Enhanced Inhibitory Effect of Ultra-Fine Granules of Red Ginseng on LPS-induced Cytokine Expression in the Monocyte-Derived Macrophage THP-1 Cells

Abstract
:1. Introduction
2. Results
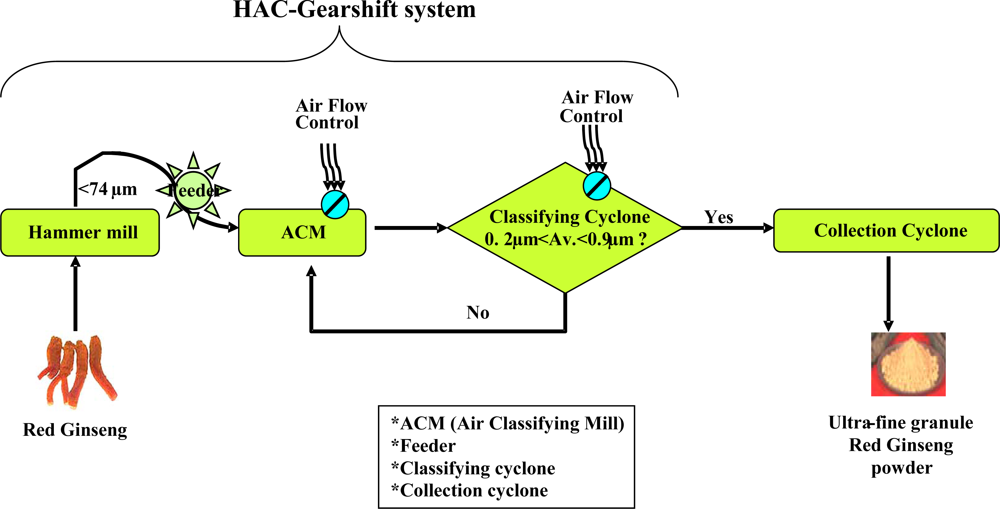
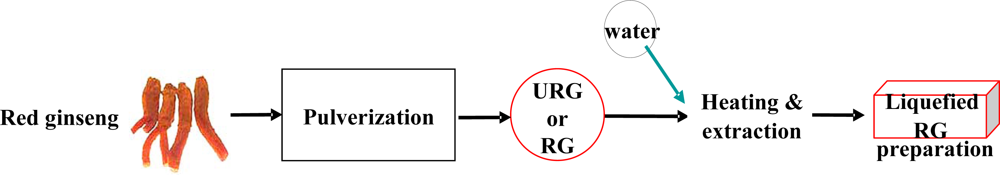
2.1. Biophysical properties of URG prepared by the HAC-gearshift system
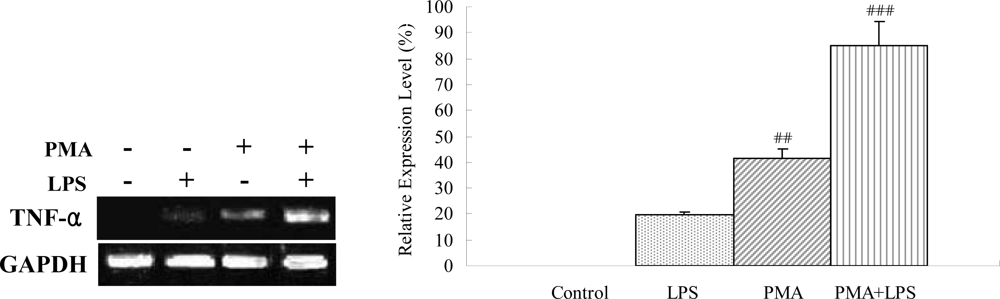
2.2. Effects of LPS and phobol 12-myristate 13-acetate in TNF-α expression in the monocyte-driven macrophage THP-1 cells
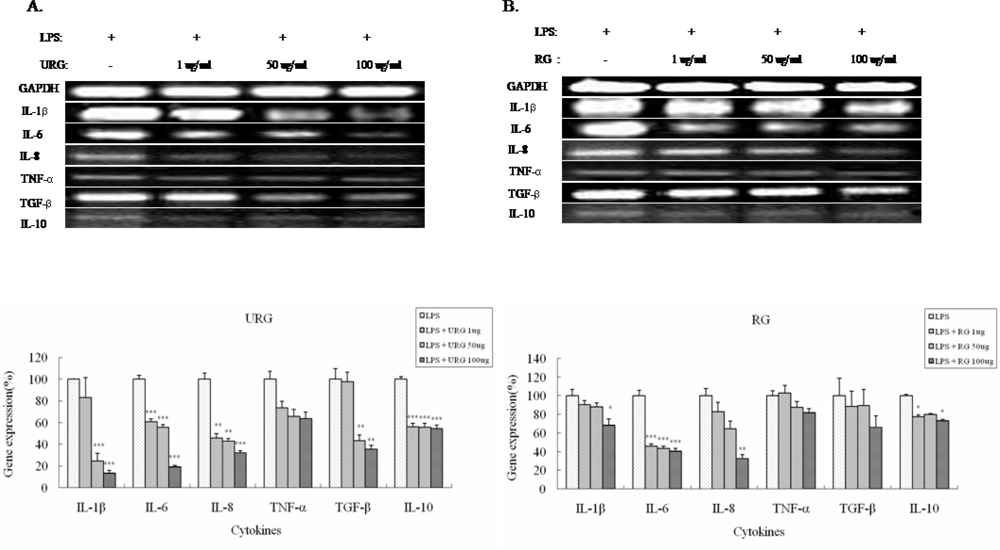
2.3. Effect of URG and RG on LPS-induced cytokine expression in the THP-1 cells
3. Discussion
4. Experimental Section
4.1. Preparation of URG by the HAC-gearshift system
4.2. Reagents
4.3. Transmission electron microscopy (TEM, JEOL, Japan)
4.4. HPLC analysis
4.5. Cell culture conditions
4.6. PMA treatment
4.7. Cell viability
4.8. Macrophage differentiation and stimulation
4.9. Reverse transcription (RT)-PCR analysis
4.10. Statistical analysis
Acknowledgments
References
- Kiefer, D; Pantuso, T. Panax Ginseng. Am. Fam. Physician 2003, 68, 1539–1542. [Google Scholar]
- Min, JK; Kim, JH; Cho, YL; Maeng, YS; Lee, SJ; Pyun, BJ; Kim, YM; Park, JH; Kwon, YG. 20(S)-Ginsenoside Rg3 Prevents Endothelial Cell Apoptosis via Inhibition of a Mitochondrial Caspase Pathway. Biochem. Biophys. Res. Commun 2006, 349, 987–994. [Google Scholar]
- Averill, L; Toossi, Z; Aung, H; Boom, WH; Ellner, JJ. Regulation of Production of Tumor Necrosis Factor Alpha in Monocytes Stimulated by the 30-Kilodalton Antigen of Mycobacterium Tuberculosis. Infect. Immun 1995, 63, 3206–3208. [Google Scholar]
- Kostyal, DA; Butler, GH; Beezhold, DH. Mycoplasma Hyorhinis Molecules That Induce Tumor Necrosis Factor Alpha Secretion by Human Monocytes. Infect. Immun 1995, 63, 3858–3863. [Google Scholar]
- Terlikowski, SJ. Tumour Necrosis Factor and Cancer Treatment: A Historical Review and Perspectives. Rocz. Akad. Med. Bialymst 2001, 46, 5–18. [Google Scholar]
- Beuscher, HU; Rodel, F; Forsberg, A; Rollinghoff, M. Bacterial Evasion of Host Immune Defense: Yersinia Enterocolitica Encodes a Suppressor for Tumor Necrosis Factor Alpha Expression. Infect. Immun 1995, 63, 1270–1277. [Google Scholar]
