
Anti-Cancer Immune Reaction and Lymph Node Macrophage; A Review from Human and Animal Studies

Abstract
:1. The Critical Role of Lymph Nodes in Anti-Cancer Immunotherapy
2. Function of Lymph Node Macrophages in Mice
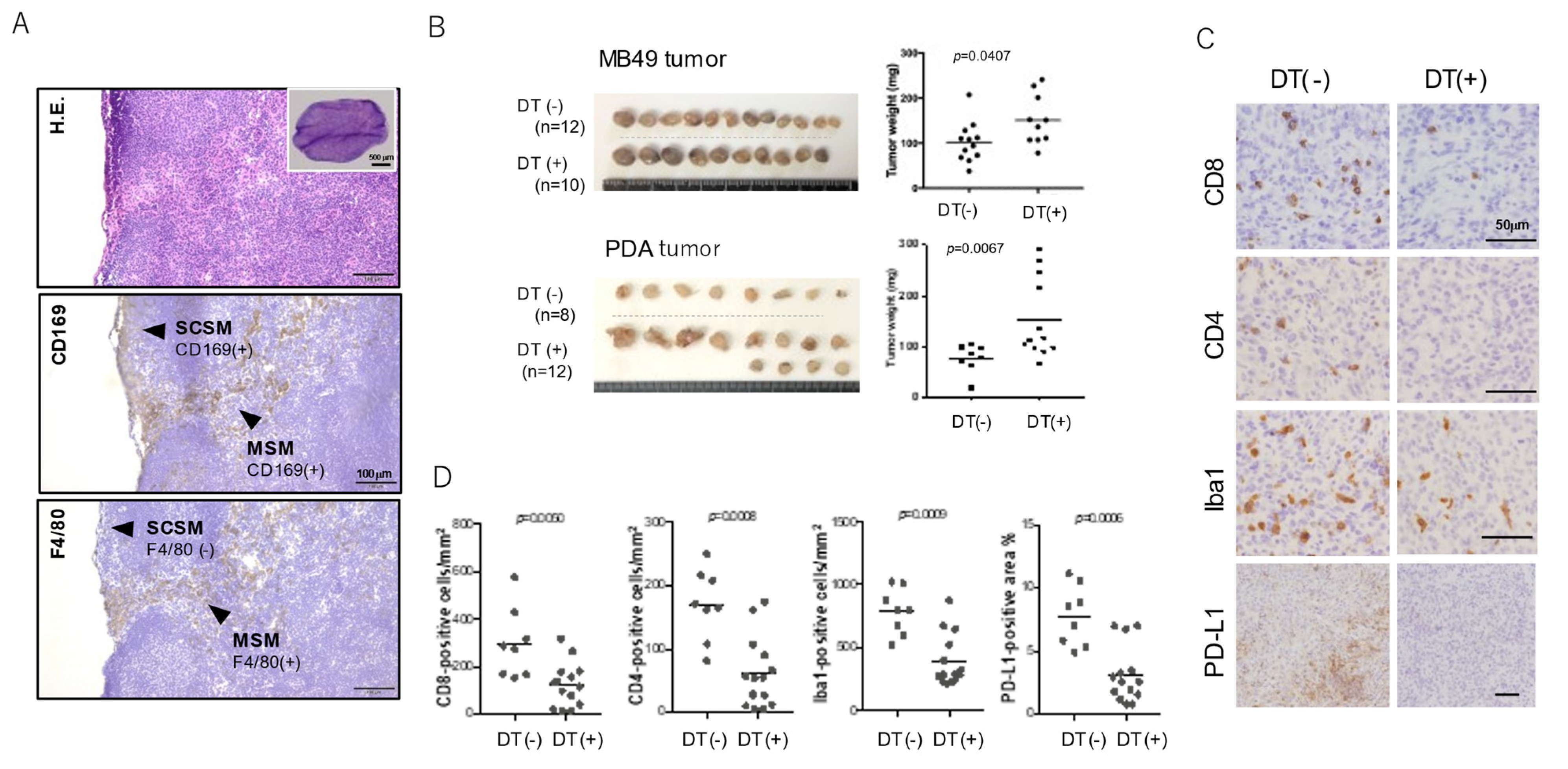
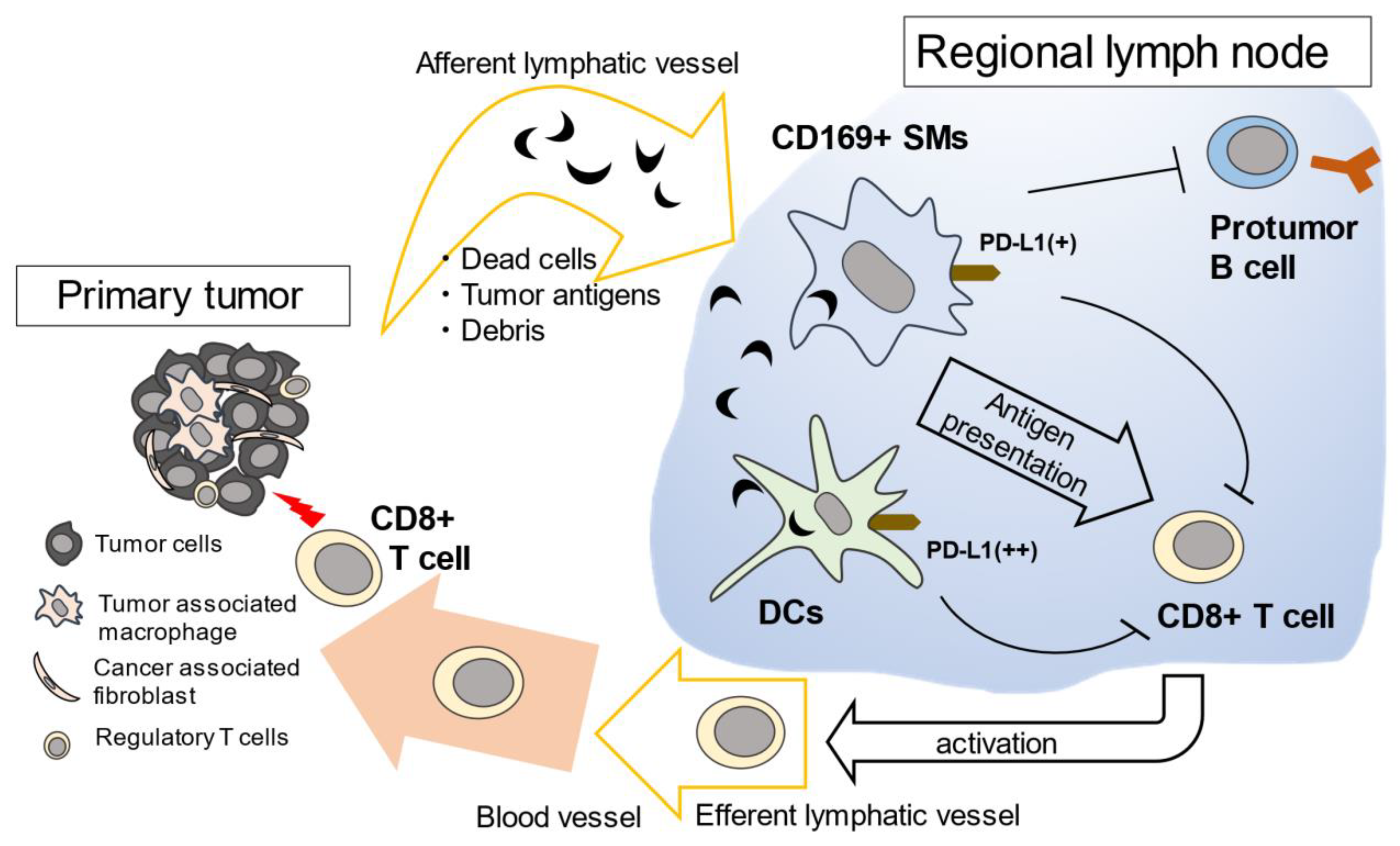
3. The Function of Lymph Node Macrophages in the Anti-Tumor Immune Response in Mice
4. The Function of SMs in Protumor Function of B-Lymphocytes in Mice
5. Potential Origin of SMs in Mice
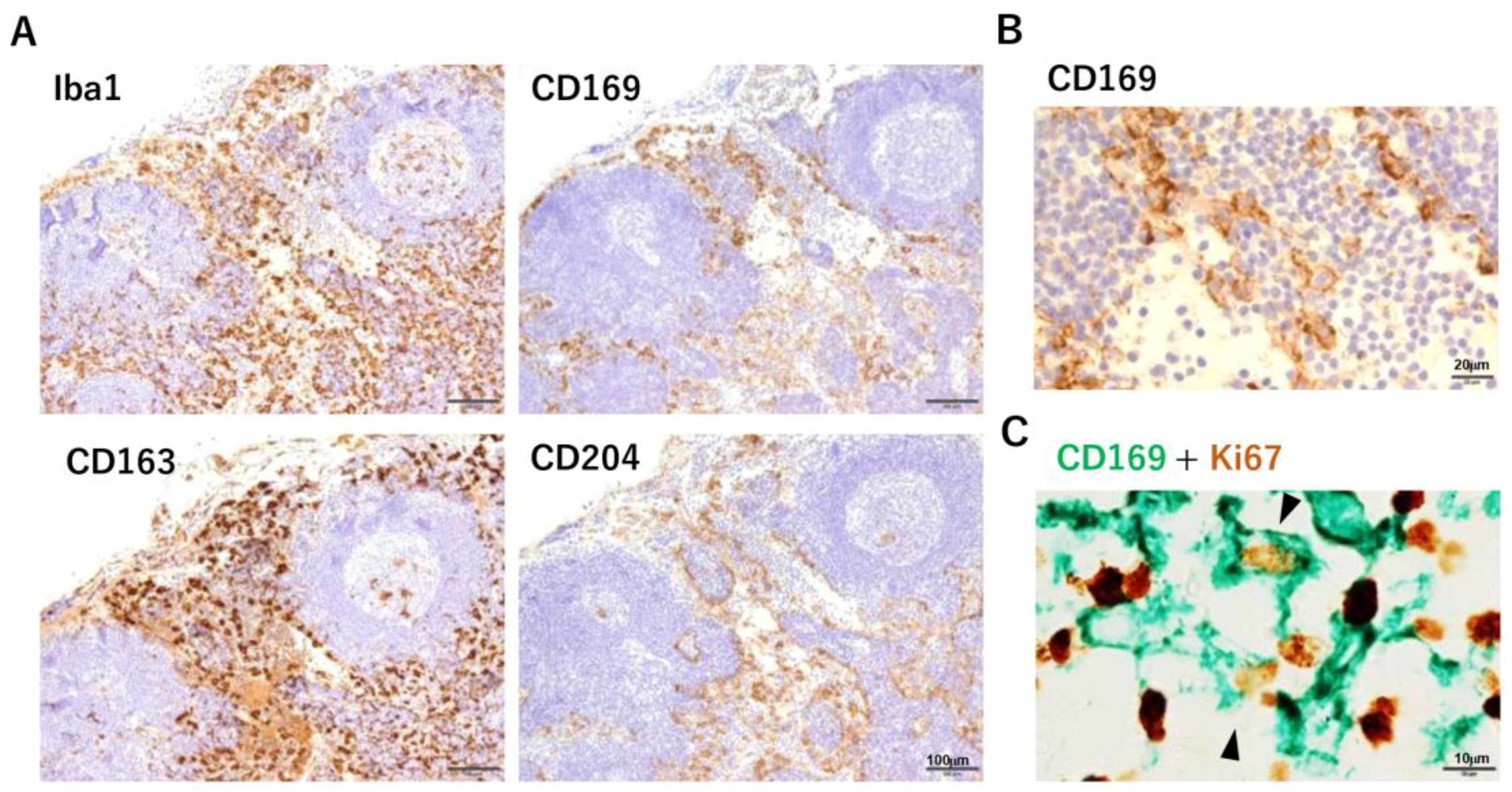
6. The Importance of SM in Human Solid Tumors
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Boon, T.; Cerottini, J.C.; Van den Eynde, B.; van der Bruggen, P.; Van Pel, A. Tumor antigens recognized by T lymphocytes. Annu. Rev. Immunol. 1994, 12, 337–365. [Google Scholar] [CrossRef]
- Chen, D.S.; Mellman, I. Oncology Meets Immunology: The Cancer-Immunity Cycle. Immunity 2013, 39, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zou, W.; Wolchok, J.D.; Chen, L. PD-L1 (B7-H1) and PD-1 pathway blockade for cancer therapy: Mechanisms, response biomarkers, and combinations. Sci. Transl. Med. 2016, 8, 328rv4. [Google Scholar] [CrossRef] [Green Version]
- Cable, J.; Greenbaum, B.; Pe’er, D.; Bollard, C.M.; Bruni, S.; Griffin, M.E.; Allison, J.P.; Wu, C.J.; Subudhi, S.K.; Mardis, E.R.; et al. Frontiers in cancer immunotherapy-a symposium report. Ann. N. Y. Acad. Sci. 2021, 1489, 30–47. [Google Scholar] [CrossRef] [PubMed]
- Andrews, L.P.; Yano, H.; Vignali, D.A.A. Inhibitory receptors and ligands beyond PD-1, PD-L1 and CTLA-4: Breakthroughs or backups. Nat. Immunol. 2019, 20, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Komohara, Y.; Harada, M.; Ohnishi, K.; Kumamoto, K.; Nakayama, T. PD-L1 expression in regional lymph nodes and predictable roles in anti-cancer immune responses. J. Clin. Exp. Hematop. 2020, 60, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Fransen, M.F.; Schoonderwoerd, M.; Knopf, P.; Camps, M.G.; Hawinkels, L.J.; Kneilling, M.; van Hall, T.; Ossendorp, F. Tumor-draining lymph nodes are pivotal in PD-1/PD-L1 checkpoint therapy. JCI Insight 2018, 3, e124507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Kassaye, B.; Wangmo, D.; Lou, E.; Subramanian, S. Chemotherapy but Not the Tumor Draining Lymph Nodes Determine the Immunotherapy Response in Secondary Tumors. iScience 2020, 23, 101056. [Google Scholar] [CrossRef]
