
Unexpected Effects of Local Management and Landscape Composition on Predatory Mites and Their Food Resources in Vineyards

 ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
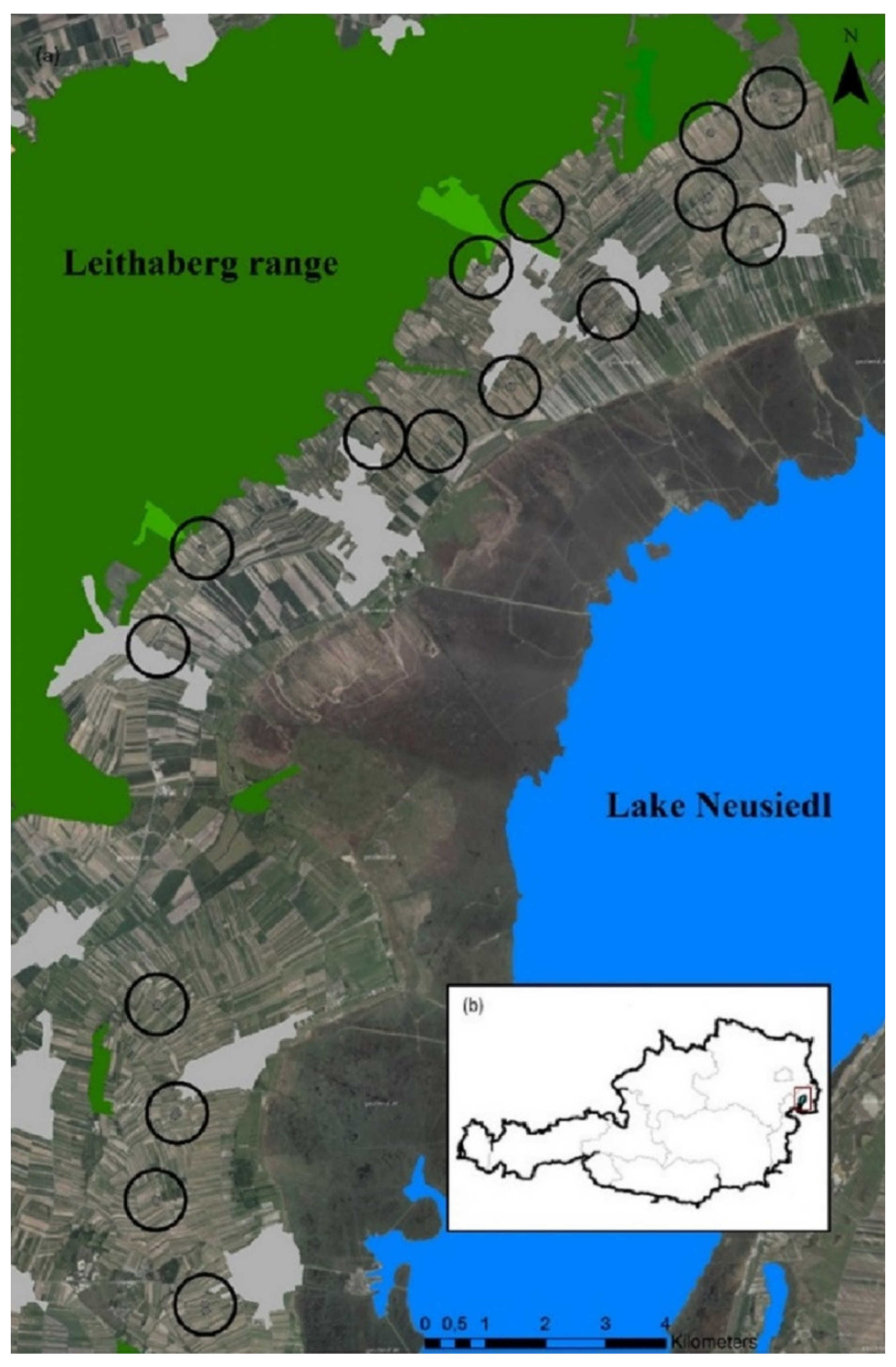
2.1. Study Sites
2.2. Mite Sampling
2.3. Pollen Sampling
2.4. Vegetation Survey
2.5. Landscape Survey
2.6. Area-Related Acute Pesticide Contact Toxicity Loading (aAPTLc)
2.7. Data Analysis
3. Results
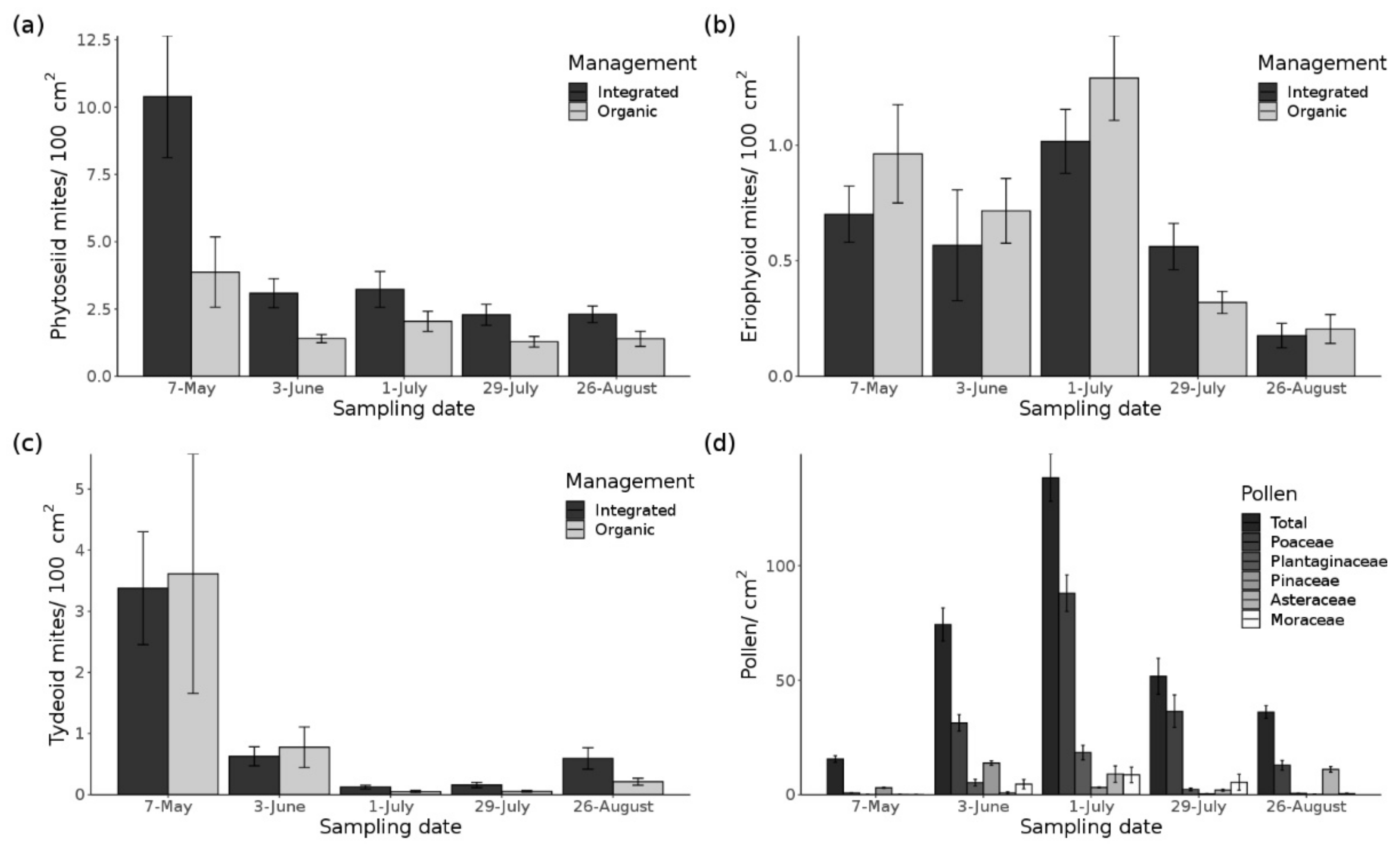
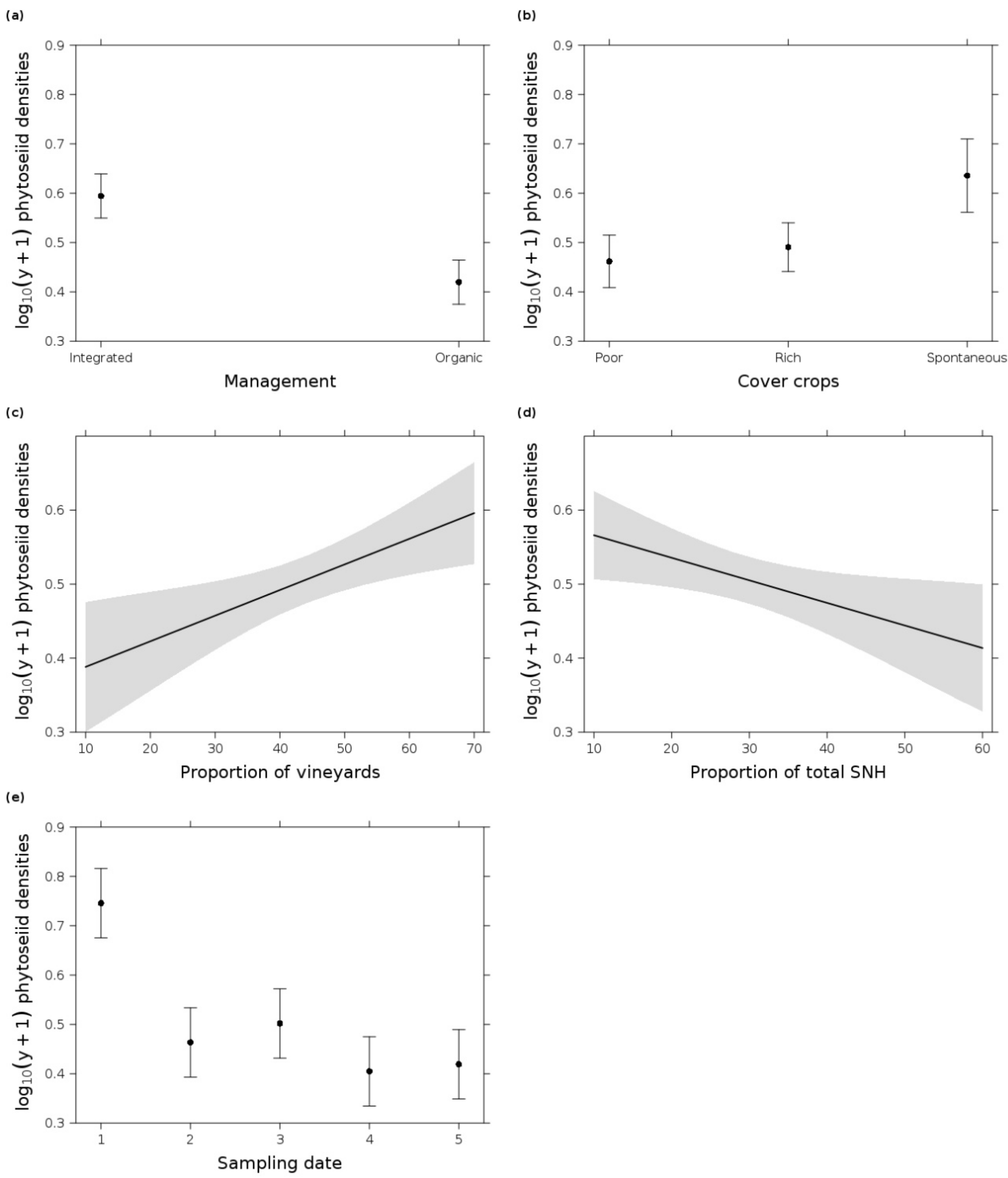
3.1. Phytoseiid Mites
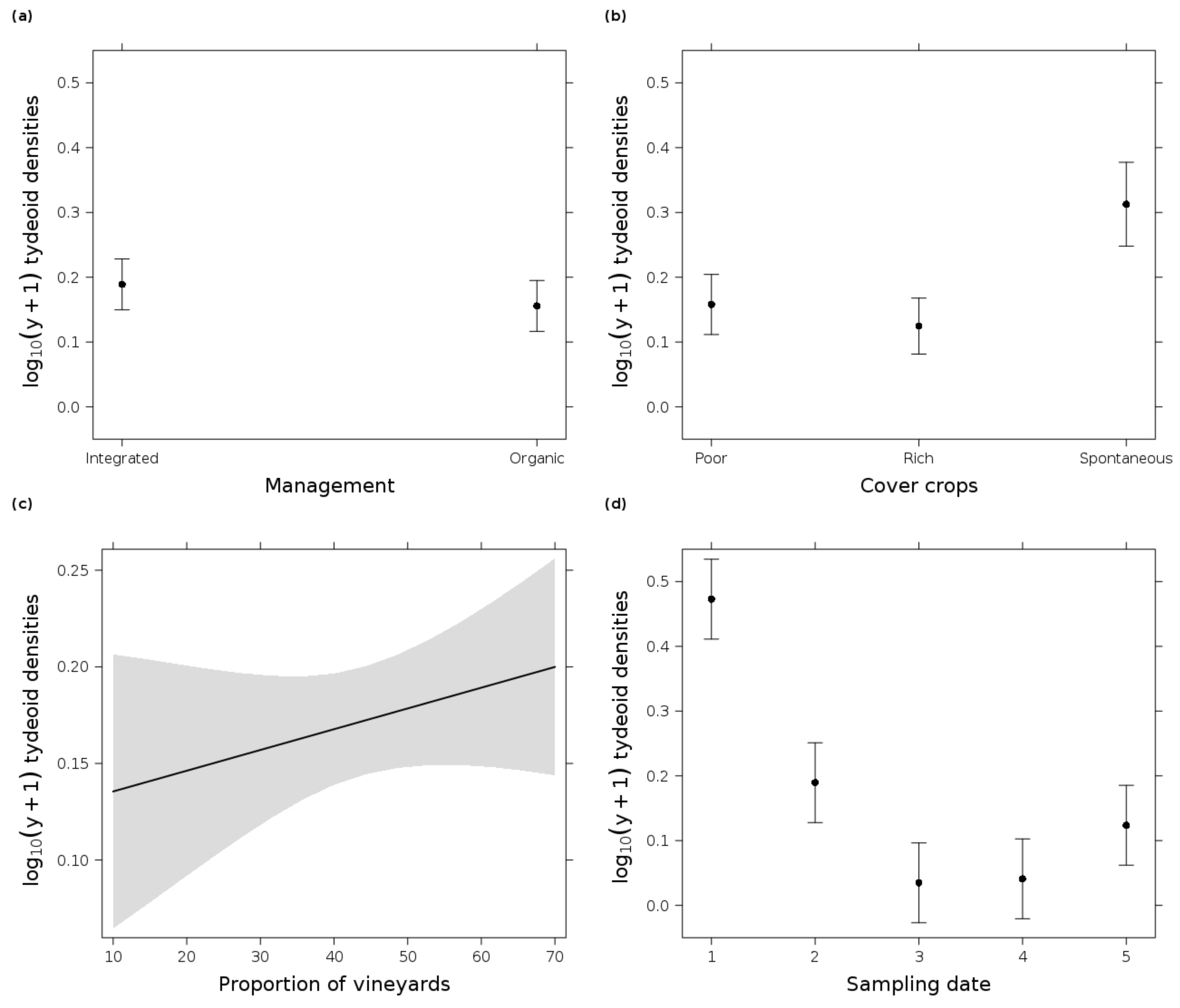
3.2. Tydeoid Mites
3.3. Phytophagous Mites
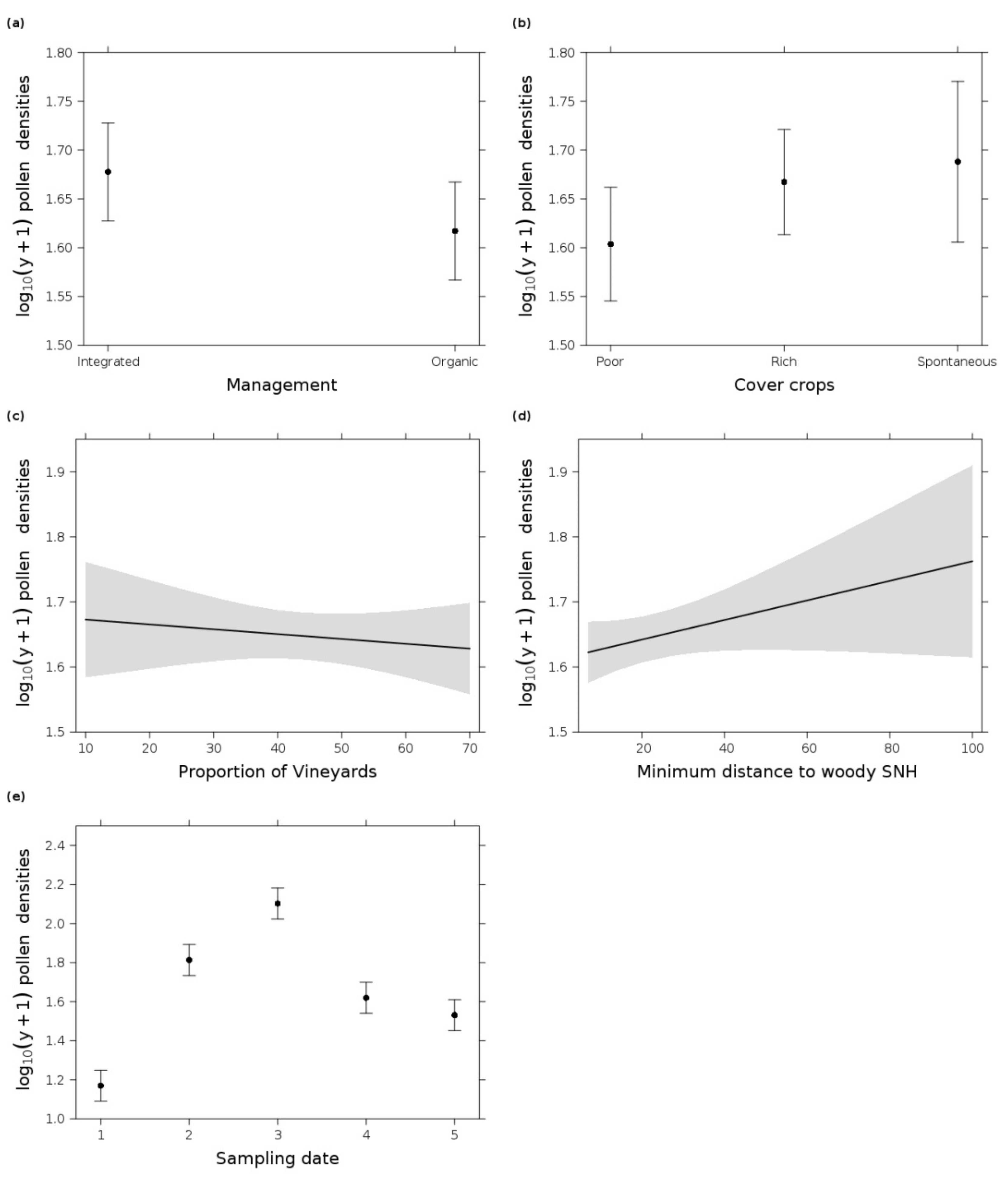
3.4. Pollen
4. Discussion
4.1. Integrated Management and Low Pesticide Use Increased Predatory Mite Densities
4.2. The Cover Crop Type Influenced the Mite Populations
4.3. Predatory Mite Densities Benefited from Surrounding Vineyards
4.4. Low Diversity of Phytoseiids
4.5. Factors Influencing Pollen Availability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Altieri, M.A. How best can we use biodiversity in agroecosystems? Outlook Agric. 1991, 20, 15–23. [Google Scholar] [CrossRef]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat Management to Conserve Natural Enemies of Arthropod Pests in Agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Haddad, N.M.; Crutsinger, G.M.; Gross, K.; Haarstad, J.; Knops, J.M.H.; Tilman, D. Plant species loss decreases arthropod diversity and shifts trophic structure. Ecol. Lett. 2009, 12, 1029–1039. [Google Scholar] [CrossRef]
- Root, R.B. Organization of a Plant-Arthropod Association in Simple and Diverse Habitats: The Fauna of Collards (Brassica oleracea). Ecol. Monogr. 1973, 43, 95–124. [Google Scholar] [CrossRef]
- Russell, E.P. Enemies Hypothesis: A Review of the Effect of Vegetational Diversity on Predatory Insects and Parasitoids. Environ. Entomol. 1989, 18, 590–599. [Google Scholar] [CrossRef] [Green Version]
- O’Rourke, M.E.; Petersen, M.J. Extending the ‘resource concentration hypothesis’ to the landscape-scale by considering dispersal mortality and fitness costs. Agric. Ecosyst. Environ. 2017, 249, 1–3. [Google Scholar] [CrossRef]
- Tscharntke, T.; Karp, D.S.; Chaplin-Kramer, R.; Batáry, P.; DeClerck, F.; Gratton, C.; Hunt, L.; Ives, A.; Jonsson, M.; Larsen, A.; et al. When natural habitat fails to enhance biological pest control—Five hypotheses. Biol. Conserv. 2016, 204, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Wan, N.F.; Ji, X.Y.; Jiang, J.X. Testing the enemies hypothesis in peach orchards in two different geographic areas in eastern China: The role of ground cover vegetation. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novais, S.M.A.; Macedo-Reis, L.E.; Neves, F.S. Predatory beetles in cacao agroforestry systems in Brazilian Atlantic forest: A test of the natural enemy hypothesis. Agrofor. Syst. 2017, 91, 201–209. [Google Scholar] [CrossRef]
- Harmon, J.P.; Hladilek, E.E.; Hinton, J.L.; Stodola, T.J.; Andow, D.A. Herbivore response to vegetational diversity: Spatial interaction of resources and natural enemies. Popul. Ecol. 2003, 45, 75–81. [Google Scholar] [CrossRef]
- Björkman, M.; Hambäck, P.A.; Hopkins, R.J.; Rämert, B. Evaluating the enemies hypothesis in a clover-cabbage intercrop: Effects of generalist and specialist natural enemies on the turnip root fly (Delia floralis). Agric. For. Entomol. 2010, 12, 123–132. [Google Scholar] [CrossRef]
- Kheirodin, A.; Cárcamo, H.A.; Costamagna, A.C. Contrasting effects of host crops and crop diversity on the abundance and parasitism of a specialist herbivore in agricultural landscapes. Landsc. Ecol. 2020, 35, 1073–1087. [Google Scholar] [CrossRef]
