
Monitoring Exotic Beetles with Inexpensive Attractants: A Case Study




1
Department of Agronomy, Food, Natural Resources, Animals and the Environment (DAFNAE), University of Padova, Viale dell’Universita 16, Legnaro, 35020 Padova, Italy
2
World Biodiversity Association Onlus c/o NAT LAB Forte Inglese, Portoferraio, 57037 Livorno, Italy
3
Department of Earth and Environmental Sciences, University of Milano-Bicocca, Piazza della Scienza 1, 20126 Milano, Italy
*
Author to whom correspondence should be addressed.
Insects 2021, 12(5), 462; https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050462
Submission received: 1 May 2021
/
Revised: 12 May 2021
/
Accepted: 14 May 2021
/
Published: 17 May 2021
(This article belongs to the Collection Surveillance Tools and Monitoring Strategies for Native and Exotic Forest Insects)
Abstract
:Simple Summary
Detecting and monitoring exotic and invasive beetles is a complex activity, and multiple species still manage to evade controls. Citizen science can be an important adjunct in alien species monitoring programs, but to have a greater chance of success, it must employ traps and attractants that are easy to gather and use. Bottle traps baited with food products are successfully used during long term faunistic surveys, and the same methodology can be adapted to alien species detection and monitoring. In this article, we tested the use of bottles baited with apple cider vinegar, red wine, and 80% ethyl alcohol in capturing exotic and invasive beetles in the surroundings of Malpensa Airport (Italy). The traps proved effective, and in the traps with vinegar as an attractant, they captured four out of five invasive Nitidulidae, as well as the only invasive Scarabaeidae present in the area. Popillia japonica’s response to apple cider vinegar is documented for the first time and suggests the use of this attractant in monitoring surveys for this species, especially if supported by citizen science programs. The substantial reduction in the activity time of the traps seems to have considerably reduced collateral catches of native fauna.
Abstract
Detecting and monitoring exotic and invasive Coleoptera is a complex activity to implement, and citizen science projects can provide significant contributions to such plans. Bottle traps are successfully used in wildlife surveys and can also be adapted for monitoring alien species; however, a sustainable, large scale trapping plan must take into account the collateral catches of native species and thus minimize its impact on local fauna. In the present paper, we tested the use of bottles baited with standard food products that can be purchased in every supermarket and immediately used (apple cider vinegar, red wine, and 80% ethyl alcohol) in capturing exotic and invasive beetles in the area surrounding Malpensa Airport (Italy). In particular, we reduced the exposition type of the traps in each sampling round to three days in order to minimize native species collecting. We found a significant effect of the environmental covariates (trap placement, temperature, humidity, and forest type) in affecting the efficiency in catching target beetles. Nearly all invasive Nitidulidae and Scarabaeidae known to be present in the area were captured in the traps, with apple cider vinegar usually being the most effective attractant, especially for the invasive Popillia japonica.
1. Introduction
Among European countries, Italy has the most exotic taxa (species that are not native to a specific ecosystem), several of which are invasive (organisms that cause ecological or economic harm in an ecosystem where they are not native) [1]; among these, Coleoptera alone account for more than 300 intercepted or established species [2,3,4,5,6,7,8,9,10,11,12,13]. Insect introductions through human-mediated dispersal is ever increasing because of globalization, and represents a serious threat to biodiversity, local economies and animal and plant health [14]. The ever-increasing number and types of goods transported and the speed at which commercial traffic occurs, associated with the opening of new trade routes, are increasingly resulting in faunal exchanges among and within biogeographic realms [15]. Italy’s predisposition to beetle (Coleoptera) introductions is likely associated with its geographic position in the center of the Mediterranean, being at the crossroads of much commerce to and from Europe [1,16].
Attention and awareness towards alien species have increased in recent decades, even though strategies to reduce future invasions have not yet been implemented in an effective and widespread manner on a global scale [17].
If we consider the number of exotic species recorded in the last few years, and especially those collected fortuitously, it is clear that current monitoring strategies are ineffective at detecting several beetle families. It is for this reason that great effort has been invested in improving monitoring strategies, survey methods, and traps [18,19], with a focus on the main entry points such as seaports and airports (i.e., [20,21]). However, much of these recent developments have targeted primarily families of forest insects, mostly wood borers, such as Curculionidae: Scolytinae and Platypodinae, Cerambycidae, and Buprestidae (e.g., [22,23,24]). Furthermore, biosecurity surveillance suffers from two major issues, namely: (1) effective monitoring strategies, especially those that target multiple taxa, which are generally expensive because of the cost of traps and pheromones, and (2) difficulty in applying targeted monitoring strategies simultaneously and on a national scale.
For this reason, the scientific community is increasingly availing itself of the support of citizen science as a means of strengthening its surveillance capacity (e.g., [25,26]). However, for a monitoring plan supported by citizen science to be effective, it is necessary that it is easily reproducible, low-cost, and does not involve a heavy workload for volunteers.
Bottle traps is a methodology commonly and successfully used in faunistic surveys [27,28]. Bottle traps are inexpensive, easy to make and transport, and have been recently suggested and applied in bark and ambrosia beetle (Scolytinae and Platypodinae) monitoring through citizen participation [29,30,31].
Based on these concepts, we decided to conduct a trial intended to evaluate if bottle traps could be used to monitor certain exotic beetles that are not commonly targeted with standard traps (e.g., Lindgren funnels traps and cross-vane panel traps) and pheromones, especially using food products as lures that can be purchased directly at the supermarket. In the scientific literature, there are many attractive mixtures made from fermented goods (such as honey, bananas, and beer; e.g., [32,33]). However, as we could not expect all volunteers to produce such attractive mixtures, we opted for three affordable products that can be used directly as they are when purchased, namely: food-grade ethyl alcohol (which will be referred to as alcohol), red wine, and apple cider vinegar.
However, as the objective of this trial was to target exotic species, the exposure times of our traps were reduced to only three days each during each survey round. This decision started from the assumption that invasive species are characterized by biological and ecological traits that make them more prone to rapidly respond to generic olfactory stimuli (e.g., food/reproduction sources) compared with native species. Given the collateral catch that these monitoring activities can have on native species, we expected that reducing the traps’ exposure time would maintain the capture of exotics, while minimizing the impact on native species.
In the present study, we evaluated (1) the effectiveness of bottle traps baited with non-fermented goods and short exposure times in catching exotic beetles, (2) the beetle families and species collected in terms of diversity and abundance, and (3) how the different baits varied in attractiveness to target beetle species. Furthermore, we tried to assess the effect of temperature, humidity, environmental surroundings, and trap placement on trapping efficiency.