- Dammeijer, F.; Van Gulijk, M.; Mulder, E.E.; Lukkes, M.; Klaase, L.; Bosch, T.V.D.; Van Nimwegen, M.; Lau, S.P.; Latupeirissa, K.; Schetters, S.; et al. The PD-1/PD-L1-Checkpoint Restrains T cell Immunity in Tumor-Draining Lymph Nodes. Cancer Cell 2020, 38, 685–700.e8. [Google Scholar] [CrossRef]
- Gray, E.E.; Cyster, J.G. Lymph node macrophages. J. Innate Immun. 2012, 4, 424–436. [Google Scholar] [CrossRef]
- Kuka, M.; Iannacone, M. The role of lymph node sinus macrophages in host defense. Ann. N. Y. Acad. Sci. 2014, 1319, 38–46. [Google Scholar] [CrossRef]
- Martinez-Pomares, L.; Gordon, S. CD169+ macrophages at the crossroads of antigen presentation. Trends Immunol. 2012, 33, 66–70. [Google Scholar] [CrossRef]
- Chen, W.C.; Kawasaki, N.; Nycholat, C.M.; Han, S.; Pilotte, J.; Crocker, P.R.; Paulson, J.C. Antigen delivery to macrophages using liposomal nanoparticles targeting si-aloadhesin/CD169. PLoS ONE 2012, 7, e39039. [Google Scholar]
- van Dinther, D.; Lopez Venegas, M.; Veninga, H.; Olesek, K.; Hoogterp, L.; Revet, M.; Ambrosini, M.; Kalay, H.; Stöckl, J.; van Kooyk, Y.; et al. Activation of CD8⁺ T Cell Responses after Melanoma Antigen Targeting to CD169⁺ Antigen Presenting Cells in Mice and Humans. Cancers 2019, 11, 183. [Google Scholar] [CrossRef] [Green Version]
- Poderoso, T.; Martínez, P.; Álvarez, B.; Handler, A.; Moreno, S.; Alonso, F.; Ezquerra, A.; Dominguez, J.; Revilla, C. Delivery of antigen to sialoadhesin or CD163 improves the specific immune response in pigs. Vaccine 2011, 29, 4813–4820. [Google Scholar] [CrossRef]
- Mellman, I.; Coukos, G.; Dranoff, G. Cancer immunotherapy comes of age. Nat. Cell Biol. 2011, 480, 480–489. [Google Scholar] [CrossRef]
- Das Mohapatra, A.; Tirrell, I.; Bénéchet, A.P.; Pattnayak, S.; Khanna, K.M.; Srivastava, P.K. Cross-dressing of CD8α+ Dendritic Cells with Antigens from Live Mouse Tumor Cells Is a Major Mechanism of Cross-priming. Cancer Immunol. Res. 2020, 8, 1287–1299. [Google Scholar] [CrossRef] [PubMed]