- Miklasiewicz, T.J.; Hammond, R.B. Density of potato leafhopper (Homoptera: Cicadellidae) in response to soybean-wheat cropping systems. Environ. Entomol. 2001, 30, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Parsa, S.; Ccanto, R.; Rosenheim, J.A. Resource concentration dilutes a key pest in indigenous potato agriculture. Ecol. Appl. 2011, 21, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Karp, D.S.; Chaplin-Kramer, R.; Meehan, T.D.; Martin, E.A.; DeClerck, F.; Grab, H.; Gratton, C.; Hunt, L.; Larsen, A.E.; Martínez-Salinas, A.; et al. Crop pests and predators exhibit inconsistent responses to surrounding landscape composition. Proc. Natl. Acad. Sci. USA 2018, 115, E7863–E7870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paredes, D.; Rosenheim, J.A.; Chaplin-Kramer, R.; Winter, S.; Karp, D.S. Landscape simplification increases vineyard pest outbreaks and insecticide use. Ecol. Lett. 2020. [Google Scholar] [CrossRef] [PubMed]
- Vincent, C.; Isaacs, R.; Bostanian, N.J.; Lasnier, J. Principles of Arthropod Pest Management in Vineyards. In Arthropod Management in Vineyards: Pests, Approaches, and Future Directions; Bostanian, N.J., Vincent, C., Isaacs, R., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 1–16. ISBN 978-94-007-4032-7. [Google Scholar]
- Kottek, M.; Grieser, J.; Beck, C.; Rudolf, B.; Rubel, F. World map of the Köppen-Geiger climate classification updated. Meteorol. Z. 2006, 15, 259–263. [Google Scholar] [CrossRef]
- Kazakou, E.; Fried, G.; Richarte, J.; Gimenez, O.; Violle, C.; Metay, A. A plant trait-based response-and-effect framework to assess vineyard inter-row soil management. Bot. Lett. 2016, 163, 373–388. [Google Scholar] [CrossRef]
- Hall, R.M.; Penke, N.; Kriechbaum, M.; Kratschmer, S.; Jung, V.; Chollet, S.; Guernion, M.; Nicolai, A.; Burel, F.; Fertil, A.; et al. Vegetation management intensity and landscape diversity alter plant species richness, functional traits and community composition across European vineyards. Agric. Syst. 2020, 177. [Google Scholar] [CrossRef]
- Fried, G.; Cordeau, S.; Metay, A.; Kazakou, E. Relative importance of environmental factors and farming practices in shaping weed communities structure and composition in French vineyards. Agric. Ecosyst. Environ. 2019, 275, 1–13. [Google Scholar] [CrossRef]
- Winter, S.; Bauer, T.; Strauss, P.; Kratschmer, S.; Paredes, D.; Popescu, D.; Landa, B.; Guzmán, G.; Gómez, J.A.; Guernion, M.; et al. Effects of vegetation management intensity on biodiversity and ecosystem services in vineyards: A meta-analysis. J. Appl. Ecol. 2018, 55, 2484–2495. [Google Scholar] [CrossRef] [Green Version]
- Burgio, G.; Marchesini, E.; Reggiani, N.; Montepaone, G.; Schiatti, P.; Sommaggio, D. Habitat management of organic vineyard in Northern Italy: The role of cover plants management on arthropod functional biodiversity. Bull. Entomol. Res. 2016, 106, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Paiola, A.; Assandri, G.; Brambilla, M.; Zottini, M.; Pedrini, P.; Nascimbene, J. Exploring the potential of vineyards for biodiversity conservation and delivery of biodiversity-mediated ecosystem services: A global-scale systematic review. Sci. Total Environ. 2020, 706. [Google Scholar] [CrossRef] [PubMed]
- Kehinde, T.; Samways, M.J. Insect-flower interactions: Network structure in organic versus conventional vineyards. Anim. Conserv. 2014, 17, 401–409. [Google Scholar] [CrossRef]
- Geldenhuys, M.; Gaigher, R.; Pryke, J.S.; Samways, M.J. Diverse herbaceous cover crops promote vineyard arthropod diversity across different management regimes. Agric. Ecosyst. Environ. 2021, 307. [Google Scholar] [CrossRef]
- Sáenz-Romo, M.G.; Martínez-García, H.; Veas-Bernal, A.; Carvajal-Montoya, L.D.; Martínez-Villar, E.; Ibáñez-Pascual, S.; Marco-Mancebón, V.S.; Pérez-Moreno, I. Effect of ground-cover management on predatory mites (Acari: Phytoseiidae) in a Mediterranean vineyard. Vitis J. Grapevine Res. 2019, 58, 25–32. [Google Scholar] [CrossRef]