2. Materials and Methods
2.1. Study Area
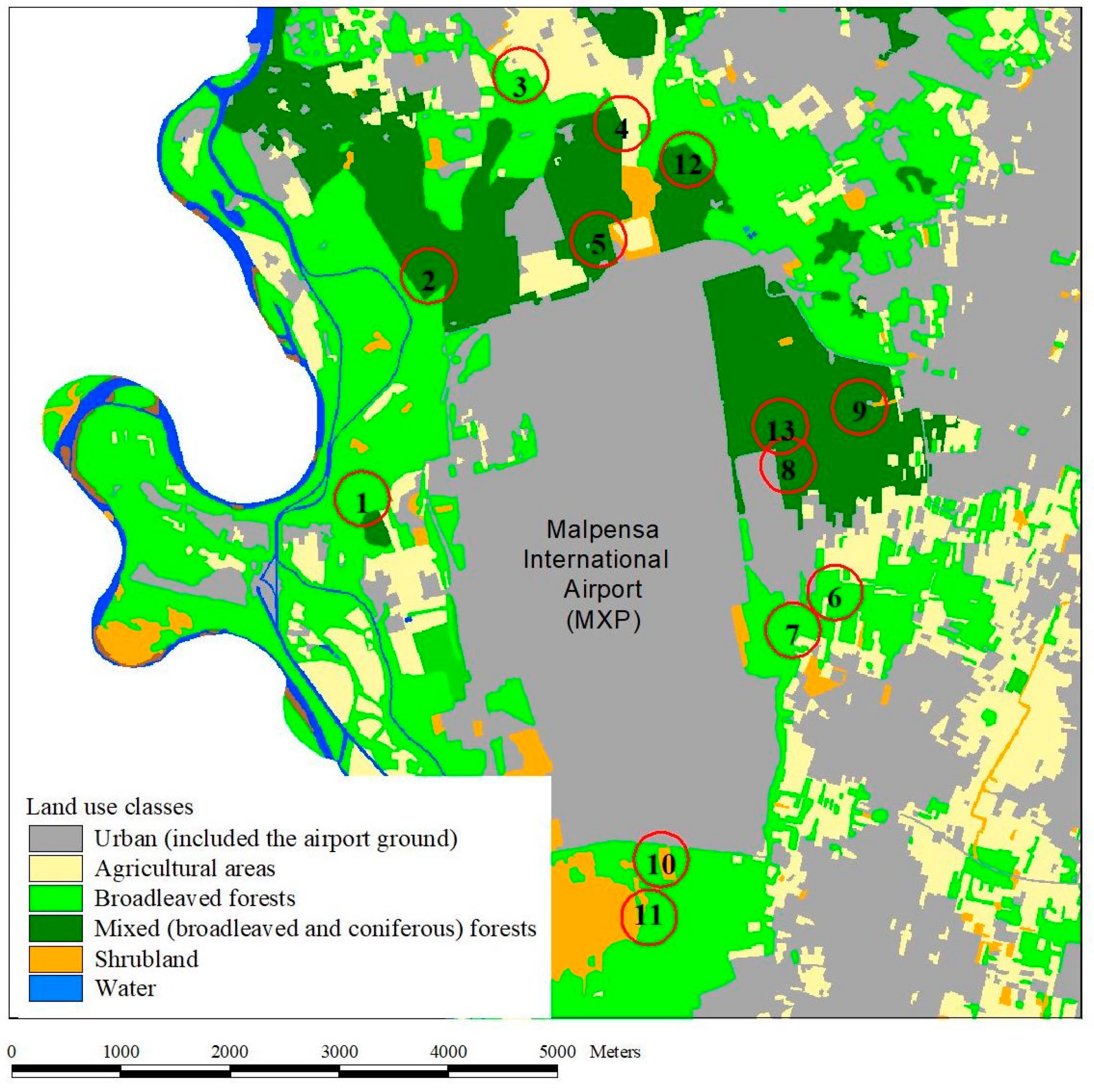
The study area is centered on the Malpensa International Airport (MXP), the largest international airport in northern Italy, located in the Ferno Municipality (Varese Province), within the Lombardy Regional Park of Ticino Valley, about 50 km northwest of Milan. The airport grounds are largely surrounded by deciduous and mixed (broad-leaved and coniferous) forests and urban areas, and, to a lesser extent, agricultural areas and shrubland (Figure 1). Among the trees, the dominant taxa are oaks (Quercus spp.), maples (Acer spp.), and Scots pine (Pinus sylvestris). Most of the woodlands are managed as coppice, and the presence of exotic trees is not negligible, with the Locust tree (Robinia pseudoacacia) among the most widespread species.
2.2. Experimental Design, Traps, and Baits
The traps were placed in 13 sampling sites identified to account for habitat covariates, i.e., the forest type (broadleaved vs. mixed) and forest edge vs. forest interior condition (interior: condition with forest fractional cover >90% evaluated in a buffer of 250 m around trapping site), based on DUSAF digital cartography [34] (Table 1). In each sampling site, three traps were used, each baited with either 80% food alcohol, commercial red wine (11.5% alcohol), and apple cider vinegar (4% acetic acid), for a total of 39 traps. No preservatives were added. In each sampling site, the three traps were placed along the perimeter of an ideal circle with about a 15 m radius (thus having a distance of at least 30 m among traps) to avoid their mutual influence on capture events; all traps were placed 2.5 m from the ground. Each trap was built using a 500 mL volume PET (polyethylene terephthalate). A 5 × 8.5 cm window was created on half of each bottle by removing a sizeable lateral portion, leaving the rest of the bottle intact (to guarantee the structural integrity and support of the modest weight of the bait). Each set of baited traps was left in action for 72 h, and then removed and emptied each week between 31 July and 29 September 2020, for a total of seven trapping periods. For each trapping session, we recorded the absolute minimum and maximum temperature during the three days of activity of the traps, the three-days mean of the mean daily temperatures, and the three-days mean of the humidity degree. As the three temperature values were correlated within each trapping session, we used the absolute maximum temperature (Table 2). The meteorological data were obtained from the airport meteorological station.
All of the non-Coleoptera (i.e., Diptera, Lepidoptera, Hymenoptera, and Hemiptera) collected during the survey were not considered in the analyses and were discarded.