- Carey, W.; Prunieras, M. An improved method for the establishment of cell lines from primary skin melanomas. Br. J. Dermatol. 1984, 110, 265–271. [Google Scholar] [CrossRef]
- Asano, K.; Nabeyama, A.; Miyake, Y.; Qiu, C.-H.; Kurita, A.; Tomura, M.; Kanagawa, O.; Fujii, S.-I.; Tanaka, M. CD169-Positive Macrophages Dominate Antitumor Immunity by Crosspresenting Dead Cell-Associated Antigens. Immunity 2011, 34, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, T.; Ohnishi, K.; Kosaki, Y.; Saito, Y.; Horlad, H.; Fujiwara, Y.; Takeya, M.; Komohara, Y. Optimum immunohistochemical procedures for analysis of macrophages in human and mouse formalin fixed paraffin-embedded tissue samples. J. Clin. Exp. Hematop. 2017, 57, 31–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veninga, H.; Borg, E.G.F.; Vreeman, K.; Taylor, P.; Kalay, H.; Van Kooyk, Y.; Kraal, G.; Martinez-Pomares, L.; Haan, J.D. Antigen targeting reveals splenic CD169+ macrophages as promoters of germinal center B-cell responses. Eur. J. Immunol. 2015, 45, 747–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabowska, J.; Lopez-Venegas, M.; Affandi, A.J.; Haan, J.D. CD169+ Macrophages Capture and Dendritic Cells Instruct: The Interplay of the Gatekeeper and the General of the Immune System. Front. Immunol. 2018, 9, 2472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernhard, C.A.; Ried, C.; Kochanek, S.; Brocker, T. CD169+ macrophages are sufficient for priming of CTLs with specificities left out by cross-priming dendritic cells. Proc. Natl. Acad. Sci. USA 2015, 112, 5461–5466. [Google Scholar] [CrossRef] [Green Version]
- Muraoka, D.; Harada, N.; Hayashi, T.; Tahara, Y.; Momose, F.; Sawada, S.-I.; Mukai, S.-A.; Akiyoshi, K.; Shiku, H. Nanogel-Based Immunologically Stealth Vaccine Targets Macrophages in the Medulla of Lymph Node and Induces Potent Antitumor Immunity. ACS Nano 2014, 8, 9209–9218. [Google Scholar] [CrossRef] [PubMed]
- Saunderson, S.; Dunn, A.C.; Crocker, P.; McLellan, A.D. CD169 mediates the capture of exosomes in spleen and lymph node. Blood 2014, 123, 208–216. [Google Scholar] [CrossRef]