- Sáenz-Romo, M.G.; Veas-Bernal, A.; Martínez-García, H.; Campos-Herrera, R.; Ibáñez-Pascual, S.; Martínez-Villar, E.; Pérez-Moreno, I.; Marco-Mancebón, V.S. Ground cover management in a Mediterranean vineyard: Impact on insect abundance and diversity. Agric. Ecosyst. Environ. 2019, 283. [Google Scholar] [CrossRef]
- Vogelweith, F.; Thiéry, D. Cover crop differentially affects arthropods, but not diseases, occurring on grape leaves in vineyards. Aust. J. Grape Wine Res. 2017, 23, 426–431. [Google Scholar] [CrossRef]
- Eckert, M.; Mathulwe, L.L.; Gaigher, R.; Joubert-van der Merwe, L.; Pryke, J.S. Native cover crops enhance arthropod diversity in vineyards of the Cape Floristic Region. J. Insect Conserv. 2020, 24, 133–149. [Google Scholar] [CrossRef]
- Isaacs, R.; Saunders, M.C.; Bostanian, N.J. Pest Thresholds: Their Development and Use in Vineyards for Arthropod Management. In Arthropod Management in Vineyards: Pests, Approaches, and Future Directions; Bostanian, N.J., Vincent, C., Isaacs, R., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 17–36. ISBN 978-94-007-4032-7. [Google Scholar]
- Communication from the Commission to the European Parliament, the Council, the E. Economic and Social Committee and the Committee of the Regions. A Farm to Fork Strategy for a Fair, Healthy and Environmentally-Friendly Food System COM(2020) 381 Final. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:52020DC0381 (accessed on 7 September 2020).
- Muneret, L.; Thiéry, D.; Joubard, B.; Rusch, A. Deployment of organic farming at a landscape scale maintains low pest infestation and high crop productivity levels in vineyards. J. Appl. Ecol. 2018, 55, 1516–1525. [Google Scholar] [CrossRef]
- Duso, C.; Malagnini, V.; Paganelli, A.; Aldegheri, L.; Bottini, M.; Otto, S. Pollen availability and abundance of predatory phytoseiid mites on natural and secondary hedgerows. BioControl 2004, 49, 397–415. [Google Scholar] [CrossRef]
- Tixier, M.S.; Kreiter, S.; Auger, P. Colonization of vineyards by phytoseiid mites: Their dispersal patterns the plot and their fate. Exp. Appl. Acarol. 2000, 24, 191–211. [Google Scholar] [CrossRef]
- Duso, C.; Pozzebon, A.; Kreiter, S.; Tixier, M.S.; Candolfi, M.; Kreiter, S. Management of phytophagous mites in European vineyards. In Arthropod Management in Vineyards: Pests, Approaches, and Future Directions; Bostanian, N.J., Vincent, C., Isaacs, R., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 191–217. ISBN 9789400740327. [Google Scholar]
- Da Silva, G.L.; Metzelthin, M.H.; Da Silva, O.S.; Ferla, N.J. Catalogue of the mite family Tydeidae (Acari: Prostigmata) with the world key to the species. Zootaxa 2016, 4135, 1–68. [Google Scholar] [CrossRef] [PubMed]
- Schruft, G. Das Vorkommen von Milben aus der Familie Tydeidae (Acari) an Reben. Z. Angew. Entomol. 1972, 71, 124–133. [Google Scholar] [CrossRef]
- McMurtry, J.A.; De Moraes, G.J.; Sourassou, N.F. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol. 2013, 18, 297–320. [Google Scholar] [CrossRef] [Green Version]
- Boller, E.; Frey, B. Blühende Rebberge in der Ostschweiz: 1 Zur Bedeutung des Pollens für die Raubmilben. Schweizierische Z. Obs. Weinbau 1990, 126, 401–405. [Google Scholar]
- Pozzebon, A.; Duso, C. Grape downy mildew Plasmopara viticola, an alternative food for generalist predatory mites occurring in vineyards. Biol. Control. 2008, 45, 441–449. [Google Scholar] [CrossRef]
- Schausberger, P. Vergleichende Untersuchungen über den Einfluß unterschiedlicher Nahrung auf die Präimaginalentwicklung und die Reproduktion von Amblyseius aberrans Oud. und Amblyseius finlandicus Oud. (Acarina, Phytoseiidae). J. Appl. Entomol. 1992, 113, 476–486. [Google Scholar] [CrossRef]