- Caron, E; Gross, A; Liautard, JP; Dornand, J. Brucella Species Release a Specific, Protease-Sensitive, Inhibitor of TNF-Alpha Expression, Active on Human Macrophage-Like Cells. J. Immunol 1996, 156, 2885–2893. [Google Scholar]
- Oppenheim, JJ. Cytokines: Past, Present, and Future. Int. J. Hematol 2001, 74, 3–8. [Google Scholar]
- Spelman, K; Burns, J; Nichols, D; Winters, N; Ottersberg, S; Tenborg, M. Modulation of Cytokine Expression by Traditional Medicines: A Review of Herbal Immunomodulators. Altern. Med. Rev 2006, 11, 128–150. [Google Scholar]
- Liu, Y; Li, W; Li, P; Deng, MC; Yang, SL; Yang, L. The Inhibitory Effect of Intestinal Bacterial Metabolite of Ginsenosides on CYP3A Activity. Biol. Pharm. Bull 2004, 27, 1555–1560. [Google Scholar]
- Auwerx, J. The Human Leukemia Cell Line, THP-1: A Multifacetted Model for the Study of Monocyte-Macrophage Differentiation. Experientia 1991, 47, 22–31. [Google Scholar]
- Auwerx, J; Staels, B; Van Vaeck, F; Ceuppens, JL. Changes in IgG Fc Receptor Expression Induced by Phorbol 12-Myristate 13-Acetate Treatment of THP-1 Monocytic Leukemia Cells. Leuk. Res 1992, 16, 317–327. [Google Scholar]
- Glauser, MP; Zanetti, G; Baumgartner, JD; Cohen, J. Septic Shock: Pathogenesis. Lancet 1991, 338, 732–736. [Google Scholar]
- Ulevitch, RJ; Tobias, PS. Receptor-Dependent Mechanisms of Cell Stimulation by Bacterial Endotoxin. Annu. Rev. Immunol 1995, 13, 437–457. [Google Scholar]
- Raetz, CRH; Ulevitch, RJ; Wright, SD; Sibley, CH; Ding, A; Nathan, CF. Gram-negative Endotoxin: An Extraordinary Lipid with Profound Effects on Eukaryotic Signal Transduction. FASEB J 1991, 5, 2652–2660. [Google Scholar]
- Natanson, C; Hoffman, WD; Suffredini, AF; Eichacker, PQ; Danner, RL. Selected Treatment Strategies for Septic Shock Based on Proposed Mechanisms of Pathogenesis. Annals Internal Med 1994, 120, 771. [Google Scholar]
- Eliopoulos, AG; Dumitru, CD; Wang, CC; Cho, J; Tsichlis, PN. Induction of COX-2 by LPS in Macrophages is Regulated by Tpl2-dependent CREB Activation Signals. EMBO J 2002, 21, 831. [Google Scholar]
