
Antennal and Behavioral Responses of Drosophila suzukii to Volatiles from a Non-Crop Host, Osyris wightiana


 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Collection of Fruit Volatiles
2.3. Coupled Gas Chromatography-Electroantennographic Detection (GC-EAD)
2.4. Chemical Analyses
2.5. Chemicals
2.6. Y-Tube Olfactometer Bioassays
2.7. Field-Trapping Experiments
2.8. Statistical Analyses
3. Results
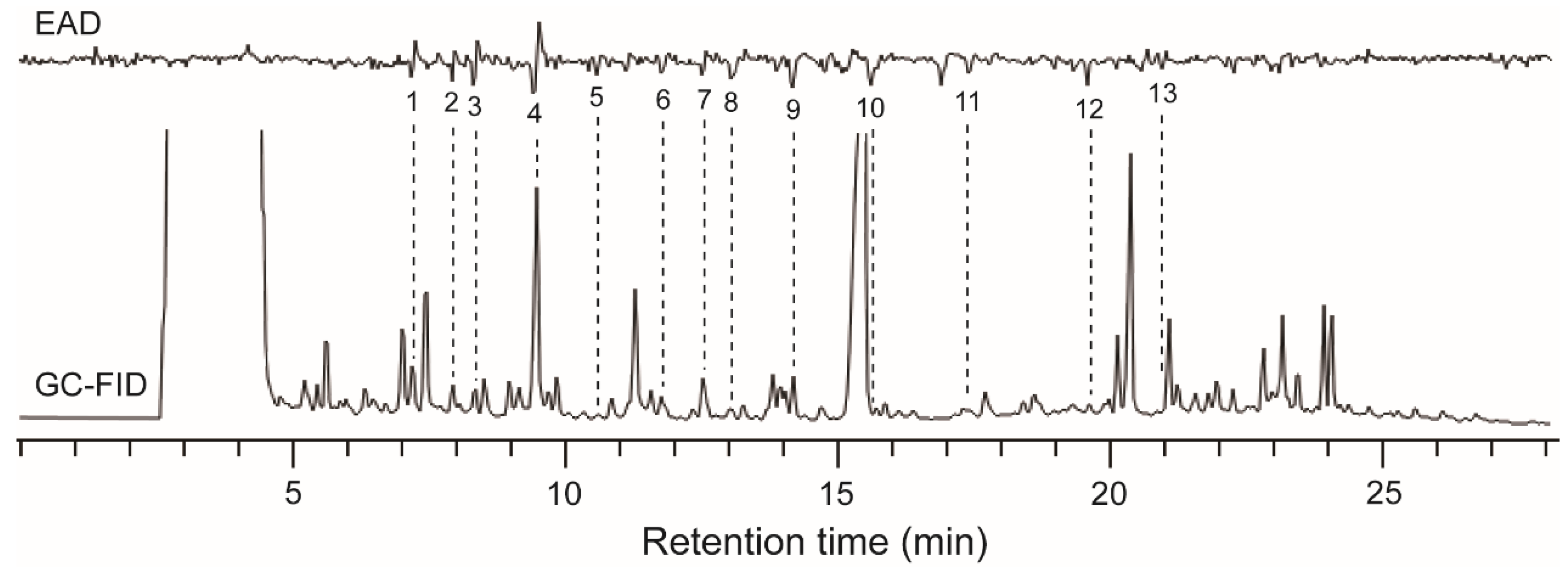
3.1. Bioactive Volatile Compounds Released by O. wightiana Fruits
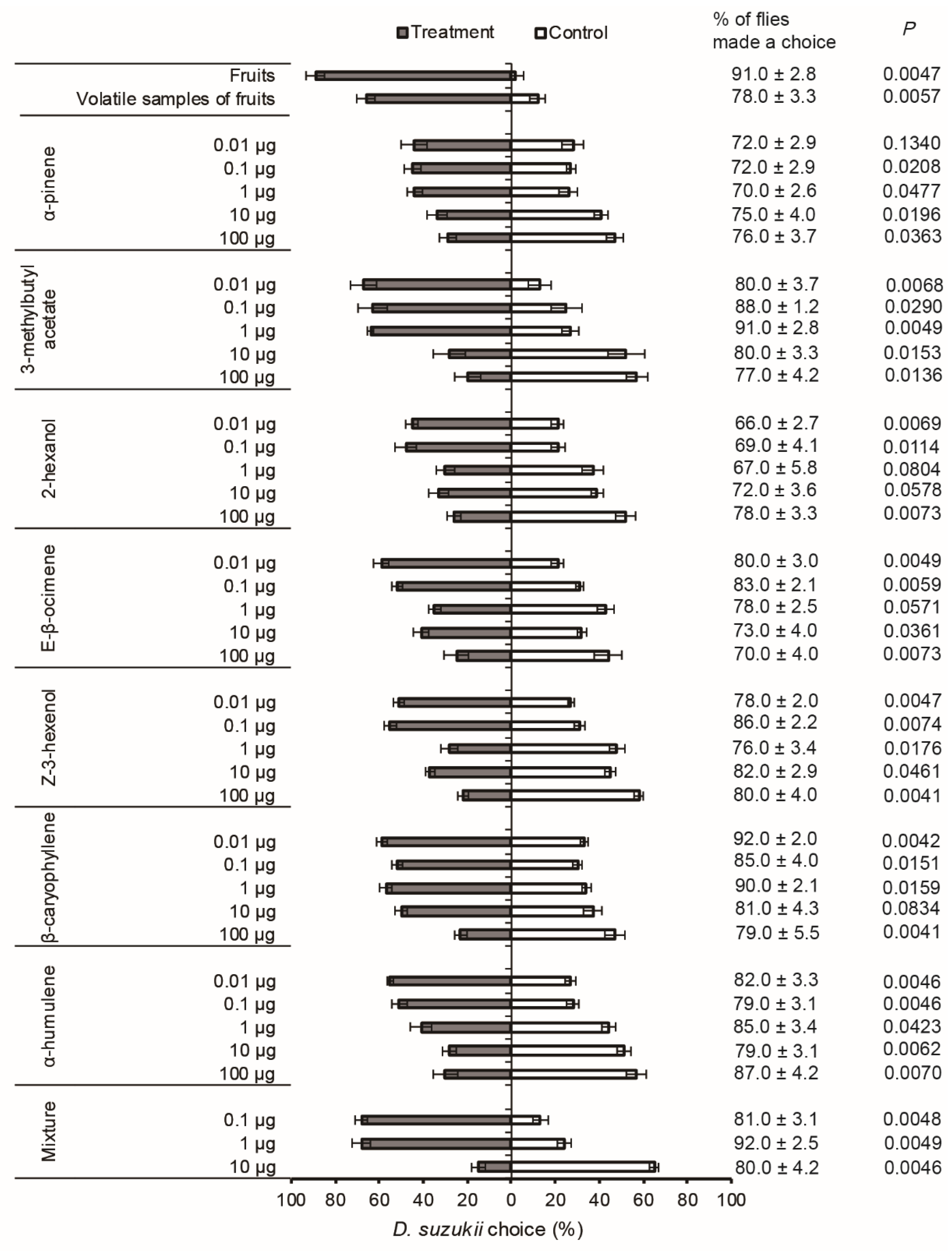
3.2. Dual-Choice Olfactometer Bioassays
3.3. Field-Trapping Experiment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cloonan, K.R.; Abraham, J.; Angeli, S.; Syed, Z.; Rodriguez-Saona, C. Advances in the chemical ecology of the spotted wing drosophila (Drosophila suzukii) and its applications. J. Chem. Ecol. 2018, 44, 922–939. [Google Scholar] [CrossRef]
- Tait, G.; Grassi, A.; Pfab, F.; Crava, C.M.; Dalton, D.T.; Magarey, R.; Ometto, L.; Vezzulli, S.; Rossi-Stacconi, M.V.; Gottardello, A.; et al. Large-scale spatial dynamics of Drosophila suzukii in Trentino, Italy. J. Pest Sci. 2018, 91, 1213–1224. [Google Scholar] [CrossRef]
- Bellamy, D.E.; Sisterson, M.S.; Walse, S.S. Quantifying host potentials: Indexing postharvest fresh fruits for spotted wing drosophila, Drosophila suzukii. PLoS ONE 2013, 8, 10. [Google Scholar] [CrossRef] [Green Version]
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive pest of ripening soft fruit expanding its geographic range and damage potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Lee, J.C.; Bruck, D.J.; Curry, H.; Edwards, D.; Haviland, D.R.; Van Steenwyk, R.A.; Yorgey, B.M. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosophila suzukii. Pest Manag. Sci. 2011, 67, 1358–1367. [Google Scholar] [CrossRef]
- Ballman, E.S.; Drummond, F.A. Infestation of wild fruit by Drosophila suzukii surrounding maine wild blueberry fields. J. Agric. Urban Entomol. 2017, 33, 61–70. [Google Scholar] [CrossRef]
- Weißinger, L.; Schrieber, K.; Breuer, M.; Müller, C. Influences of blackberry margins on population dynamics of Drosophila suzukii and grape infestation in adjacent vineyards. J. Appl. Entomol. 2019, 143, 802–812. [Google Scholar] [CrossRef]
- Rodriguez-Saona, C.; Cloonan, K.R.; Sanchez-Pedraza, F.; Zhou, Y.; Giusti, M.M.; Benrey, B. Differential susceptibility of wild and cultivated blueberries to an invasive frugivorous pest. J. Chem. Ecol. 2019, 45, 286–297. [Google Scholar] [CrossRef]