- Engel, R.; Ohnesorge, B. Die Rolle von Ersatznahrung und Mikroklima im System Typhlodromus pyri Scheuten (Acari, Phytoseiidae)—Panonychus ulmi Koch (Acari, Tetranychidae) auf Weinreben I. Untersuchungen im Labor. J. Appl. Entomol. 1994, 118, 129–150. [Google Scholar] [CrossRef]
- Knop, N.F.; Hoy, M.A. Biology of a tydeid mite, Homeopronematus anconai (n. comb.) (Acari: Tydeidae), important in San Joaquin Valley vineyards. Hilgardia 1983, 51, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Hessein, N.A.; Perring, T.M. The importance of alternate foods for the mite Homeopronematus anconai (Acari: Tydeidae). Ann. Entomol. Soc. Am. 1988, 81, 488–492. [Google Scholar] [CrossRef]
- Wiedmer, U.; Boller, E. Blühende Rebberge in der Ostschweiz: 2. Zum Pollenangebot auf den Rebblättern. Schweiz. Z. Obs. Weinbau 1990, 426–431. [Google Scholar]
- Duso, C.; Malagnini, V.; Paganelli, A.; Aldegheri, L.; Bottini, M. Phytoseiid mites—Pollen relationships: Observations in a vineyard and the surrounding vegetation. In Acarid Phylogeny and Evolution: Adaptation in Mites and Ticks; Bernini, F., Nanelli, R., Nuzzace, G., de Lillo, E., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 373–387. ISBN 978-94-017-0611-7. [Google Scholar]
- Sabbatini Peverieri, G.; Simoni, S.; Goggioli, D.; Liguori, M.; Castagnoli, M. Effects of variety and management practices on mite species diversity in Italian vineyards. Bull. Insectol. 2009, 62, 53–60. [Google Scholar]
- Silva, D.E.; Nascimento, J.M.D.; Meira, A.D.A.; Johann, L.; Costa Corrêa, L.L.; Rodrigues, R.; Ferla, N.J.; Correâ, L.L.C.; Rodrigues, R.; Ferla, N.J. Phytoseiid mites under different vineyard managements in the subregions of Lima and Cávado of the Vinho Verde region in Portugal. Syst. Appl. Acarol. 2019, 24, 918–928. [Google Scholar] [CrossRef]
- Caprio, E.; Nervo, B.; Isaia, M.; Allegro, G.; Rolando, A. Organic versus conventional systems in viticulture: Comparative effects on spiders and carabids in vineyards and adjacent forests. Agric. Syst. 2015, 136, 61–69. [Google Scholar] [CrossRef]
- Muneret, L.; Auriol, A.; Thiéry, D.; Rusch, A. Organic farming at local and landscape scales fosters biological pest control in vineyards. Ecol. Appl. 2019, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nash, M.A.; Hoffmann, A.A.; Thomson, L.J. Identifying signature of chemical applications on indigenous and invasive nontarget arthropod communities in vineyards. Ecol. Appl. 2010, 20, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Gadino, A.N.; Walton, V.M.; Dreves, A.J. Impact of Vineyard Pesticides on a Beneficial Arthropod, Typhlodromus pyri (Acari: Phytoseiidae), in Laboratory Bioassays. J. Econ. Entomol. 2011, 104, 970–977. [Google Scholar] [CrossRef]
- Bernard, M.B.; Cole, P.; Kobelt, A.; Horne, P.A.; Altmann, J.; Wratten, S.D.; Yen, A.L. Reducing the impact of pesticides on biological control in Australian vineyards: Pesticide mortality and fecundity effects on an indicator species, the predatory mite Euseius victoriensis (Acari: Phytoseiidae). J. Econ. Entomol. 2010, 103, 2061–2071. [Google Scholar] [CrossRef] [PubMed]
- Gravesen, L. OECD Survey of National Pesticide Risk Indicators, 1999–2000. Available online: http://www.oecd.org/env/ehs/pesticides-biocides/1934154.pdf (accessed on 6 April 2020).
- Halberg, N.; Verschuur, G.; Goodlass, G. Farm level environmental indicators; are they useful? An overview of green accounting systems for European farms. Agric. Ecosyst. Environ. 2005, 105, 195–212. [Google Scholar] [CrossRef] [Green Version]
- Van Bol, V.; Claeys, S.; Debongnie, P.; Godfriaux, J.; Pussemier, L.; Steurbaut, W.; Maraite, H. Pesticide indicators. Pestic. Outlook 2003, 14, 159–163. [Google Scholar] [CrossRef]