2.3. Analyses
We evaluated the overall effectiveness of each type of bait (red wine, vinegar, or alcohol), considering the overall number of individuals of exotic species pooled together caught by each kind of bait in each site and session, accounting for both habitat and meteorological covariates. The same model was applied to the overall number of individuals for the native species subset. The analysis was performed using a negative binomial regression for modelling count data using MASS package [35] in R version 4.0.3 [36]. The assessment of the data distribution for the dependent variable was performed using the fitdistrplus package [37]. Plots of the conditional effects (habitat variables) and main effect (meteorological variables) of covariates were made using the sjPlot package [38], whose functionality depends on the ggplot2 package [39]. The same analysis was then performed to evaluate the specific overall effectiveness of each bait, considering the number of individuals pertaining to the most commonly trapped species.
3. Results
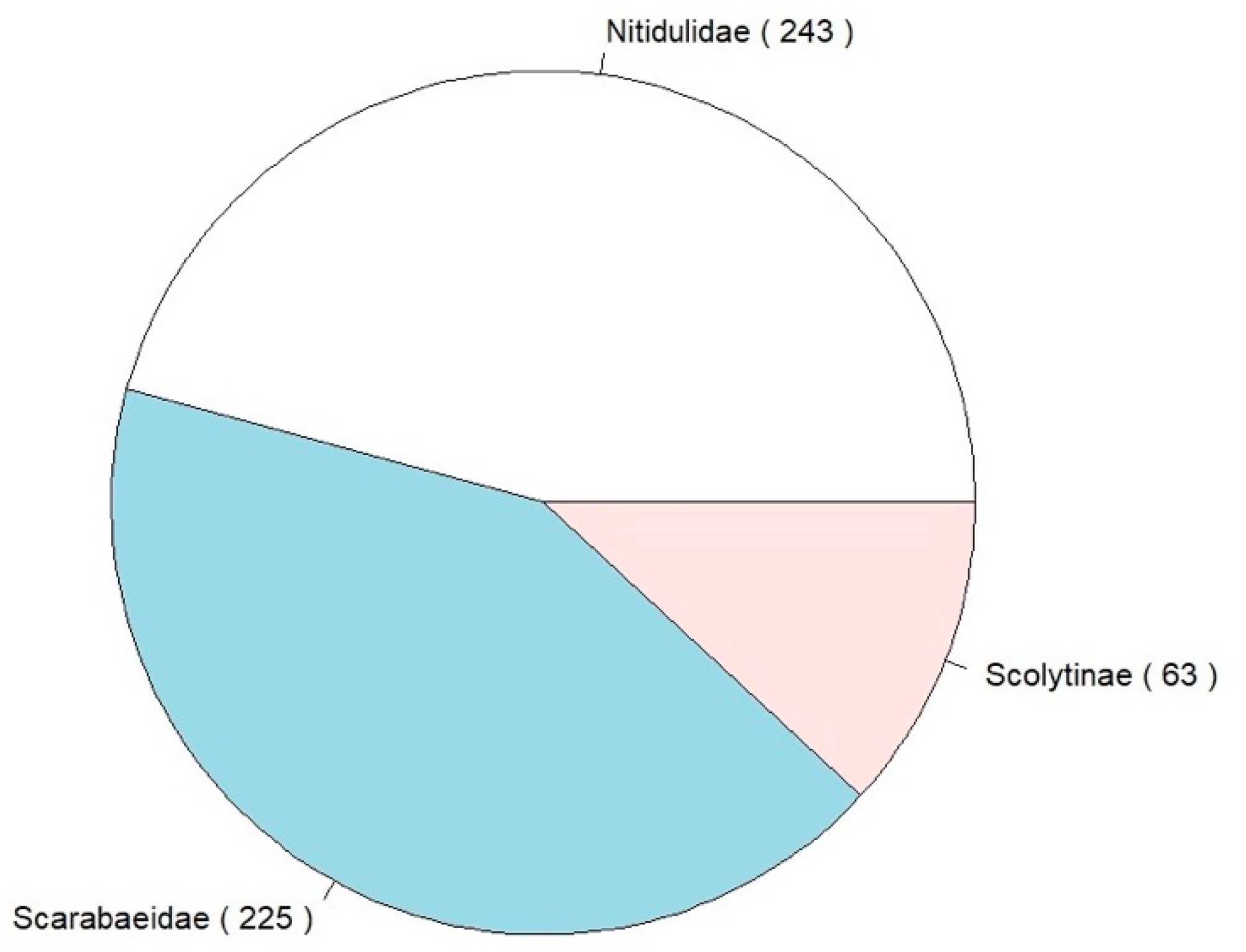
During the two-months survey period (i.e., seven three-day sessions), in the 13 sampling sites, we caught a total of 531 individuals (437 pertaining to exotic and 94 to native species). The 14 species of beetles collected belonged to Scolytinae, Nitidulidae, and Scarabaeidae (Figure 2 and Table 3). Among the species collected, five are considered invasive in Europe (Carpophilus lugubris, Epuraea luteola, Epuraea ocularis, Glischrochilus quadrisignatus, and Popilia japonica; Figure 3).
The most common species caught were P. japonica (Scarabaeidae: Rutelinae), with 212 individuals (40% of the overall individual caught and 49% of the individuals of exotic species), and E. ocularis (Nitidulidae), with 159 individuals (30% of the overall individual caught and 36% of the exotic species). The third most common species was the native X. saxesenii (Scolytinae), with 58 individuals (11% of the overall individual caught and 62% of native species).