- Lee, J.C.; Dreves, A.J.; Cave, A.M.; Kawai, S.; Isaacs, R.; Miller, J.C.; Van Timmeren, S.; Bruck, D.J. Infestation of wild and ornamental noncrop fruits by Drosophila suzukii (Diptera: Drosophilidae). Ann. Entomol. Soc. Am. 2015, 108, 117–129. [Google Scholar] [CrossRef]
- Elsensohn, J.E.; Loeb, G.M. Non-crop host sampling yields insights into small-scale population dynamics of Drosophila suzukii (Matsumura). Insects 2018, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenis, M.; Tonina, L.; Eschen, R.; Sluis, B.V.D.; Sancassani, M.; Mori, N.; Haye, T.; Helsen, H. Non-crop plants used as hosts by Drosphila suzukii in Europe. J. Pest Sci. 2016, 89, 735–748. [Google Scholar] [CrossRef] [Green Version]
- Poyet, M.; Le Roux, V.; Gibert, P.; Meirland, A.; Prévost, G.; Eslin, P.; Chabrerie, O. The wide potential trophic niche of the asiatic fruit fly Drosophila suzukii: The key of its invasion success in temperate Europe? PLoS ONE 2015, 10, e0142785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arno, J.; Sola, M.; Riudavets, J.; Gabarra, R. Population dynamics, non-crop hosts, and fruit susceptibility of Drosophila suzukii in Northeast Spain. J. Pest Sci. 2016, 89, 713–723. [Google Scholar] [CrossRef]
- Keesey, I.W.; Knaden, M.; Hansson, B.S. Olfactory specialization in Drosophila suzukii supports an ecological shift in host preference from rotten to fresh fruit. J. Chem. Ecol. 2015, 41, 121–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burrack, H.J.; Fernandez, G.E.; Spivey, T.; Kraus, D.A. Variation in selection and utilization of host crops in the field and laboratory by Drosophila suzukii Matsumara (Diptera: Drosophilidae), an invasive frugivore. Pest Manage. Sci. 2013, 69, 1173–1180. [Google Scholar] [CrossRef]
- Liu, Y.; Dong, W.X.; Zhang, F.; Kenis, M.; Griepink, F.; Zhang, J.P.; Chen, L.; Xiao, C. Identification of active components from volatiles of Chinese bayberry, Myrica rubra attractive to Drosophila suzukii. Arthropod-Plant Interact. 2018, 12, 435–442. [Google Scholar] [CrossRef]
- Abraham, J.; Zhang, A.; Angeli, S.; Abubeker, S.; Michel, C.; Feng, Y.; Rodriguezsaona, C. Behavioral and antennal responses of Drosophila suzukii (Diptera: Drosophilidae) to volatiles from fruit extracts. Environ. Entomol. 2015, 44, 356–367. [Google Scholar] [CrossRef] [Green Version]