- European Parliament; Council of the European Union Regulation (EC) No 1107/2009 of the European Parliament and of the Council of 21 October 2009 Concerning the Placing of Plant Protection Products on the Market and Repealing Council Directives 79/117/EEC and 91/414/EEC, COD 2006/0136. Available online: https://eur-lex.europa.eu/legal-content/EN/ALL/?uri=CELEX%3A32009R1107 (accessed on 10 September 2020).
- DiBartolomeis, M.; Kegley, S.; Mineau, P.; Radford, R.; Klein, K. An assessment of acute insecticide toxicity loading (AITL) of chemical pesticides used on agricultural land in the United States. PLoS ONE 2019, 14. [Google Scholar] [CrossRef] [Green Version]
- BOKU Institut für Pflanzenschutz Wetterstatistik. Available online: http://rebschutz.boku.ac.at (accessed on 20 March 2020).
- Amt der Burgenländischen Landesregierung Geodaten Burgenland. Available online: https://geodaten.bgld.gv.at/de/downloads.html (accessed on 23 October 2019).
- Boller, E. Eine einfache Ausschwemm-Methode zur schnellen Erfassung von Raubmilben, Thrips und anderen Kleinarthropoden im Weinbau. Schweiz. Z. Obs. Weinbau 1984, 120, 16–17. [Google Scholar]
- Hill, K.; Schlamp, H.A. Einsatz der Waschmethode zur Ermittlung des Raubmilbenbesatzes auf Rebblättern. Die Wein Wiss. 1984, 39, 255–262. [Google Scholar]
- Walter, D.E.; Lindquist, E.E.; Smith, I.M.; Cook, D.R.; Krantz, G.W.; Cook, D.R.; Krantz, G.W.; Cook, D.R.; Krantz, G.W. Order Trombidiformes. In A Manual of Acarology, 3rd ed.; Walter, D.E., Krantz, G.W., Eds.; Texas Tech University Press: Lubbock, TX, USA, 2009; pp. 233–420. ISBN 978-0-89672-620-8. [Google Scholar]
- Lindquist, E.E.; Krantz, G.W.; Walter, D.E. Order Mesostigmata. In A Manual of Acarology, 3rd ed.; Walter, D.W., Krantz, G.E., Eds.; Texas Tech University Press: Lubbock, TX, USA, 2009; pp. 124–232. ISBN 978-0-89672-620-8. [Google Scholar]
- Lindquist, E.E.; Amrine, J.W. 1.1.2 Systematics, diagnoses for major taxa, and keys to families and genera with species on plants of economic importance. In Eriophyoid Mites Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; World Crop Pests; Elsevier: Amsterdam, The Netherlands, 1996; Volume 6, ISBN 9780080531236. [Google Scholar]
- Walter, D.E.; Krantz, G.W. Collection, Rearing, and Preparing Specimens. In A Manual of Acarology, 3rd ed.; Walter, D.E., Krantz, G.W., Eds.; Texas Tech University Press: Lubbock, TX, USA, 2009; pp. 83–96. ISBN 978-0-89672-620-8. [Google Scholar]
- Tixier, M.S.; Baldassar, A.; Duso, C.; Kreiter, S. Phytoseiidae in European grape (Vitis vinifera L.): Bio-ecological aspects and keys to species (Acari: Mesostigmata). Zootaxa 2013, 3721, 101–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addison, J.A.; Hardman, J.M.; Walde, S.J. Pollen availability for predaceous mites on apple: Spatial and temporal heterogeneity. Exp. Appl. Acarol. 2000, 24, 1–18. [Google Scholar] [CrossRef]
- Jones, G.D. Pollen analyses for pollination research, unacetolyzed pollen. J. Pollinat. Ecol. 2012, 9, 1–12. [Google Scholar] [CrossRef]
- Beug, H.-J. Leitfaden der Pollenbestimmung für Mitteleuropa und angrenzende Gebiete; Verlag Friedrich Pfeil: München, Germany, 2015; ISBN 9783899370430. [Google Scholar]
- Rossi, V.; Caffi, T.; Legler, S.E. Dynamics of ascospore maturation and discharge in Erysiphe necator, the causal agent of grape powdery mildew. Phytopathology 2010, 100, 1321–1329. [Google Scholar] [CrossRef] [PubMed]
- Londo, G. The decimal scale for releves of permanent quadrats. Vegetatio 1976, 33, 61–64. [Google Scholar] [CrossRef]
- European Environment Agency (EEA) EUNIS Habitat Type. Available online: https://eunis.eea.europa.eu/habitats.jsp (accessed on 23 October 2019).