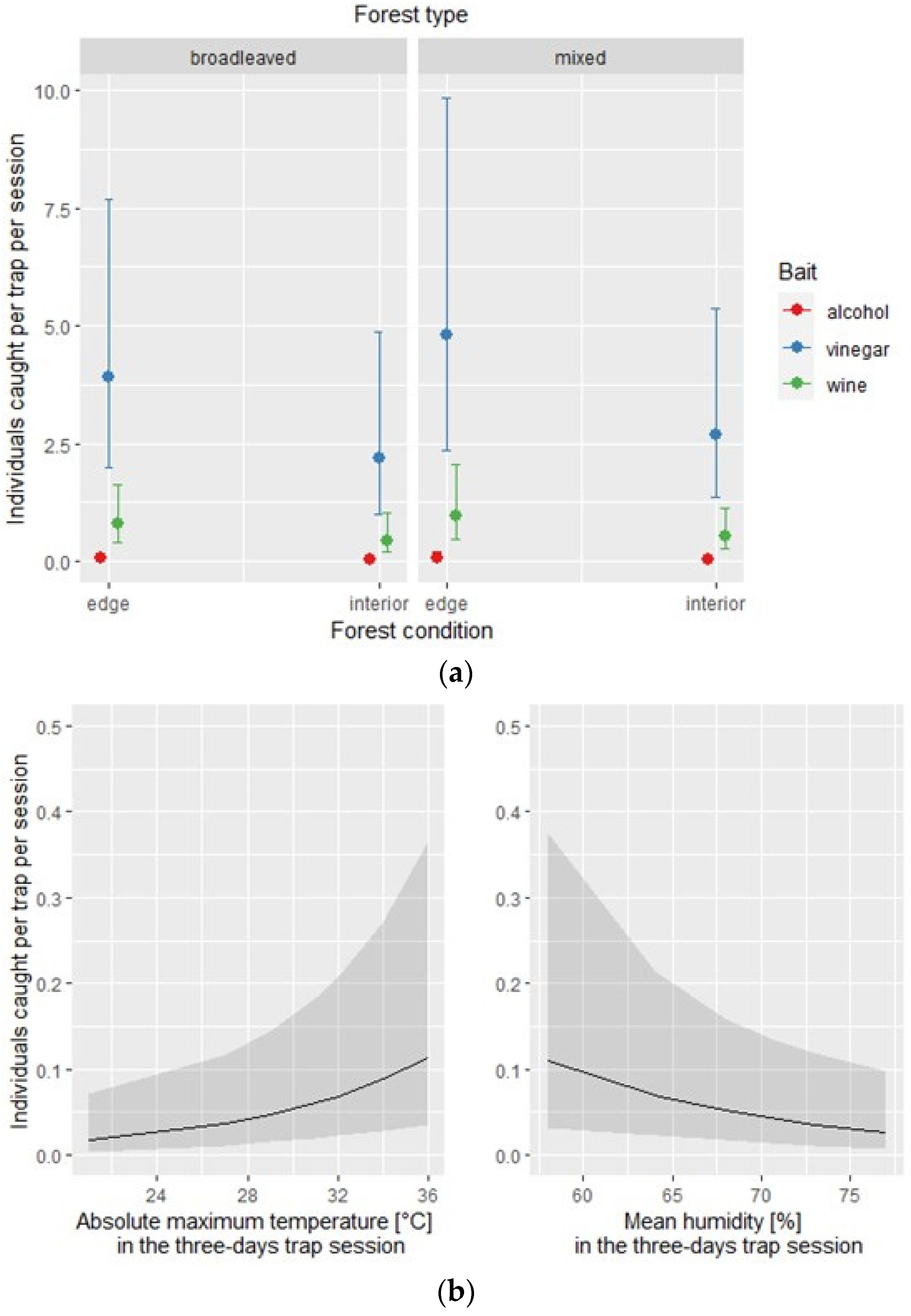
The negative binomial regression models used to assess each bait’s effectiveness in attracting the overall number of exotic and native species explained 40% and 13% of the sample deviance, respectively. For the exotic species subset, the model identified the type of bait, the absolute maximum temperature, and the mean humidity as statistically significant for affecting the overall number of individuals caught in each of the three traps per site and session (Table 4). The conditional effect of each covariate is depicted in Figure 3.
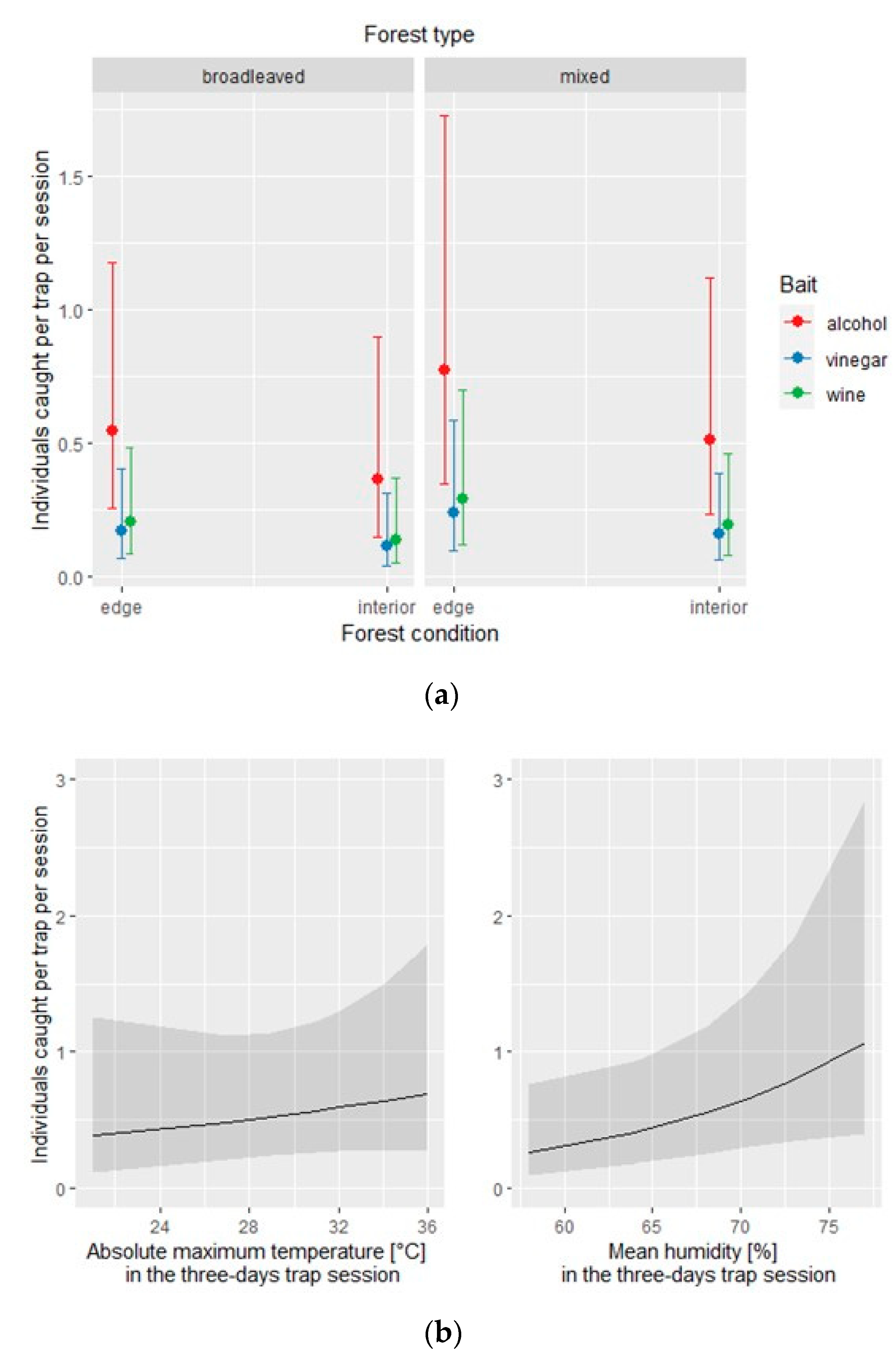
The same model applied to the native species subset identified the type of bait and the mean humidity as statistically significant (Table 5). The conditional effect of each covariate is depicted in Figure 4.