- Pang, L; Sawada, T; Decker, SJ; Saltiel, AR. Inhibition of MAP Kinase Kinase Blocks the Differentiation of PC-12 Cells Induced by Nerve Growth Factor. J. Biol. Chem 1995, 270, 13585–13588. [Google Scholar]
- Ma, W; Lim, W; Gee, K; Aucoin, S; Nandan, D; Kozlowski, M; Diaz-Mitoma, F; Kumar, A. The p38 Mitogen-activated Kinase Pathway Regulates the Human Interleukin-10 Promoter via the Activation of Sp1 Transcription Factor in Lipopolysaccharide-stimulated Human Macrophages. J. Biol. Chem 2001, 276, 13664–13674. [Google Scholar]
- Kumar, A; Angel, JB; Daftarian, MP; Parato, K; Cameron, WD; Filion, L; Diaz-Mitoma, F. Differential Production of IL-10 by T Cells and Monocytes of HIV-infected Individuals: Association of IL-10 Production with CD28-mediated Immune Responsiveness. Clin. Exp. Immunol 1998, 114, 78–86. [Google Scholar]
- Wu, JY. Saponin Adjuvant Enhancement of Antigen-specific Immune Responses to an Experimental HIV-1 Vaccine. J. Immunol 1992, 148, 1519–1525. [Google Scholar]
- Bae, EA; Han, MJ; Choo, MK; Park, SY; Kim, DH. Metabolism of 20 (S)-and 20 (R)-Ginsenoside Rg3 by Human Intestinal Bacteria and Its Relation to in vitro Biological Activities. Biol. Pharmaceut. Bull 2002, 25, 58–63. [Google Scholar]
- Kim, ND; Kang, SY; Kim, MJ; Park, JH; Schini-Kerth, VB. The Ginsenoside Rg3 Evokes Endothelium-independent Relaxation in Rat Aortic Rings: Role of K+ Channels. Europ. J. Pharmacol 1999, 367, 51–57. [Google Scholar]
- Wakabayashi, C; Hasegawa, H; Murata, J; Saiki, I. In vivo Antimetastatic Action of Ginseng Protopanaxadiol Saponins Is Based on Their Intestinal Bacterial Metabolites After Oral Administration. Oncol. Res 1997, 9, 411–417. [Google Scholar]
- Park, WS; Shin, DY; Kim, DR; Yang, WM; Chang, MS; Park, SK. Korean Ginseng Induces Spermatogenesis in Rats Through the Activation of Camp-Responsive Element Modulator (CREM). Fertil. Steril 2007, 88, 1000–1002. [Google Scholar]
- Alessi, DR; Cuenda, A; Cohen, P; Dudley, DT; Saltiel, AR. PD 098059 Is a Specific Inhibitor of the Activation of Mitogen-activated Protein Kinase Kinase in vitro and in vivo. J. Biol. Chem 1995, 270, 27489–27494. [Google Scholar]