- Jackson, H.B.; Fahrig, L. Are ecologists conducting research at the optimal scale? Glob. Ecol. Biogeogr. 2015, 24, 52–63. [Google Scholar] [CrossRef]
- Redlich, S.; Martin, E.A.; Steffan-Dewenter, I. Landscape-level crop diversity benefits biological pest control. J. Appl. Ecol. 2018, 55, 2419–2428. [Google Scholar] [CrossRef]
- Hoy, M.A.; Groot, J.J.R.; van de Baan, H.E. Influence of aerial dispersal on persistence and spread of pesticide-resistant Metaseiulus occidentalis in California almond orchards. Entomol. Exp. Appl. 1985, 37, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Jung, C.; Croft, B.A. Aerial dispersal of phytoseiid mites (Acari: Phytoseiidae): Estimating falling speed and dispersal distance of adult females. Oikos 2001, 94, 182–190. [Google Scholar] [CrossRef]
- Environmental System Research Institute ArcGIS Desktop. Available online: http://desktop.arcgis.com/de (accessed on 23 October 2019).
- RStudio Team RStudio. Integrated Development for R; RStudio, Inc.: Boston, MA, USA. Available online: http://www.rstudio.com (accessed on 20 April 2020).
- Hesselbarth, M.H.K.; Sciaini, M.; With, K.A.; Wiegand, K.; Nowosad, J. Landscapemetrics: An open-source R tool to calculate landscape metrics. Ecography 2019, 42, 1648–1657. [Google Scholar] [CrossRef] [Green Version]
- OEPP/EPPO. Environmental risk assessment scheme for plant protection products, Chapter 10: Honeybees. Bull. OEPP/EPPO Bull. 2010, 40, 323–331. [Google Scholar]
- Lewis, K.A.; Tzilivakis, J.; Warner, D.J.; Green, A. An international database for pesticide risk assessments and management. Hum. Ecol. Risk Assess. An Int. J. 2016, 22, 1050–1064. [Google Scholar] [CrossRef] [Green Version]
- Austrian Federal Office for Food Safety (BAES). Pflanzenschutzmittel-Register—Verzeichnis der in Österreich Zugelassenen/Genehmigten Pflanzenschutzmittel. Available online: https://psmregister.baes.gv.at/psmregister/faces/main?_afrLoop=3293465951961555&_afrWindowMode=0&_adf.ctrl-state=18vmkamvgh_4 (accessed on 20 April 2020).
- National Assessment—Federal Republic of Germany Draft Registration Report—Central Zone, Product Code: ACH-07547-F-0-SPKumar: 007547-00/10. Available online: https://www.bvl.bund.de/SharedDocs/Downloads/04_Pflanzenschutzmittel/01_zulassungsberichte/007547-00-10.pdf?__blob=publicationFile&v=3 (accessed on 9 April 2020).
- Hassan, S.A. Standard methods to test the side-effects of pesticides on natural enemies of insects and mites developed by the lOBC/WPRS Working Group ‘Pesticides and Beneficial Organisms. Bull. OEPP/EPPO Bull. 1985, 15, 214–255. [Google Scholar] [CrossRef]
- Abteilung für Pflanzenschutzmittel und Anwendungstechnik Datenanforderungen und Entscheidungskriterien der Europäischen Union und der Bundesrepublik Deutschland im Zulassungsverfahren für Pflanzenschutzmittel. Mitt. Biol. Bundesanst. Land Forstwirtsch. 1998, 354, 1–156.
- Bundesamt für Verbraucherschutz und Lebensmittelsicherheit Verzeichnis zugelassener Pflanzenschutzmittel. Available online: https://apps2.bvl.bund.de/psm/jsp/index.jsp (accessed on 4 August 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria. Available online: https://www.r-project.org (accessed on 20 April 2020).
- Laliberté, E.; Legendre, P.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology (R Package Version 1.0-12). Available online: https://cran.r-project.org/web/packages/FD/index.html (accessed on 20 April 2020).
- Sarkar, D. Lattice: Multivariate Data Visualization with R.; Springer: New York, NY, USA, 2008; ISBN 978-0-387-75968-5. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; ISBN 0-387-98454-2. [Google Scholar]
- Fox, J. Effect Displays in R for Generalised Linear Models. J. Stat. Softw. 2003, 8, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix (Version 0.84). Available online: https://github.com/taiyun/corrplot (accessed on 20 April 2020).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Bartoń, K. MuMIn: Multi-Model Inference. R Package Version 1.43.17. Available online: https://cran.r-project.org/package=MuMIn (accessed on 20 April 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- O’Hara, R.B.; Kotze, D.J. Do not log-transform count data. Methods Ecol. Evol. 2010, 1, 118–122. [Google Scholar] [CrossRef] [Green Version]
- Curran-Everett, D. Explorations in statistics: The log transformation. Adv. Physiol. Educ. 2018, 42, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Wang, H.; Lu, N.; Chen, T.; He, H.; Lu, Y.; Tu, X.M. Log-transformation and its implications for data analysis. Shanghai Arch. Psychiatry 2014, 26, 105–109. [Google Scholar] [CrossRef]
- Bolker, B.M.; Gardner, B.; Maunder, M.; Berg, C.W.; Brooks, M.; Comita, L.; Crone, E.; Cubaynes, S.; Davies, T.; de Valpine, P.; et al. Strategies for fitting nonlinear ecological models in R, AD Model Builder, and BUGS. Methods Ecol. Evol. 2013, 4, 501–512. [Google Scholar] [CrossRef] [Green Version]
- Montgomery, D.C.; Peck, E.A.; Vining, G.G. Introduction to Linear Regression Analysis, 5th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; ISBN 978-0-470-54281-1. [Google Scholar]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of akaike information criterion and bayesian approaches over likelihood ratio tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef]
- Mazerolle, M.J. AICcmodavg: Model Selection and Multimodel Inference Based on (Q)AIC(c). R P Package Version 2.3-0. Available online: https://cran.r-project.org/web/packages/AICcmodavg/index.html (accessed on 20 April 2020).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference a Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2002; ISBN 0-387-95364-7. [Google Scholar]
- Motulsky, H.; Christopoulos, A. Fitting Models to Biological Data using Linear and Nonlinear Regression: A Practical Guide to Curve Fitting; Oxford University Press: Oxford, UK, 2004; ISBN 1425919448. [Google Scholar]
- Weaver, K.F.; Morales, V.; Dunn, S.L.; Godde, K.; Weaver, P.F. Pearson’s and Spearman’s Correlation. In An Introduction to Statistical Analysis in Research; Weaver, K.F., Morales, V., Dunn, S.L., Godde, K., Weaver, P.F., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 435–471. ISBN 9781119454205. [Google Scholar]
- Schober, P.; Schwarte, L.A. Correlation coefficients: Appropriate use and interpretation. Anesth. Analg. 2018, 126, 1763–1768. [Google Scholar] [CrossRef]
- Revelle, W. psych: Procedures for Psychological, Psychometric, and Personality Research, R package version 2.0.9. Available online: https://cran.r-project.org/package=psych (accessed on 20 April 2020).