For the three most common trapped species (two exotic and one native), the models for the P. japonica, E. oculari, and X. saxesenii explained 72%, 40%, and 32% of the sample deviance, respectively. We must highlight that all of the individuals, but one of X. saxesenii, were caught in alcohol. Thus, to find the covariate that affects the number of individuals caught, we used only those collected by the traps triggered by alcohol.
For P. japonica, the type of bait’s effectiveness differed significantly, and both the habitat and meteorological covariates significantly affected the number of individuals caught. Specifically, both vinegar and wine positively enhanced the trap efficiency in collecting P. japonica, with vinegar being more effective than wine. Furthermore, the number of collected specimens increased with increasing temperatures and captures, and were substantially higher when traps were placed at the edges of broadleaved forests (Table 6). The conditional effect of each covariate is depicted in Figure 5.
For E. ocularis, the type of bait also significantly influenced the effectiveness of trapping, but only the habitat covariates showed a substantial effect on the number of individuals caught, and particularly the forest type. For this species, vinegar and wine positively enhanced the trap efficiency, with vinegar also being more efficient than wine in this case; in addition, mixed forest enhanced the traps’ efficiency (Table 7). The conditional effect of each covariate is depicted in Figure 6.
4. Discussion
The survey allowed for the collection of thirteen different species among Scolytinae, Nitidulidae, and Scarabaeidae. It is striking that no other beetle family responded positively to the attractants, suggesting that neither the type of traps nor the attractants were adequate; it is also plausible that the sampling season was too late compared with most of the species’ phenology. If we exclude Scolytinae, represented by two native species which were almost exclusively collected with alcohol baited traps, both Nitidulidae and Scarabaeidae were included as invasive species. Four out of the eight species of Nitidulidae belong to invasive taxa, representing the 90% of the total number of the nitidulid specimens. Carpophilus lugubris is one of the ten Carpophilus species indicated as invasive in Italy [40]; however, it most probably is the only species present in the area, given its recent introduction and expansion throughout Veneto and Friuli Venezia Giulia [41,42]. Epuraea luteola and Epuraea guttata are two of the three invasive Epuraea occurring in Italy, but the only occur in Lombardy, as the third species is limited to southern Italy [40]. Glischrochilus quadrisignatus is the only invasive species of the genus in Italy [40]. Stelidota geminata (Say, 1825), the only other invasive nitidulid present in Lombardy [43], was not recorded in our survey. Popillia japonica is the only invasive Scarabaeidae in Italy [16]; this species alone constituted about 97% of the scarabaeoid specimens.
The fact that this simple trial was able to detect almost, if not all, of the exotic Nitidulidae and Scarabaeidae present in the area suggests, at least for these two families, how home-made traps baited with food products, using apple cider vinegar in particular, may be effective at detecting exotic and invasive beetles, and could be an important addition to monitoring plans around entry points. Long standing food-baited traps are very effective in catching beetles and, in several cases, they have to be modified in order not to kill rare and threatened species (e.g., [44,45]). The proportion of exotic compared with native species collected in our trial suggests that minimizing the activity period of the traps does not substantially affect the traps’ capacity in catching non-native species. Trapping efficiency against exotic species seems to be substantially affected by trap placement (margin vs. interior of forest), absolute maximum temperature, and mean humidity; however, this result may be substantially biased by the two overrepresented E. ocularis and P. japonica. A similar effect may be caused by X. saxesenii on the native species pool. As expected, Scolytinae was the only group responding systematically to alcohol traps, and the fact that the native X. saxesenii constituted almost all of the individuals collected is perfectly in line with the tendencies of this species to respond to a wide range of ethanol concentrations [46]. Furthermore, the efficiency when catching X. saxesenii is not affected by the environmental covariates, and is most probably attributable to the species dispersal capabilities, long phenology, and polyphagy.
Nitidulidae is a group of primary interest, given the number of exotic and invasive species introduced in Europe [40]. The effectiveness of vinegar traps in rapidly detecting exotic sap beetles may serve as an important tool in monitoring new introductions or the spread of newly acclimatized species. Fermented baits are commonly used to investigate Nitidulidae [47,48]; the capability of E. luteola, E. ocularis, C. lugubris, and G. quadrisignatus to rapidly respond to wine and vinegar seems promising in using bottle traps for their capture; furthermore, given the similar ecological niche that these have with most of the others invasive species, we may expect a similar attraction efficiency by both attractants. In addition to direct damage to crops, Nitidulidae can be vectors of important pathogenic fungi such as Ophiostomatales [49] and Microascales [50]; consequently, their early detection may have a relevant role as phytosanitary security. The substantial effect given by the forest type in the capture efficiency of E. ocularis is probably attributable to its generalist habits and polyphagy, with adults able to feed on rotten fruit, flowers, sapping trees, and larvae developing in the fruit body of tree-fungi or decaying organic matter [40,51].
Popillia japonica is a highly polyphagous invasive pest outside its native range, so far limited to Lombardy and Piedmont in Italy [52]. Because of the substantial damage it is able to cause, as well as its excellent dispersal capacity, this species is currently controlled through mass trapping, with traps commonly baited with chemical attractants such as food-type volatile and sex pheromones [53,54,55]. The capture of this species with both wine and vinegar traps was unexpected and constituted an absolute novelty, as it is the first response of P. japonica towards these two attractants, vinegar in particular. Scarabaeidae Rutelinae are rarely collected with fruit/fermented and baited traps in the Palearctic [56,57], while they constitute a substantial fraction of the tropics’ trapped biomass [58]. However, a particular sensitivity of P. japonica towards volatile compounds released during fruit ripening or rotting can be deduced from Hammons et al. [59,60], where the species have been repeatedly documented as feeding on grapes in the USA. Concerning environmental covariates, the trapping efficiency against P. japonica is substantially affected by forest type, forest cover condition, and absolute maximum temperature. Forest composition and trap placement (forest edge/interior) have a significant effect, as P. japonica prefers ecotones, areas hosting the greater variety of feeding plants, and where the species move by flying. Furthermore, ecotonal areas may present grass patches suitable for egg deposition and larval development [61]. The positive effect of high temperatures in increasing trapping efficiency is defined by a general increase of P. japonica activity combined with a greater attractant volatility [62].