- Muhsin-Sharafaldine, M.R.; Saunderson, S.C.; Dunn, A.C.; McLellan, A.D. Melanoma growth and lymph node metastasis is inde-pendent of host CD169 expression. Biochem. Biophys. Res. Commun. 2017, 486, 965–970. [Google Scholar] [CrossRef]
- Pucci, F.; Garris, C.; Lai, C.P.; Newton, A.; Pfirschke, C.; Engblom, C.; Alvarez, D.; Sprachman, M.; Evavold, C.; Magnuson, A.; et al. SCS macrophages suppress melanoma by restricting tumor-derived vesicle-B cell interactions. Science 2016, 352, 242–246. [Google Scholar] [CrossRef] [Green Version]
- Tacconi, C.; Commerford, C.D.; Dieterich, L.C.; Schwager, S.; He, Y.; Ikenberg, K.; Friebel, E.; Becher, B.; Tugues, S.; Detmar, M. CD169+ lymph node macrophages have protective functions in mouse breast cancer metastasis. Cell Rep. 2021, 35. [Google Scholar] [CrossRef]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef]
- Mills, C.D. M1 and M2 Macrophages: Oracles of Health and Disease. Crit. Rev. Immunol. 2012, 32, 463–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Xia, Y.; Qiu, C.-H. Functions of CD169 positive macrophages in human diseases (Review). Biomed. Rep. 2020, 14, 1. [Google Scholar] [CrossRef]
- Hashimoto, D.; Chow, A.; Noizat, C.; Teo, P.; Beasley, M.B.; Leboeuf, M.; Becker, C.D.; See, P.; Price, J.; Lucas, D.; et al. Tissue-resident macrophages self-maintain locally throughout adult life with minimal contribution from circulating monocytes. Immunity 2013, 38, 792–804. [Google Scholar] [CrossRef] [Green Version]
- Ginhoux, F.; Greter, M.; Leboeuf, M.; Nandi, S.; See, P.; Gokhan, S.; Mehler, M.F.; Conway, S.J.; Ng, L.G.; Stanley, E.R.; et al. Fate Mapping Analysis Reveals That Adult Microglia Derive from Primitive Macrophages. Science 2010, 330, 841–845. [Google Scholar] [CrossRef] [Green Version]
- Perdiguero, E.G.; Klapproth, K.; Schulz, C.; Busch, K.; Azzoni, E.; Crozet, L.; Garner, H.; Trouillet, C.; de Bruijn, M.F.; Geissmann, F.; et al. Tissue-resident macrophages originate from yolk-sac-derived erythro-myeloid progenitors. Nature 2015, 518, 547–551. [Google Scholar] [CrossRef] [PubMed]
- Blériot, C.; Chakarov, S.; Ginhoux, F. Determinants of Resident Tissue Macrophage Identity and Function. Immunity 2020, 52, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Ngo, V.N.; Cornall, R.J.; Cyster, J.G. Splenic T Zone Development Is B Cell Dependent. J. Exp. Med. 2001, 194, 1649–1660. [Google Scholar] [CrossRef]