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6. Available online: https://cran.r-project.org/package=vegan (accessed on 20 April 2020).
- Paliy, O.; Shankar, V. Application of multivariate statistical techniques in microbial ecology. Mol. Ecol. 2016, 25, 1032–1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Oksanen, J. Multivariate Analysis of Ecological Communities in R: Vegan Tutorial. Available online: https://www.mooreecology.com/uploads/2/4/2/1/24213970/vegantutor.pdf (accessed on 20 April 2020).
- Hoffmann, C.; Köckerling, J.; Biancu, S.; Gramm, T.; Michl, G.; Entling, M.H. Can flowering greencover crops promote biological control in German vineyards? Insects 2017, 8, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duso, C.; Vettorazzo, E. Mite population dynamics on different grape varieties with or without phytoseiids released (Acari: Phytoseiidae). Exp. Appl. Acarol. 1999, 23, 741–763. [Google Scholar] [CrossRef]
- Duso, C.; Pozzebon, A.; Capuzzo, C.; Malagnini, V.; Otto, S.; Borgo, M. Grape downy mildew spread and mite seasonal abundance in vineyards: Effects on Tydeus caudatus and its predators. Biol. Control. 2005, 32, 143–154. [Google Scholar] [CrossRef]
- Linder, C.; Viret, O.; Spring, J.-L. Viticulture integree et bio-organique: Synthese de sept ans d’observation. Rev. Suisse Vitic. Arboric. Hortic. 2006, 38, 235–244. [Google Scholar]
- Reiff, J.M.; Ehringer, M.; Hoffmann, C.; Entling, M.H. Fungicide reduction favors the control of phytophagous mites under both organic and conventional viticulture. Agric. Ecosyst. Environ. 2021, 305. [Google Scholar] [CrossRef]
- Hluchý, M.; Pospíšil, Z.; Zacharda, M. Phytophagous and predatory mites (Acari: Tetranychidae, Eriophyidae, Phytoseiidae, Stigmaeidae) in South Moravian vineyards, Czechoslovakia, treated with various types of chemicals. Exp. Appl. Acarol. 1991, 13, 41–52. [Google Scholar] [CrossRef]
- James, D.G.; Price, T.S.; Wright, L.C.; Perez, J. Abundance and phenology of mites, leafhoppers, and thrips on pesticide-treated and untreated wine grapes in southcentral Washington. J. Agric. Urban Entomol. 2002, 19, 45–54. [Google Scholar]
- Pennington, T.; Kraus, C.; Alakina, E.; Entling, M.H.; Hoffmann, C. Minimal pruning and reduced plant protection promote predatory mites in Grapevine. Insects 2017, 8, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, M.J. Impact of sulfur on density of Tetranychus pacificus (Acari: Tetranychidae) and Galendromus occidentalis (Acari: Phytoseiidae) in a central California vineyard. Exp. Appl. Acarol. 2007, 42, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Pozzebon, A.; Duso, C.; Pavanetto, E. Side effects of some fungicides on phytoseiid mites (Acari, Phytoseiidae) in north-Italian vineyards. J. Pest Sci. 2002, 75, 132–136. [Google Scholar] [CrossRef]
- Kemmitt, G.; Valverde-Garcia, P.; Hufnagl, A.; Bacci, L.; Zotz, A. The impact of three commonly used fungicides on Typhlodromus pyri (Acari: Phytoseiidae) in European vineyards. J. Econ. Entomol. 2015, 108, 611–620. [Google Scholar] [CrossRef]
- English-Loeb, G.; Norton, A.P.; Gadoury, D.M.; Seem, R.C.; Wilcox, W.F. Control of powdery mildew in wild and cultivated grapes by a tydeid mite. Biol. Control. 1999, 14, 97–103. [Google Scholar] [CrossRef]
- Maixner, M. Studies on the insecticide resistance of the predatory mite Typhlodromus pyri on grapevines of the wine-region Mosel-Saar-Ruwer. Mitt. Biol. Bundesanst. Land Forstwirtsch. Berlin Dahl. 1990, 257, 118. [Google Scholar]
- Blümel, S.; Bakker, F.M.; Baier, B.; Brown, K.; Candolfi, M.P.; Goßmann, A.; Grimm, C.; Jäckel, B.; Nienstedt, K.; Schirra, K.J.; et al. Laboratory residual contact test with the predatory mite Typhlodromus pyri Scheuten (Acari: Phytoseiidae) for regulatory testing of plant protection products. In Guidelines to Evaluate Side-Effects of Plant Protection Products to Non-Target Arthropods; Candolfi, M.P., Blümel, S., Forster, R., Bakker, F.M., Grimm, C., Hassan, S.A., Heimbach, U., Mead, M., Reber, B.F., Schmuck, R., Vogt, H., Eds.; IOBC/WPRS: Gent, Belgium, 2000; pp. 121–143. ISBN 92-9067-129-7. [Google Scholar]
- Daane, K.M.; Hogg, B.N.; Wilson, H.; Yokota, G.Y. Native grass ground covers provide multiple ecosystem services in Californian vineyards. J. Appl. Ecol. 2018, 55, 2473–2483. [Google Scholar] [CrossRef] [Green Version]
- Lorenzon, M.; Pozzebon, A.; Duso, C. Effects of potential food sources on biological and demographic parameters of the predatory mites Kampimodromus aberrans, Typhlodromus pyri and Amblyseius andersoni. Exp. Appl. Acarol. 2012, 58, 259–278. [Google Scholar] [CrossRef]
- Mailloux, J.; Le Bellec, F.; Kreiter, S.; Tixier, M.S.; Dubois, P. Influence of ground cover management on diversity and density of phytoseiid mites (Acari: Phytoseiidae) in Guadeloupean citrus orchards. Exp. Appl. Acarol. 2010, 52, 275–290. [Google Scholar] [CrossRef] [PubMed]
- De Villiers, M.; Pringle, K.L. The presence of Tetranychus urticae (Acari: Tetranychidae) and its predators on plants in the ground cover in commercially treated vineyards. Exp. Appl. Acarol. 2011, 53, 121–137. [Google Scholar] [CrossRef]
- Wari, D.; Funayama, K.; Kishimoto, H.; Toyama, M.; Sonoda, S. Molecular verification of dispersal of phytoseiid mites from groundcover plants to tree leaves in Japanese peach orchards. Biol. Control. 2015, 80, 143–155. [Google Scholar] [CrossRef]
- Sahraoui, H.; Kreiter, S.; Lebdi-Grissa, K.; Tixier, M.S. Sustainable weed management and predatory mite (acari: Phytoseiidae) dynamics in Tunisian citrus orchards. Acarologia 2016, 56, 517–532. [Google Scholar] [CrossRef] [Green Version]
- Duffner, K. Untersuchungen zur Biologie, Morphologie und Bekämpfung der Kräuselmilbe Calepitrimerus vitis Nalepa 1905 (Acari, Eriophyoidea). Ph.D. Thesis, University of Freiburg, Freiburg im Breisgau, Germany, 1999. [Google Scholar]
- Duso, C.; De Lillo, E. 3.2.5 Grape. In Eriophyoid Mites Their Biology, Natural Enemies and Control; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; World Crop Pests; Elsevier: Amsterdam, The Netherlands, 1996; Volume 6, pp. 571–582. ISBN 9780080531236. [Google Scholar]
- Hluchý, M. Zur biologischen Bekämpfung der Kräuselmilbe Calepitrimerus vitis Nalepa (Acari, Eriophyidae) auf der Weinrebe durch die Raubmilbe Typhlodromus pyri Scheuten (Acari, Phytoseiidae). J. Appl. Entomol. 1993, 116, 449–458. [Google Scholar] [CrossRef]
- Rusch, A.; Delbac, L.; Thiéry, D. Grape moth density in Bordeaux vineyards depends on local habitat management despite effects of landscape heterogeneity on their biological control. J. Appl. Ecol. 2017, 54, 1794–1803. [Google Scholar] [CrossRef] [Green Version]