Vinegar efficiency against Nitidulidae and P. japonica in heterogeneous habitats suggests a good effectiveness of the traps in agricultural and peri-urban environments, as well as other anthropized entry points such as airports and ports. Furthermore, adopting short trapping sections repeated over time seems to not affect native beetles through an unnecessary over-trapping.
5. Conclusions
The traps used in this study can be produced in a short time, using recycled or easily available materials. Furthermore, as the traps are light and compact, they can be comfortably managed by one person, even in large numbers. The attractants are readily available at a low cost, making this survey technique very useful in citizen science projects for large-scale monitoring projects. Apple cider vinegar has proven to be the most affordable and still the most efficient bait, capable of attracting exotic and invasive beetles in almost all conditions; wine has intermediate attractivity and may be used as a coadjutant of vinegar; conversely, in our context, high-grade alcohol targets exclusively native Scolytinae. As what has been presented here is only a first trial, it will certainly be interesting to replicate the monitoring using different types of vinegar or wine and to evaluate the influence of other potential co-factors, including seasonality. It would be interesting to identify and test new types of inexpensive attractants for other families of beetles other than those used in this study.
As citizen science data are not usually collected following a sampling design typical of standardized monitoring programs, we stress that the collection of environmental variables, beside the collection of the specimens, is crucial for a correct interpretation of the records. Specifically, the position of the traps is essential information that must be provided by citizens involved in monitoring activities, along with the baits used and the time of exposure of the traps. In particular, if these monitoring projects are directed against invasive species, citizen scientists should keep in mind that the time exposure of baited traps should be minimized in order to reduce the impact on native fauna.
Given the strong attractivity of vinegar against P. japonica, it is plausible that the massive trapping of this species through a supervised citizen science action may become a substantial contribution and integration to the control strategies developed by local phytosanitary institutions, especially in cultivated and suburban areas.
Author Contributions
Conceptualization, E.R. and L.B.; methodology, E.R. and A.G.; formal analysis, L.B.; investigation, A.G.; data curation, E.R., A.G., and L.B.; writing—original draft preparation, E.R. and L.B.; writing—review and editing, E.R., A.G., and L.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The original dataset is available under request.
Acknowledgments
The authors thanks Robert Haack (USDA), Gareth Powell (Brigham Young University), Massimo Faccoli (Università degli Studi di Padova), and Jorge Ari Noriega (Universidad de los Andes) for providing valuable comments and recommendations that improved the quality of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Roques, A. Taxonomy, Time and Geographic Patterns. Chapter 2. BioRisk 2010, 4, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Ratti, E. Coleotteri alieni in Italia/Alien Coleoptera in Italy. Vers. 2007-05-25. Available online: https://msn.visitmuve.it/it/ricerca/settori/entomologia/progetti/coleotteri-alieni-in-italia-1957-2006/lista/ (accessed on 22 April 2021).
- Cocquempot, C.; Lindelöw, Å. Longhorn beetles (Coleoptera, Cerambycidae). Chapter 8.1. BioRisk 2010, 4, 193–218. [Google Scholar] [CrossRef]
- Sauvard, D.; Branco, M.; Lakatos, F.; Faccoli, M.; Kirkendall, L. Weevils and Bark Beetles (Coleoptera, Curculionoidea). Chapter 8.2. BioRisk 2010, 4, 219–266. [Google Scholar] [CrossRef]
- Beenen, R.; Roques, A. Leaf and Seed Beetles (Coleoptera, Chrysomelidae). Chapter 8.3. BioRisk 2010, 4, 267–292. [Google Scholar] [CrossRef]
- Roy, H.; Migeon, A. Ladybeetles (Coccinellidae). Chapter 8.4. BioRisk 2010, 4, 293–313. [Google Scholar] [CrossRef] [Green Version]
- Denux, O.; Zagatti, P. Coleoptera Families Other than Cerambycidae, Curculionidae sensu lato, Chrysomelidae sensu lato, and Coccinellidae. Chapter 8.5. BioRisk 2010, 4, 315–406. [Google Scholar] [CrossRef] [Green Version]
- Montagna, M.; Zoia, S.; Leonardi, C.; Di Taddeo, V.; Caldara, R.; Sassi, D. Colasposoma dauricum Mannerheim, 1849 an Asian species adventive to Piedmont, Italy (Coleoptera: Chrysomelidae: Eumolpinae). Zootaxa 2016, 4097, 127–129. [Google Scholar] [CrossRef]
- Rassati, D.; Lieutier, F.; Faccoli, M. Alien wood-boring beetles in Mediterranean regions. In Insects and Diseases of Mediterranean For. Systems; Paine, T.D., Lieutier, F., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 293–327. [Google Scholar] [CrossRef]
- Binazzi, F.; Del Nista, D.; Sabbatini Peverieri, G.; Marianelli, L.; Roversi, P.F.; Pennacchio, F. Saperda tridentata Olivier (Coleoptera Cerambycidae Lamiinae): Continuous interceptions at the italian port of Livorno represent a growing challenge for phytosanitary services. Redia 2019, 102, 171–176. [Google Scholar] [CrossRef]
- Ruzzier, E.; Colla, A. Micromalthus debilis LeConte, 1878 (Coleoptera: Micromalthidae), an American wood-boring beetle new to Italy. Zootaxa 2019, 4623, 589–594. [Google Scholar] [CrossRef]