- Revadi, S.; Eccher, F.; Mazzoni, V.; Alani, S.; Carlin, S.; Vrhovsek, U.; Anfora, G.; Ioriatti, C.; Altindisli, F.Ö.; Børve, J. Olfactory responses of Drosophila suzukii females to host plant volatiles. Physiol. Entomol. 2015, 40, 54–64. [Google Scholar] [CrossRef]
- Knolhoff, L.M.; Heckel, D.G. Behavioral assays for studies of host plant choices and adaptation in herbivorous insects. Annu. Rev. Entomol. 2014, 59, 263–278. [Google Scholar] [CrossRef]
- Cha, D.H.; Yee, W.L.; Goughnour, R.B.; Sim, S.B.; Powell, T.H.Q.; Feder, J.L.; Linn, C.E. Identification of host fruit volatiles from domestic apple (Malus domestica), native black hawthorn (Crataegus douglasii) and introduced ornamental hawthorn (C. monogyna) attractive to Rhagoletis pomonella flies from the western United States. J. Chem. Ecol. 2012, 38, 319–329. [Google Scholar] [CrossRef] [Green Version]
- Kang, W.; Li, Y.; Xu, Y.; Jiang, W.; Tao, Y. Characterization of aroma compounds in Chinese bayberry (Myrica rubra Sieb. et Zucc.) by gas chromatography mass spectrometry (GC-MS) and olfactometry (GC-O). J. Food Sci. 2012, 77, C1030–C1035. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Chen, J.; Chen, S.; Wu, D.; Liu, D.; Ye, X. Characterization of aroma-active volatiles in three Chinese bayberry (Myrica rubra) cultivars using GC-MS-olfactometry and an electronic nose combined with principal component analysis. Food Res. Int. 2015, 72, 8–15. [Google Scholar] [CrossRef]
- Zhu, J.; Xiao, Z. Characterization of the key aroma compounds in peach by gas chromatography-olfactometry, quantitative measurements and sensory analysis. Eur. Food Res. Technol. 2019, 245, 129–141. [Google Scholar] [CrossRef]
- Wu, Y.S.; Zhang, W.W.; Yu, W.J.; Zhao, L.P.; Song, S.R.; Xu, W.P.; Zhang, C.X.; Ma, C.; Wang, L.; Wang, S.P. Study on the volatile composition of table grapes of three aroma types. LWT Food Sci. Technol. 2019, 115, 9. [Google Scholar] [CrossRef]
- Perestrelo, R.; Silva, C.; Silva, P.; Medina, S.; Camara, J.S. Differentiation of fresh and processed fruit juices using volatile composition. Molecules 2019, 24, 974. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Lu, J.; He, Z.; Zhang, F.; Zhang, S.; Zhang, H. Investigations into the production of volatile compounds in Korla fragrant pears (Pyrus sinkiangensis Yu). Food Chem. 2020, 302, 1–6. [Google Scholar] [CrossRef]
- Feng, J.R.; Xi, W.P.; Li, W.H.; Liu, H.N.; Liu, X.F.; Lu, X.Y. Volatile characterization of major apricot cultivars of Southern Xinjiang region of China. J. Am. Soc. Hort. Sci. 2015, 140, 466–471. [Google Scholar] [CrossRef] [Green Version]
