Ivanchenko, A.; Karamitros, M.; Mantero, A.; Moretto, P.; Tran, N.H.; Mascialino, B.; Champion, C.; Ivanchenko, V.; Bernal, A.M.; et al. Comparison of GEANT4 very low energy cross section models with experimental data in water. Med. Phys. 2010, 37, 4692–4708. [Google Scholar] [CrossRef]
- Bernal, A.M.; Bordage, M.; Brown, J.; Davídková, M.; Delage, E.; Bitar, E.Z.; Enger, S.; Francis, Z.; Guatelli, S.; Ivanchenko, V.; et al. Track structure modeling in liquid water: A review of the Geant4-DNA very low energy extension of the Geant4 Monte Carlo simulation toolkit. Phys. Med. 2015, 31, 861–874. [Google Scholar] [CrossRef]
- Agostinelli, S.; Allison, J.; Amako, K.; Apostolakis, J.; Araujo, H.; Arce, P.; Asai, M.; Axen, D.; Banerjee, S.; Barrand, G.; et al. GEANT4—A simulation toolkit. Nucl. Instrum. Methods Phys. Res. Sect. A 2003, 506, 250–303. [Google Scholar] [CrossRef] [Green Version]
- Apostolakis, J.; Asai, M.; Bogdanov, A.; Burkhardt, H.; Cosmo, G.; Elles, S.; Folger, G.; Grichine, V.; Gumplinger, P.; Heikkinen, A.; et al. Geometry and physics of the geant4 toolkit for high and medium energy applications. Radiat. Phys. Chem. 2009, 78, 859–873. [Google Scholar] [CrossRef]
- Nikjoo, H.; Goodhead, D.T.; Charlton, E.D.; Paretzke, H.G. Energy deposition in small cylindrical targets by ultrasoft X-rays. Phys. Med. Biol. 1989, 34, 691. [Google Scholar] [CrossRef]
- Charlton, D.E.; Nikjoo, H.; Humm, J.L. Calculation of initial yields of single-and double-strand breaks in cell nuclei from electrons, protons and alpha particles. Int. J. Radiat. Biol. 1989, 56, 1–19. [Google Scholar] [CrossRef]
- Pomplun, E. A new DNA target model for track structure calculations and its first application to I-125 Auger electrons. Int. J. Radiat. Biol. 1991, 59, 625–642. [Google Scholar] [CrossRef]
- Pinak, M.; Ito, A. Energy deposition in structural parts of DNA by monoenergetic electrons. J. Radiat. Res. 1993, 34, 221–234. [Google Scholar] [CrossRef]
- Nikjoo, H.; Martin, R.F.; Charlton, D.E.; Terrissol, M.; Kandaiya, S.; Lobachevsky, P. Modelling of Auger-induced DNA damage by incorporated125I. Acta Oncol. 1996, 5, 849–856. [Google Scholar] [CrossRef] [Green Version]
- Holley, W.R.; Chatterjee, A. Clusters of DNA damage induced by ionizing radiation: Formation of short DNA fragments. I. Theoretical modeling. Radiat. Res. 1996, 45, 188–199. [Google Scholar] [CrossRef]
- Michalik, V.; Begusová, M. Target model of nucleosome particle for track structure calculations and DNA damage modelling. Int. J. Radiat. Biol. 1994, 66, 267–277. [Google Scholar] [CrossRef]
- Kreth, G.; Finsterle, J.; von Hase, J.; Cremer, M.; Cremer, C. Radial arrangement of chromosome territories in human cell nuclei: A computer model approach based on gene density indicates a probabilistic global positioning code. Biophys. J. 2004, 86, 2803–2812. [Google Scholar] [CrossRef] [Green Version]
- Friedland, W.; Paretzke, H.G.; Ballarini, F.; Ottolenghi, A.; Kreth, G.; Cremer, C. First steps towards systems radiation biology studies concerned with DNA and chromosome structure within living cells. Radiat. Environ. Biophys. 2008, 47, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Nikjoo, H.; Girard, P. A model of the cell nucleus for DNA damage calculations. Int. J. Radiat. Biol. 2012, 88, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Bernal, M.; Sikansi, D.; Cavalcante, F.; Incerti, S.; Champion, C.; Ivanchenko, V.; Francis, Z. An atomistic geometrical model of the B-DNA configuration for DNA–radiation interaction simulations. Comput. Phys. Commun. 2013, 184, 2840–2847. [Google Scholar] [CrossRef]