- Nardi, G.; Bocci, M.; Poggi, R. Ptilodactyla exotica Chapin, 1927 in Italy (Coleoptera: Ptilodactylidae). Zootaxa 2020, 4861, 63–70. [Google Scholar] [CrossRef]
- Ruzzier, E.; Morin, L.; Glerean, P.; Forbicioni, L. New and Interesting Records of Coleoptera from Northeastern Italy and Slovenia (Alexiidae, Buprestidae, Carabidae, Cerambycidae, Ciidae, Curculionidae, Mordellidae, Silvanidae). Coleopt. Bull. 2020, 74, 523–531. [Google Scholar] [CrossRef]
- Stoett, P.; Roy, H.E.; Pauchard, A. Invasive alien species and planetary and global health policy. LANCET Planet. Health 2019, 3, 400–401. [Google Scholar] [CrossRef] [Green Version]
- Meurisse, N.; Rassati, D.; Hurley, B.P.; Brockerhoff, E.G.; Haack, R.A. Common pathways by which non-native forest insects move internationally and domestically. J. Pest Sci. 2019, 92, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Ruzzier, E.; Tomasi, F.; Poso, M.; Martinez-Sañudo, I. Archophileurus spinosus Dechambre, 2006 (Coleoptera: Scarabaeidae: Dynastinae), a new exotic scarab possibly acclimatized in Italy, with a compilation of exotic Scarabaeidae found in Europe. Zootaxa 2020, 4750, 577–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Poland, T.M.; Rassati, D. Improved Biosecurity Surveillance of Non-Native Forest Insects: A Review of Current Methods. J. Pest Sci. 2019, 92, 37–49. [Google Scholar] [CrossRef]
- Marchioro, M.; Rassati, D.; Faccoli, M.; Rooyen, K.V.; Kostanowicz, C.; Webster, V.; Mayo, P.; Sweeney, J. Maximizing bark and ambrosia beetle (Coleoptera: Curculionidae) catches in trapping surveys for longhorn and jewel beetles. J. Econ. Entomol. 2020, 113, 2745–2757. [Google Scholar] [CrossRef]
- Hamilton, R.M.; Foster, R.E.; Gibb, T.J.; Sadof, C.S.; Holland, J.D.; Engel, B.A. Distribution and Dynamics of Japanese Beetles Along the Indianapolis Airport Perimeter and the Influence of Land Use on Trap Catch. Environ. Entomol. 2007, 36, 287–296. [Google Scholar] [CrossRef]
- Rassati, D.; Faccoli, M.; Petrucco Toffolo, E.; Battisti, A.; Marini, L.; Clough, Y. Improving the early detection of alien wood-boring beetles in ports and surrounding forests. J. Appl. Ecol. 2015, 52, 50–58. [Google Scholar] [CrossRef]
- Faccoli, M.; Gallego, D.; Branco, M.; Brockerhoff, E.G.; Corley, J.; Coyle, D.R.; Hurley, B.P.; Hervé, J.; Lakatos, F.; Lantschner, V.; et al. A first worldwide multispecies survey of invasive Mediterranean pine bark beetles (Coleoptera: Curculionidae, Scolytinae). Biol. Invasions 2020, 22, 1785–1799. [Google Scholar] [CrossRef]
- Rizzo, D.; Luchi, N.; Da Lio, D.; Bartolini, L.; Nugnes, F.; Cappellini, G.; Bruscoli, T.; Salemi, C.; Griffo, R.V.; Garonna, A.P.; et al. Development of a loop-mediated isothermal amplification (LAMP) assay for the identification of the invasive wood borer Aromia bungii (Coleoptera: Cerambycidae) from frass. 3 Biotech 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Tobin, P.C.; Strom, B.L.; Francese, J.A.; Herms, D.A.; McCullough, G.D.; Poland, T.M.; Ryall, K.L.; Scarr, T.; Silk, P.J.; Thistle, H.W. Evaluation of Trapping Schemes to Detect Emerald Ash Borer (Coleoptera: Buprestidae). J. Econ. Entomol. 2021, toab065. [Google Scholar] [CrossRef]
- Caley, P.; Welvaert, M.; Barry, S.C. Crowd Surveillance: Estimating Citizen Science Reporting Probabilities for Insects of Biosecurity Concern: Implications for Plant Biosecurity Surveillance. J. Pest Sci. 2020, 93, 543–550. [Google Scholar] [CrossRef] [Green Version]
- Ruzzier, E.; Menchetti, M.; Bortolotti, L.; Selis, M.; Monterastelli, M.; Forbicioni, L. Updated Distribution of the Invasive Megachile sculpturalis (Hymenoptera: Megachilidae) in Italy and Its First Record on a Mediterranean Island. Biodivers. Data J. 2020, 8, e57783. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A.; Vikhrev, N.E.; Esin, M.N. The use of simple crown traps for the insects collection. Nat. Conserv. Res. 2020, 5, 87–108. [Google Scholar] [CrossRef] [Green Version]
- Touroult, J.; Witté, I. Beer, Wine, or Fruit Juice: Which Is Best? A Case Study of Bait Efficiency to Sample Saproxylic Beetles (Coleoptera) in an Oak Woodland. Coleopt. Bull. 2020, 74, 763–771. [Google Scholar] [CrossRef]
- Carvalho, A.G.; Trevisan, H. A New Trap Model for Scolytinae and Platypodinae (Insecta, Coleoptera). Floresta Ambiente 2015, 22, 575–578. [Google Scholar] [CrossRef]
- Steininger, M.S.; Hulcr, J.; Šigut, M.; Lucky, A. Simple and Efficient Trap for Bark and Ambrosia Beetles (Coleoptera: Curculionidae) to Facilitate Invasive Species Monitoring and Citizen Involvement. J. Econ. Entomol. 2015, 108, 1115–1123. [Google Scholar] [CrossRef] [Green Version]
- Rosenberger, D.W.; Aukema, B.H. Stimulating curiosity and engagement with insects beyond the college classroom through citizen science. Am. Entomol. 2016, 62, 120–122. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.C.; Hanks, L.M. Influence of fermenting bait and vertical position of traps on attraction of cerambycid beetles to pheromone lures. J. Econ. Entomol. 2016, 109, 2145–2150. [Google Scholar] [CrossRef]
- Ruchin, A.B.; Egorov, L.V.; Khapugin, A.A. Usage of Fermental Traps for the Study of the Species Diversity of Coleoptera. Insects 2021, 12, 407. [Google Scholar] [CrossRef] [PubMed]
- Uso e copertura del suolo in Regione Lombardia. Available online: https://www.geoportale.regione.lombardia.it/download-ricerca (accessed on 10 October 2020).
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002; pp. 1–497. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 10 October 2020).