- O’Toole, SA; Sheppard, BL; McGuinness, EPJ; Gleeson, NC; Yoneda, M; Bonnar, J. The MTS Assay as an Indicator of Chemosensitivity/Resistance in Malignant Gynaecological Tumours. Canc. Detect. Prevent 2003, 27, 47–54. [Google Scholar]
- Dixon, CJ; Bowler, WB; Littlewood-Evans, A; Dillon, JP; Bilbe, G; Sharpe, GR; Gallagher, JA. Regulation of Epidermal Homeostasis Through P2Y 2 Receptors. Brit. J. Pharmacol 1999, 127, 1680–1686. [Google Scholar]
- Glue, C; Hansen, JB; Schjerling, P; Jinquan, T; Poulsen, LK. LPS-induced Cytokine Production in the Monocytic Cell Line THP-1 Determined by Multiple Quantitative Competitive PCR (QC-PCR). Scand. J. Clin. Labor. Investig 2002, 62, 405–412. [Google Scholar]
- Na, YJ; Jeon, YJ; Suh, JH; Kang, JS; Yang, KH; Kim, HM. Suppression of IL-8 Gene Expression by Radicicol Is Mediated Through the Inhibition of ERK1/2 And p38 Signaling and Negative Regulation of NF- κB and AP-1. Int. Immunopharmacol 2001, 1, 1877–1887. [Google Scholar]
- de Groot-Kruseman, HA; Mol, WM; Niesters, HGM; Maat, APW; van Gelder, T; Balk, A; Weimar, W; Baan, CC. Differential Intragraft Cytokine Messenger RNA Profiles During Rejection and Repair of Clinical Heart Transplants. A Longitudinal Study. Transplant Int 2003, 16, 9–14. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GinsenosidesA | RG (mg/mL) | URG (mg/mL) | Fold increase (URG vs. RG) | |
|---|---|---|---|---|
| PD | Rb1 | 2.32 | 5.00 | 2.16 |
| Rb2 | 1.05 | 2.41 | 2.29 | |
| Rc | 1.10 | 2.50 | 2.27 | |
| Rd | 0.63 | 1.70 | 2.69 | |
| Rg3s | 0.84 | 1.70 | 2.02 | |
| Rg3r | 0.39 | 0.73 | 1.87 | |
| Rh2s | 0.14 | 0.28 | 2.00 | |
| PT | Re | 1.01 | 2.17 | 2.14 |
| Rf | 0.58 | 0.87 | 1.50 | |
| Rg1 | 0.60 | 0.95 | 1.57 | |
| Rg2s | 0.83 | 1.49 | 1.79 | |
| Rg2r | 0.87 | 0.59 | 0.68 | |
| Rh1 | 0.33 | 0.48 | 1.45 | |
| Total sum | 10.69 | 20.87 | 1.95 | |
| CytokinesA | Control | LPS | LPS+URG | LPS+RG | LPS+PD98059B |
|---|---|---|---|---|---|
| IL-1β | 0.0 | 100 ± 0.2 | 13.7 ± 2.4 | 59.2 ± 6.7 | 4.9 ± 1.3 |