- Santos, M.D. Modélisation de la Topologie des Dépôts D’énergie Créés par un Rayonnement Ionisant à l’Échelle Nanométrique dans les Noyaux Cellulaires et Relation avec les Événements Précoces Radio-Induits. Ph.D. Thesis, Université de Bordeaux, Bordeaux, France, 2013. [Google Scholar]
- Friedland, W.; Kundrát, P. Track structure based modelling of chromosome aberrations after photon and alpha-particle irradiation. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2013, 756, 213–223. [Google Scholar] [CrossRef]
- Delage, E.; Pham, Q.; Karamitros, M.; Payno, H.; Stepan, V.; Incerti, S.; Maigne, L.; Perrot, Y. PDB4DNA: Implementation of DNA geometry from the Protein Data Bank (PDB) description for Geant4-DNA Monte-Carlo simulations. Comput. Phys. Commun. 2015, 192, 282–288. [Google Scholar] [CrossRef] [Green Version]
- Meylan, S. Développement d’un Outil de Simulation Multi-Échelle Adapté au Calcul des Dommages Radio-Induits Précoces dans des Cellules Exposées à des Irradiations d’Ions Légers (Proton et Alpha). Ph.D. Thesis, Université de Bordeaux, Bordeaux, France, 2016. [Google Scholar]
- McNamara, A.L.; Ramos-Méndez, J.; Perl, J.; Held, K.; Dominguez, N.; Moreno, E.; Henthorn, N.T.; Kirkby, K.J.; Meylan, S.; Villagrasa, C.; et al. Geometrical structures for radiation biology research as implemented in the TOPAS-nBio toolkit. Phys. Med. Biol. 2018, 63, 175018. [Google Scholar] [CrossRef] [Green Version]
- Papanikolaou, N.; Mayles, P.; Nahum, A.; Rosenwald, J.C. Handbook of Radiotherapy Physics: Theory and Practice; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Incerti, S.; Kyriakou, I.; Bernal, M.A.; Bordage, M.C.; Francis, Z.; Guatelli, S.; Ivanchenko, V.; Karamitros, M.; Lampe, N.; Lee, S.B.; et al. Geant4-DNA example applications for track structure simulations in liquid water: A report from the Geant4-DNA Project. Med. Phys. 2018, 45, 722–739. [Google Scholar] [CrossRef] [Green Version]
- Kyriakou, I.; Incerti, S.; Francis, Z. Improvements in geant4 energy-loss model and the effect on low-energy electron transport in liquid water. Med. Phys. 2015, 42, 3870–3876. [Google Scholar] [CrossRef]
- Kyriakou, I.; Šefl, M.; Nourry, V.; Incerti, S. The impact of new Geant4-DNA cross section models on electron track structure simulations in liquid water. J. Appl. Phys. 2016, 119, 194902. [Google Scholar] [CrossRef]
- Shin, W.G. Development and Application of the Geant4-DNA Toolkit for the Simulation of Radiobiological Effects at the Sub-Cellular Scale. Ph.D. Thesis, Université de Bordeaux, Bordeaux, France, 2020. [Google Scholar]
- Shin, W.-G.; Bordage, M.-C.; Emfietzoglou, D.; Kyriakou, I.; Sakata, D.; Min, C.H.; Lee, S.B.; Guatelli, S.; Incerti, S. Development of a new Geant4-DNA electron elastic scattering model for liquid-phase water using the ELSEPA code. J. Appl. Phys. 2018, 124, 224901. [Google Scholar] [CrossRef] [Green Version]
- Lund, C.; Famulari, G.; Montgomery, L.; Kildea, J. A microdosimetric analysis of the interactions of mono-energetic neutrons with human tissue. Phys. Med. 2020, 73, 29–42. [Google Scholar] [CrossRef]
- Montgomery, L.; Lund, C.M.; Landry, A.; Kildea, J. Towards the characterization of neutron carcinogenesis through direct action simulations of clustered DNA damage. Phys. Med. Biol. 2021, 66, 205011. [Google Scholar] [CrossRef]