- Phan, T.G.; Green, J.A.; Gray, E.E.; Xu, Y.; Cyster, J.G. Immune complex relay by subcapsular sinus macrophages and noncognate B cells drives antibody affinity maturation. Nat. Immunol. 2009, 10, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Komohara, Y.; Ohnishi, K.; Takeya, M. Possible functions of CD169-positive sinus macrophages in lymph nodes in anti-tumor immune responses. Cancer Sci. 2017, 108, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnishi, K.; Komohara, Y.; Saito, Y.; Miyamoto, Y.; Watanabe, M.; Baba, H.; Takeya, M. CD169-positive macrophages in regional lymph nodes are associated with a favorable prognosis in patients with colorectal carcinoma. Cancer Sci. 2013, 104, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Ohnishi, K.; Miyashita, A.; Nakahara, S.; Fujiwara, Y.; Horlad, H.; Motoshima, T.; Fukushima, S.; Jinnin, M.; Ihn, H.; et al. Prognostic Significance of CD169+ Lymph Node Sinus Macrophages in Patients with Malignant Melanoma. Cancer Immunol. Res. 2015, 3, 1356–1363. [Google Scholar] [CrossRef] [Green Version]
- Ohnishi, K.; Yamaguchi, M.; Erdenebaatar, C.; Saito, F.; Tashiro, H.; Katabuchi, H.; Takeya, M.; Komohara, Y. Prognostic significance of CD 169-positive lymph node sinus macrophages in patients with endometrial carcinoma. Cancer Sci. 2016, 107, 846–852. [Google Scholar] [CrossRef]
- Asano, T.; Ohnishi, K.; Shiota, T.; Motoshima, T.; Sugiyama, Y.; Yatsuda, J.; Kamba, T.; Ishizaka, K.; Komohara, Y. CD 169-positive sinus macrophages in the lymph nodes determine bladder cancer prognosis. Cancer Sci. 2018, 109, 1723–1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumamoto, K.; Tasaki, T.; Ohnishi, K.; Shibata, M.; Shimajiri, S.; Harada, M.; Komohara, Y.; Nakayama, T. CD169 Expression on Lymph Node Macrophages Predicts in Patients with Gastric Cancer. Front. Oncol. 2021, 11, 824. [Google Scholar] [CrossRef]