- Semmelhack, J.L.; Wang, J.W. Select Drosophila glomeruli mediate innate olfactory attraction and aversion. Nature 2009, 459, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Stensmyr, M.C.; Giordano, E.; Balloi, A.; Angioy, A.M.; Hansson, B.S. Novel natural ligands for Drosophila olfactory receptor neurons. J. Exp. Biol. 2003, 206, 715–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleiber, J.R.; Unelius, C.R.; Lee, J.C.; Suckling, D.M.; Qian, M.C.; Bruck, D.J. Attractiveness of fermentation and related products to spotted wing drosophila (Diptera: Drosophilidae). Environ. Entomol. 2014, 43, 439–447. [Google Scholar] [CrossRef]
- Rossi-Stacconi, M.V.; Kaur, R.; Mazzoni, V.; Ometto, L.; Grassi, A.; Gottardello, A.; Rota-Stabelli, O.; Anfora, G. Multiple lines of evidence for reproductive winter diapause in the invasive pest Drosophila suzukii: Useful clues for control strategies. J. Pest Sci. 2016, 89, 689–700. [Google Scholar] [CrossRef]
- Holland, J.; Fahrig, L. Effect of woody borders on insect density and diversity in crop fields: A landscape-scale analysis. Agric., Ecosyst. Environ. 2000, 78, 115–122. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Moonen, A.C. Field margins in northern Europe: Their functions and interactions with agriculture. Agric. Ecosyst. Environ. 2002, 89, 5–21. [Google Scholar] [CrossRef]
- Cini, A.; Ioriatti, C.; Anfora, G. A review of the invasion of Drosophila suzukii in Europe and a draft research agenda for integrated pest management. Bull. Insectol. 2012, 65, 149–160. [Google Scholar]
- Wang, X.-G.; Stewart, T.J.; Biondi, A.; Chavez, B.A.; Ingels, C.; Caprile, J.; Grant, J.A.; Walton, V.M.; Daane, K.M. Population dynamics and ecology of Drosophila suzukii in Central California. J. Pest Sci. 2016, 89, 701–712. [Google Scholar] [CrossRef]
- Klick, J.; Yang, W.Q.; Walton, V.M.; Dalton, D.T.; Hagler, J.R.; Dreves, A.J.; Lee, J.C.; Bruck, D.J. Distribution and activity of Drosophila suzukii in cultivated raspberry and surrounding vegetation. J. Appl. Entomol. 2016, 140, 37–46. [Google Scholar] [CrossRef]
- Lasa, R.; Toledo-Hernández, R.A.; Rodríguez, D.; Williams, T. Raspberry as a source for the development of Drosophila suzukii attractants: Laboratory and commercial polytunnel trials. Insects 2019, 10, 137. [Google Scholar] [CrossRef] [Green Version]