- Wilson, H.; Miles, A.F.; Daane, K.M.; Altieri, M.A. Landscape diversity and crop vigor influence biological control of the western grape leafhopper (E. elegantula Osborn) in vineyards. PLoS ONE 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Judt, C.; Guzmán, G.; Gómez, J.A.; Cabezas, J.M.; Entrenas, J.A.; Winter, S.; Zaller, J.G.; Paredes, D. Diverging effects of landscape factors and inter-row management on the abundance of beneficial and herbivorous arthropods in andalusian vineyards (Spain). Insects 2019, 10, 320. [Google Scholar] [CrossRef] [Green Version]
- Gavinelli, F.; Fusaro, S.; Ivan, D.; Ragusa, S.; Paoletti, M.G. Unpredicted ecological and ecosystem services of biodiversity. Spontaneous vegetation, hedgerows, and maple trees as useful landscape components to increase predatory mite population in agroecosystems. Appl. Soil Ecol. 2020, 154. [Google Scholar] [CrossRef]
- Tixier, M.S.; Kreiter, S.; Auger, P.; Weber, M. Colonization of Languedoc vineyards by phytoseiid mites (Acari: Phytoseiidae): Influence of wind and crop environment. Exp. Appl. Acarol. 1998, 22, 523–542. [Google Scholar] [CrossRef]
- Tixier, M.S.; Kreiter, S.; Cheval, B.; Guichou, S.; Auger, P.; Bonafos, R. Immigration of phytoseiid mites from surrounding uncultivated areas into a newly planted vineyard. Exp. Appl. Acarol. 2006, 39, 227–242. [Google Scholar] [CrossRef]
- Kreiter, S.; Sentenac, G.; Rusch, A.; Zriki, G.; Thiery, J.; Delbac, L.; Madejsky, M.; Guisset, M.; Tixier, M.S. Effects of the plot landscape and pollen concentration on phytoseiid mites (Acari: Phytoseiidae) in three French viticultural regions. In Proceedings of the AFPP—11e Conférence Internationale sur les Ravageurs et Auxiliaires en Agriculture, Montpellier, France, 25–26 October 2017; pp. 361–370. [Google Scholar]
- Fischer-Colbrie, P.; El-Borolossy, M. Untersuchungen zum Einfluß des Klimas, der Pflanzenart und der Wirtstiere auf das Vorkommen verschiedener Raubmilbenarten im österreichischen Obst- und Weinbau. Pflanzenschutzberichte 1990, 51, 101–124. [Google Scholar]
- Ahmad, S.; Pozzebon, A.; Duso, C. Predation on heterospecific larvae by adult females of Kampimodromus aberrans, Amblyseius andersoni, Typhlodromus pyri and Phytoseius finitimus (Acari: Phytoseiidae). Exp. Appl. Acarol. 2015, 67, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Kingsolver, J.G.; Diamond, S.E.; Buckley, L.B. Heat stress and the fitness consequences of climate change for terrestrial ectotherms. Funct. Ecol. 2013, 27, 1415–1423. [Google Scholar] [CrossRef]
- Landers, A.J. Spray technology for grapevines. In Compendium of Grape Diseases, Disorders, and Pests; Wilcox, W.F., Gubler, W.D., Uyemoto, J.K., Eds.; The American Phytopathological Society: St. Paul, MN, USA, 2015; pp. 185–192. ISBN 978-0-89054-479-2. [Google Scholar]
- Faegri, K.; Iversen, J.; Waterbolk, H.T. Textbook of Pollen Analysis, 2nd ed.; Munksgaard: Copenhagen, Denmark, 1964. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Management-Related Numeric Explanatory Variables | Organic | Integrated |
|---|---|---|
| aAPTLc (area-related acute pesticide contact toxicity loading) | 19.91 ± 6.69 | 10.63 ± 14.59 |
| Categorical toxicity rating for Typhlodromus pyri | 14.65 ± 3.57 | 13.19 ± 5.13 |
| Number of pesticide applications (total) | 9.69 ± 1.7 | 6.31 ± 2.24 |
| Insecticide applications | 0 ± 0 | 0.25 ± 0.45 |
| Acaricide applications | 0.25 ± 0.45 | 0.38 ± 0.5 |
| Synthetic fungicide applications | 0 ± 0 | 5.79 ± 2.58 |
| Sulfur applications | 9.19 ± 1.6 | 4.13 ± 1.84 |
| Copper applications | 8.88 ± 1.84 | 1.25 ± 0.83 |
| Potassium bicarbonate applications | 4.44 ± 2.76 | 0 ± 0 |
| Vegetation cover in spring (%) | 82.38 ± 18.94 | 80.84 ± 19.51 |
| Vegetation cover in summer (%) | 61.70 ± 22.26 | 40.36 ± 24.78 |
| Response Variable | Best Model | AICc | Δi | Adjusted R2 |
|---|---|---|---|---|
| Phytoseiid mite densities | null model | 37.22 | - | - |
| date + management + cover crop type + proportion of vineyards + proportion of total SNHs | −45.87 | 0.0 | 0.44 | |
| date + management + cover crop type + proportion of vineyards + proportion of total SNHs + pollen total | −43.74 | 2.13 | 0.44 | |
| Tydeoid mite densities | null model | 9.22 | - | - |
| date + cover crop type | −90.55 | 0.0 | 0.49 | |
| date + management + cover crop type | −89.71 | 0.84 | 0.49 | |
| date + management + cover crop type + proportion of vineyards | −88.75 | 1.80 | 0.49 | |
| date + management + cover crop type + proportion of total SNHs | −87.97 | 2.58 | 0.49 | |
| Eriophyoid mite densities | null model | −158.21 | - | - |
| date + proportion of vineyards + vegetation cover summer + densities of phytoseiid mites | −221.32 | 0.0 | 0.36 | |
| date + proportion of vineyards + vegetation cover summer | −220.29 | 1.03 | 0.35 | |
| date + proportion of vineyards + vegetation cover summer + vegetation cover spring | −219.35 | 1.97 | 0.35 | |
| date + vegetation cover summer | −218.20 | 3.12 | 0.34 | |
| Pollen densities | null model | 150.85 | - | - |
| date + management + minimum distance to the next woody SNHs | −10.43 | 0.0 | 0.65 | |
| date + management | −10.04 | 0.39 | 0.65 | |
| date + minimum distance to the next woody SNHs | −9.72 | 0.71 | 0.65 | |
| date | −9.53 | 0.90 | 0.64 | |
| date + cover crop type | −8.94 | 1.49 | 0.65 | |
| date + management + cover crop type + minimum distance to the next woody SNHs | −8.64 | 1.79 | 0.65 | |
| date + management + cover crop type | −8.60 | 1.83 | 0.65 | |
| date + management + proportion of vineyards + minimum distance to the next woody SNHs | −8.59 | 1.84 | 0.65 | |
| date + management + proportion of total SNHs + minimum distance to the next woody SNHs | −8.34 | 2.09 | 0.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Möth, S.; Walzer, A.; Redl, M.; Petrović, B.; Hoffmann, C.; Winter, S. Unexpected Effects of Local Management and Landscape Composition on Predatory Mites and Their Food Resources in Vineyards. Insects 2021, 12, 180. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020180
Möth S, Walzer A, Redl M, Petrović B, Hoffmann C, Winter S. Unexpected Effects of Local Management and Landscape Composition on Predatory Mites and Their Food Resources in Vineyards. Insects. 2021; 12(2):180. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020180
Chicago/Turabian StyleMöth, Stefan, Andreas Walzer, Markus Redl, Božana Petrović, Christoph Hoffmann, and Silvia Winter. 2021. "Unexpected Effects of Local Management and Landscape Composition on Predatory Mites and Their Food Resources in Vineyards" Insects 12, no. 2: 180. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12020180