- Takeya, H.; Shiota, T.; Yagi, T.; Ohnishi, K.; Baba, Y.; Miyasato, Y.; Kiyozumi, Y.; Yoshida, N.; Takeya, M.; Baba, H.; et al. High CD169 expression in lymph node macrophages predicts a favorable clinical course in patients with esophageal cancer. Pathol. Int. 2018, 68, 685–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strömvall, K.; Sundkvist, K.; Ljungberg, B.; Halin Bergström, S.; Bergh, A. Reduced number of CD169+ macrophages in pre-metastatic regional lymph nodes is associated with subsequent metastatic disease in an animal model and with poor out-come in prostate cancer patients. Prostate 2017, 77, 1468–1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björk Gunnarsdottir, F.; Auoja, N.; Bendahl, P.O.; Rydén, L.; Fernö, M.; Leandersson, K. Co-localization of CD169+ macrophages and cancer cells in lymph node metastases of breast cancer patients is linked to improved prognosis and PDL1 expression. Onco-immunology 2020, 9, 1848067. [Google Scholar]
- Shiota, T.; Miyasato, Y.; Ohnishi, K.; Yamamoto-Ibusuki, M.; Yamamoto, Y.; Iwase, H.; Takeya, M.; Komohara, Y. The Clinical Significance of CD169-Positive Lymph Node Macrophage in Patients with Breast Cancer. PLoS ONE 2016, 11, e0166680. [Google Scholar] [CrossRef] [Green Version]
- Topf, M.C.; Harshyne, L.; Tuluc, M.; Mardekian, S.; Vimawala, S.; Cognetti, D.M.; Curry, J.M.; Rodeck, U.; Luginbuhl, A. Loss of CD169+ Subcapsular Macrophages during Metastatic Spread of Head and Neck Squamous Cell Carcinoma. Otolaryngol. Neck Surg. 2019, 161, 67–73. [Google Scholar] [CrossRef]
- Strömvall, K.; Thysell, E.; Bergström, S.H.; Bergh, A. Aggressive rat prostate tumors reprogram the benign parts of the prostate and regional lymph nodes prior to metastasis. PLoS ONE 2017, 12, e0176679. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Tumor Type | Link to Prognosis | Link to TIL | Comments | Reference |
|---|---|---|---|---|
| Colorectal cancer | better OS | yes | High density of CD169+ SMs were observed in cases with T1/2 stage or without lymph node metastasis. CD169 expression in human monocyte-derived macrophages was increased by type-I interferons (IFNs). | [39] |
| Melanoma | better OS | yes | IFN-alpha secreted from plasmacytoid dendritic cells was suggested to link to CD169 expression. | [40] |
| Endometrial cancer | better OS | yes | High density of CD169+ SMs were correlated with high density of natural killer cells more significantly than that of TILs. | [41] |