- Cai, P.; Yi, C.; Zhang, Q.; Zhang, H.; Lin, J.; Song, X.; Yang, J.; Wang, B.; Ji, Q.; Chen, J. Evaluation of protein bait manufactured from brewery yeast waste for controlling Drosophila suzukii (Diptera: Drosophilidae). J. Econ. Entomol. 2018, 1–10. [Google Scholar] [CrossRef]
- Cha, D.H.; Adams, T.; Werle, C.T.; Sampson, B.J.; Adamczyk, J.J., Jr.; Rogg, H.; Landolt, P.J. A four-component synthetic attractant for Drosophila suzukii (Diptera: Drosophilidae) isolated from fermented bait headspace. Pest Manag. Sci. 2014, 70, 324–331. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Peak No. | Compounds | RI a | Relative Content (ng/µL) | Percentage b (%) | |
|---|---|---|---|---|---|
| Exp. | Lit. | ||||
| 1 | α-Pinene | 1015 | 989–1052 | 2.7 ± 0.9 | 0.38 ± 0.13 |
| 2 | Unidentified | 1043 | - | 1.5 ± 0.3 | 0.21 ± 0.04 |
| 3 | Unidentified | 1063 | - | 1.3 ± 0.5 | 0.18 ± 0.07 |
| 4 | 3-methylbutyl acetate | 1119 | 1102–1140 | 9.0 ± 1.6 | 1.27 ± 0.23 |
| 5 | Unidentified | 1167 | - | 0.4 ± 0.0 | 0.05 ± 0.00 |
| 6 | 2-Hexanol | 1216 | 1192–1217 | 1.5 ± 0.5 | 0.21 ± 0.07 |
| 7 | Unidentified | 1246 | - | 0.7 ± 0.1 | 0.38 ± 0.05 |
| 8 | Unidentified | 1269 | - | 0.7 ± 0.1 | 0.10 ± 0.01 |
| 9 | E-β-ocimene | 1301 | 1232–1290 | 2.2 ± 0.7 | 0.31 ± 0.10 |
| 10 | Z-3-hexenol | 1369 | 1351–1390 | 1.2 ± 0.3 | 0.17 ± 0.04 |
| 11 | Unidentified | 1422 | - | 0.2 ± 0.1 | 0.03 ± 0.02 |
| 12 | β-Caryophyllene | 1556 | 1556–1615 | 0.6 ± 0.2 | 0.08 ± 0.03 |
| 13 | α-Humulene | 1676 | 1623–1705 | 0.4 ± 0.1 | 0.05 ± 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Cui, Z.; Shi, M.; Kenis, M.; Dong, W.; Zhang, F.; Zhang, J.; Xiao, C.; Chen, L. Antennal and Behavioral Responses of Drosophila suzukii to Volatiles from a Non-Crop Host, Osyris wightiana. Insects 2021, 12, 166. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020166
Liu Y, Cui Z, Shi M, Kenis M, Dong W, Zhang F, Zhang J, Xiao C, Chen L. Antennal and Behavioral Responses of Drosophila suzukii to Volatiles from a Non-Crop Host, Osyris wightiana. Insects. 2021; 12(2):166. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020166
Chicago/Turabian StyleLiu, Yan, Zhihao Cui, Mi Shi, Marc Kenis, Wenxia Dong, Feng Zhang, Jinping Zhang, Chun Xiao, and Li Chen. 2021. "Antennal and Behavioral Responses of Drosophila suzukii to Volatiles from a Non-Crop Host, Osyris wightiana" Insects 12, no. 2: 166. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020166