- McNamara, A.; Geng, C.; Turner, R.; Mendez, J.R.; Perl, J.; Held, K.; Faddegon, B.; Paganetti, H.; Schuemann, J. Validation of the radiobiology toolkit TOPAS-nBio in simple DNA geometries. Phys. Med. 2017, 33, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Schuemann, J.; McNamara, A.L.; Ramos-Méndez, J.; Held, K.D.; Paganetti, H.; Incerti, S.; Faddegon, B. TOPAS-nBio: An extension to the TOPAS simulation toolkit for cellular and sub-cellular radiobiology. Radiat. Res. 2019, 191, 125–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perl, J.; Shin, J.; Schümann, J.; Faddegon, B.; Paganetti, H. TOPAS: An innovative proton Monte Carlo platform for research and clinical applications. Med. Phys. 2012, 39, 6818–6837. [Google Scholar] [CrossRef]
- Ashley, J.C. Interaction of low-energy electrons with condensed matter: Stopping powers and inelastic mean free paths from optical data. J. Electron. Spectrosc. Relat. Phenom. 1988, 46, 199–214. [Google Scholar] [CrossRef]
- Emfietzoglou, D.; Papamichael, G.; Nikjoo, H. Monte Carlo electron track structure calculations in liquid water using a new model dielectric response function. Radiat. Res. 2017, 188, 355–368. [Google Scholar] [CrossRef]
- Wiklund, K.; Fernández-Varea, J.M.; Lind, B.K. A Monte Carlo program for the analysis of low-energy electron tracks in liquid water. Phys. Med. Biol. 2011, 56, 1985. [Google Scholar] [CrossRef] [PubMed]
- Wilson, W.E.; Miller, J.H.; Lynch, D.J.; Lewis, R.R.; Batdorf, M. Analysis of low-energy electron track structure in liquid water. Radiat. Res. 2004, 161, 591–596. [Google Scholar] [CrossRef]
- Watt, D.E. Quantities for Generalized Dosimetry of Ionizing Radiations in Liquid Water; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Seltzer, S.M.; Fernandez-Varea, J.M.; Andreo, P.; Bergstrom, P.M.; Burns, D.T.; Krajcar, B. Key Data for Ionizing-Radiation Dosimetry: Measurement Standards and Applications, ICRU Report 90. 2016. Available online: https://www.scribd.com/document/478503312/ICRU-90-KEY-DATA-FOR-IONIZING-RADIATIONDOSIMETRY-MEASUREMENT-STANDARDS-AND-APPLICATIONS (accessed on 24 April 2022).
- Meesungnoen, J.; Jay-Gerin, J.-P.; Filali-Mouhim, A.; Mankhetkorn, S. Low-energy electron penetration range in liquid water. Radiat. Res. 2002, 158, 657–660. [Google Scholar] [CrossRef]
- Nikjoo, H.; Uehara, S.; Emfietzoglou, D.; Cucinotta, F.; Nikjoo, H.; Uehara, S.; Emfietzoglou, D.; Cucinotta, F. Track-structure codes in radiation research. Radiat. Meas. 2006, 41, 1052–1074. [Google Scholar] [CrossRef]
- Sakata, D.; Belov, O.; Bordage, M.; Emfietzoglou, D.; Guatelli, S.; Inaniwa, T.; Ivanchenko, V.; Karamitros, M.; Kyriakou, I.; Lampe, N.; et al. Fully integrated Monte Carlo simulation for evaluating radiation induced DNA damage and subsequent repair using Geant4-DNA. Sci. Rep. 2020, 10, 20788. [Google Scholar] [CrossRef]
- Frankenberg, D.; Brede, H.J.; Schrewe, U.J.; Steinmetz, C.; Frankenberg-Schwager, M.; Kasten, G.; Pralle, E. Induction of DNA double-strand breaks by 1 H and 4 He ions in primary human skin fibroblasts in the LET range of 8 to 124 keV/μm. Radiat. Res. 1999, 151, 540–549. [Google Scholar] [CrossRef]
- Campa, A.; Ballarini, F.; Belli, M.; Cherubini, R.; Dini, V.; Esposito, G.; Friedland, W.; Gerardi, S.; Molinelli, S.; Ottolenghi, A.; et al. DNA DSB induced in human cells by charged particles and gamma rays: Experimental results and theoretical approaches. Int. J. Radiat. Biol. 2005, 81, 841–854. [Google Scholar] [CrossRef] [PubMed]
- Lampe, N.; Karamitros, M.; Breton, V.; Brown, J.M.; Kyriakou, I.; Sakata, D.; Sarramia, D.; Incerti, S. Mechanistic DNA damage simulations in Geant4-DNA part 1: A parameter study in a simplified geometry. Phys. Med. 2018, 48, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Henthorn, N.T.; Warmenhoven, J.W.; Sotiropoulos, M.; Mackay, R.I.; Kirkby, K.J.; Merchant, M.J. Nanodosimetric simulation of direct ion-induced DNA damage using different chromatin geometry models. Radiat. Res. 2017, 188, 770–783. [Google Scholar] [CrossRef] [PubMed]