| Bladder cancer | better OS | yes | High density of CD169+ SMs were observed in cases with T1/2 stage. | [42] |
| Esophageal cancer | better OS | yes | The density of CD169+ SMs were higher in female than male. Significant correlation between CD169+ SMs and TILs were seen in cases with neoadjuvant therapy. CD169+ SMs partially expressed IDO1. | [44] |
| Breast cancer | not significant | yes | High density of CD169+ SMs was correlated with high density of TILs in cases with high Ki67 index. | [47] |
| Breast cancer | better PFS | not done | High density of CD169+ SMs were seen in cases with low tumor size, and correlated with PD-L1 expression both in primary tumor and metastatic tumor. Co-expression of CD169 and PD-L1 was seen in cases of younger age. | [46] |
| Gastric cancer | better OS and PFS | yes | High density of CD169+ SMs was associated with better OS in cases with advanced cancer or without metastasis and correlated to better PFS in cases with diffuse type or high tumor-stroma ratio. | [43] |
| Prostate cancer | better OS | not done | The significance of CD169+ SMs was suggested by rat prostate cancer metastatic model. | [45] |
| Head and neck cancer | not done | not done | High density of CD169+ SMs were observed in lymph node without metastasis. | [48] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komohara, Y.; Anami, T.; Asano, K.; Fujiwara, Y.; Yatsuda, J.; Kamba, T. Anti-Cancer Immune Reaction and Lymph Node Macrophage; A Review from Human and Animal Studies. Immuno 2021, 1, 223-230. https://0-doi-org.brum.beds.ac.uk/10.3390/immuno1030014
Komohara Y, Anami T, Asano K, Fujiwara Y, Yatsuda J, Kamba T. Anti-Cancer Immune Reaction and Lymph Node Macrophage; A Review from Human and Animal Studies. Immuno. 2021; 1(3):223-230. https://0-doi-org.brum.beds.ac.uk/10.3390/immuno1030014
Chicago/Turabian StyleKomohara, Yoshihiro, Toshiki Anami, Kenichi Asano, Yukio Fujiwara, Junji Yatsuda, and Tomomi Kamba. 2021. "Anti-Cancer Immune Reaction and Lymph Node Macrophage; A Review from Human and Animal Studies" Immuno 1, no. 3: 223-230. https://0-doi-org.brum.beds.ac.uk/10.3390/immuno1030014