| IL-6 | 0.0 | 100 ± 3.6 | 19.7 ± 0.8 | 60.5 ± 2.8 | 13.2 ± 0.5 |
| IL-8 | 0.3 | 100 ± 5.5 | 32.3 ± 1.8 | 48.6 ± 4.1 | 30.5 ± 1.7 |
| TNF-α | 6.5 ± 0.5 | 100 ± 7.4 | 66.0 ± 5.9 | 94.0 ± 4.2 | 11.8 ± 1.0 |
| TGF-β | 0.7 ± 0.3 | 100 ± 9.6 | 35.6 ± 3.5 | 86.5 ± 12.5 | 33.4 ± 3.1 |
| Name | Oligonucleotide sequence (5′–3′) | Expected PCR product (bp) | |
|---|---|---|---|
| GAPDH | sense | GGTGAAGGTCGGAGTCAACGG | 500[28] |
| anti-sense | GGTCATGAGTCCTTCCACGAT | ||
| IL-1β | sense | GGGCCTCAAGGAAAAGAATC | 470 [29] |
| anti-sense | AGCTGACTGTCCTGGCTGAT | ||
| IL-6 | sense | AAAGAGGCACTGGCAGAAAA | 408 [29] |
| anti-sense | GAGGTGCCCATGCTACATTT | ||
| IL-8 | sense | AGGGTTGCCAGATGCAATAC | 378 [30] |
| anti-sense | AGACTAGGGTTGCCAGA | ||
| TNF-α | sense | AGCCCATGTTGTAGCAAACC | 424 [31] |
| anti-sense | CCAAAGTAGACCTGCCCAGA | ||
| TGF-β | sense | GACTGCGGATCTCTGTGTCA | 480 |
| anti-sense | CTGGTCTCAAATGCCTGGAT |
© 2008 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/). This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Lee, H.-C.; Vinodhkumar, R.; Yoon, J.W.; Park, S.-K.; Lee, C.-W.; Kim, H.-Y. Enhanced Inhibitory Effect of Ultra-Fine Granules of Red Ginseng on LPS-induced Cytokine Expression in the Monocyte-Derived Macrophage THP-1 Cells. Int. J. Mol. Sci. 2008, 9, 1379-1392. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms9081379
Lee H-C, Vinodhkumar R, Yoon JW, Park S-K, Lee C-W, Kim H-Y. Enhanced Inhibitory Effect of Ultra-Fine Granules of Red Ginseng on LPS-induced Cytokine Expression in the Monocyte-Derived Macrophage THP-1 Cells. International Journal of Molecular Sciences. 2008; 9(8):1379-1392. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms9081379
Chicago/Turabian StyleLee, Hyoung-Cheol, Radhakrishnan Vinodhkumar, Jang W. Yoon, Seong-Kyu Park, Chang-Won Lee, and Hong-Yeoul Kim. 2008. "Enhanced Inhibitory Effect of Ultra-Fine Granules of Red Ginseng on LPS-induced Cytokine Expression in the Monocyte-Derived Macrophage THP-1 Cells" International Journal of Molecular Sciences 9, no. 8: 1379-1392. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms9081379