- Finch, J.T.; Klug, A. Solenoidal model for superstructure in chromatin. Proc. Natl. Acad. Sci. USA 1976, 73, 1897–1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordage, M.C.; Bordes, J.; Edel, S.; Terrissol, M.; Franceries, X.; Bardiès, M.; Lampeef, N.; Incertief, S. Implementation of new physics models for low energy electrons in liquid water in Geant4-DNA. Phys. Med. 2016, 32, 1833–1840. [Google Scholar] [CrossRef]
- Kellerer, A.M.; Chmelevsky, D. Concepts of microdosimetry. Radiat. Environ. Biophys. 1975, 12, 321–335. [Google Scholar] [CrossRef] [Green Version]
- Kyriakou, I.; Emfietzoglou, D.; Ivanchenko, V.; Bordage, M.C.; Guatelli, S.; Lazarakis, P.; Tran, N.H.; Incerti, S. Microdosimetry of electrons in liquid water using the low-energy models of Geant4. J. Appl. Phys. 2017, 122, 024303. [Google Scholar] [CrossRef]
- Humm, J.; Charlton, D. A new calculational method to assess the therapeutic potential of Auger electron emission. Int. J. Radiat. Oncol. 1989, 17, 351–360. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process | Physics Constructor | ||||
|---|---|---|---|---|---|
| Option8 | Option2and4 | Option2 with ELSEPA | Option4 with ElSEPA | Option6 with ElSEPA | |
| Elastic | CPA100 Model (11 eV–256 keV) | Uehara Model (10 eV–10 keV) | ELSEPA Model (10 eV–1 GeV) | ELSEPA Model (10 eV–1 GeV) | ELSEPA Model (10 eV–1 GeV) |
| Champion Model (256 keV–1 MeV) | Champion Model (10 keV–1 MeV) | ||||
| Ionization | Born Model (11 eV–1 MeV) | Emfietzoglou Model (10 eV–10 keV) | Born Model (11 eV–1 MeV) | Emfietzoglou Model (10 eV–10 keV) | CPA100 Model (11 eV–256 keV) |
| Born Model (10 keV–1 MeV) | |||||
| Excitation | Born Model (9 eV–1 MeV) | Emfietzoglou model (8 eV–10 keV) | Born Model (9 eV–1 MeV) | Emfietzoglou Model (8 eV–10 keV) | CPA100 Model (11 eV–256 keV) |
| Born Model (10 keV–1 MeV) | |||||
| Process | Physics Constructor | ||
|---|---|---|---|
| Option2 | Option4 | Option6 | |
| Elastic | Champion model | Uehara model | CPA100 model (64) |
| (7.4 eV–1 MeV) | (9 eV–10 keV) | (11 eV–256 keV) | |
| Ionization | Born Model | Emfietzoglou model (54) | CPA100 model |
| (11 eV–1 MeV) | (10 eV–10 keV) | (11 eV–256 keV) | |
| Excitation | Born Model | Emfietzoglou model | CPA100 model |
| (9 eV–1 MeV) | (8 eV–10 keV) | (11 eV–256 keV) | |
| Attachment | Melton model | - | - |
| (4–13 eV) | |||
| VibExcitationa | Sanche model | - | - |
| (2–100 eV) | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, K.; Wu, C.; Peng, X.; Ji, X.; Luo, S.; Liu, Y.; Wang, X. Nanoscale Calculation of Proton-Induced DNA Damage Using a Chromatin Geometry Model with Geant4-DNA. Int. J. Mol. Sci. 2022, 23, 6343. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116343
Zhu K, Wu C, Peng X, Ji X, Luo S, Liu Y, Wang X. Nanoscale Calculation of Proton-Induced DNA Damage Using a Chromatin Geometry Model with Geant4-DNA. International Journal of Molecular Sciences. 2022; 23(11):6343. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116343
Chicago/Turabian StyleZhu, Kun, Chun Wu, Xiaoyu Peng, Xuantao Ji, Siyuan Luo, Yuchen Liu, and Xiaodong Wang. 2022. "Nanoscale Calculation of Proton-Induced DNA Damage Using a Chromatin Geometry Model with Geant4-DNA" International Journal of Molecular Sciences 23, no. 11: 6343. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23116343