- Delignette-Muller, M.L.; Dutang, C. fitdistrplus: An R package for fitting distributions. J. Stat. Softw. 2015, 64, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Lüdecke, D. sjPlot: Data Visualization for Statistics in Social Science. R Package Version 2.8.6. 2020. Available online: https://CRAN.R-project.org/package=sjPlot (accessed on 10 October 2020).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; pp. 1–260. [Google Scholar]
- Jelínek, J.; Audisio, P.; Hajek, J.; Baviera, C.; Moncourtier, B.; Barnouin, T.; Brustel, H.; Genç, H.; Leschen, R.A. Epuraea imperialis (Reitter, 1877). New invasive species of Nitidulidae (Coleoptera) in Europe, with a checklist of sap beetles introduced to Europe and Mediterranean areas. AAPP Phys. Math. Nat. Sci. 2016, 94, 1–24. [Google Scholar]
- Marini, F.; Mutinelli, F.; Montarsi, F.; Cline, A.R.; Gatti, E.; Audisio, P. First report in Italy of the dusky sap beetle, Carpophilus lugubris, a new potential pest for Europe. J. Pest Sci. 2013, 86, 157–160. [Google Scholar] [CrossRef]
- Audisio, P.; Marini, F.; Gatti, E.; Montarsi, F.; Mutinelli, F.; Campanaro, A.; Cline, A.R. A scientific note on rapid host shift of the invasive dusky sap-beetle (Carpophilus lugubris) in Italian beehives: New commensal or potential threat for European apiculture? Apidologie 2014, 45, 464–466. [Google Scholar] [CrossRef] [Green Version]
- Grottolo, M. Indagine sulla entomofauna della riserva naturale “della rocca, del sasso e parco lacuale” (Manerba del Garda, Lombardia). Nat. Brescia. Annu. Mus. Civ. Stor. Nat. Brescia 2020, 43, 73–117. [Google Scholar]
- Torres-Vila, L.M.; Sanchez-González, Á.; Ponce-Escudero, F.; Martín-Vertedor, D.; Ferrero-Garcia, J.J. Assessing mass trapping efficiency and population density of Cerambyx welensii Küster by mark-recapture in dehesa open woodlands. Eur. J. For. Res. 2012, 131, 1103–1116. [Google Scholar] [CrossRef]
- Bardiani, M.; Tini, M.; Carpaneto, G.M.; Audisio, P.; Bussola, E.; Campanaro, A.; Cini, A.; Maurizi, E.; Mason, F.; Sabbatini Peverieri, G.; et al. Effects of trap baits and height on stag beetle and flower chafer monitoring: Ecological and conservation implications. J. Insect Conserv. 2017, 21, 157–168. [Google Scholar] [CrossRef]
- Chen, Y.; Coleman, T.W.; Ranger, C.M.; Seybold, S.J. Differential flight responses of two ambrosia beetles to ethanol as indicators of invasion biology: The case with Kuroshio shot hole borer (Euwallacea kuroshio) and fruit-tree pinhole borer (Xyleborinus saxesenii). Ecol. Entomol. 2021, 52, 243–259. [Google Scholar] [CrossRef]
- Powell, G.S. A checklist of the sap beetle (Coleoptera: Nitidulidae) fauna of Indiana, with notes on effective trapping methods. Insecta Mundi 2015, 424, 1–9. [Google Scholar]
- Hadden, R.A.; Saxton, N.A.; Gerlach, P.S.; Nielson, P.L.; Brown, S.D.; Bybee, S.M.; Powell, G.S. Nitidulidae (Coleoptera: Cucujoidea) of Vanuatu. J. Asia-Pac. Entomol. 2020, 23, 470–476. [Google Scholar] [CrossRef]
- Musvuugwa, T.; Dreyer, L.L.; Roets, F. Future danger posed by fungi in the Ophiostomatales when encountering new hosts. Fungal Ecol. 2016, 22, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Jagemann, S.M.; Juzwik, J.; Tobin, P.C.; Raffa, K.F. Seasonal and regional distributions, degree-day models, and phoresy rates of the major sap beetle (Coleoptera: Nitidulidae) vectors of the oak wilt fungus, Bretziella fagacearum, in Wisconsin. Environ. Entomol. 2018, 47, 1152–1164. [Google Scholar] [CrossRef]
- Cline, A.R.; Audisio, P. Epuraea (Haptoncus) ocularis Fairmaire (Coleoptera: Nitidulidae) recently found in the USA, with comments on Nearctic members of Epuraea Erichson. Coleopt. Bull. 2011, 6, 24–26. [Google Scholar] [CrossRef]
- Popillia japonica (POPIJA). Available online: https://gd.eppo.int/taxon/POPIJA (accessed on 22 April 2021).
- Piñero, J.C.; Dudenhoeffer, A.P. Mass trapping designs for organic control of the Japanese beetle, Popillia japonica (Coleoptera: Scarabaeidae). Pest Manag. Sci. 2018, 74, 1687–1693. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); Schrader, G.; Camilleri, M.; Ciubotaru, R.M.; Diakaki, M.; Vos, S. Pest survey card on Popillia japonica. EFSA Supporting Publ. 2019, 16, 1568E. [Google Scholar] [CrossRef] [Green Version]
- Shanovich, H.N.; Dean, A.N.; Koch, R.L.; Hodgson, E.W. Biology and management of Japanese beetle (Coleoptera: Scarabaeidae) in corn and soybean. J. Integr. Pest Manag. 2019, 10, 9. [Google Scholar] [CrossRef]
- Tezcan, S.; Pehlivan, E. Evaluation of the Lucanoidea and Scarabaeoidea (Coleoptera) fauna of ecological cherry orchards in İzmir and Manisa provinces of Turkey. Ziraat Fak. Derg. 2001, 3, 31–38. [Google Scholar]
- Voigt, E.; Tóth, M. Three years of mass trapping with sex attractant traps for control of Anomala Scarabs in ripening peaches. IOBC wprs Bull. 2004, 27, 69–76. [Google Scholar]
- Puker, A.; Correa, A.C.M.; Butzske, L.S.; Pacheco, R.A. Using aerial fruit-baited traps with different naturally fermented baits to survey scarab beetles in the Amazon rainforest. Stud. Neotrop. Fauna Environ. 2020, 1–6. [Google Scholar] [CrossRef]
- Hammons, D.L.; Kurtural, S.K.; Potter, D.A. Japanese beetles facilitate feeding by green June beetles (Coleoptera: Scarabaeidae) on ripening grapes. Environ. Entomol. 2008, 37, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Hammons, D.L.; Kurtural, S.K.; Newman, M.C.; Potter, D.A. Invasive Japanese beetles facilitate aggregation and injury by a native scarab pest of ripening fruits. Proc. Natl. Acad. Sci. USA 2009, 106, 3686–3691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, D.A.; Held, D.W. Biology and management of the Japanese beetle. Annu. Rev. Entomol. 2002, 47, 175–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, A.R.; Cole, W.H. The response of Popillia japonica to light and the Weber-Fechner law. J. Gen. Physiol. 1921, 3, 331–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Study area. Map illustrating land use classes and the sampling localities in the surrounding of the Malpensa International Airport, MXP (Ferno and Somma Lombardo municipalities, Varese Province, Italy); numbers indicate the 13 sampling sites where the trial was performed.
Figure 1.
Study area. Map illustrating land use classes and the sampling localities in the surrounding of the Malpensa International Airport, MXP (Ferno and Somma Lombardo municipalities, Varese Province, Italy); numbers indicate the 13 sampling sites where the trial was performed.

Figure 2.
Beetle families collected during the survey; total number of individuals given in brackets.
Figure 2.
Beetle families collected during the survey; total number of individuals given in brackets.

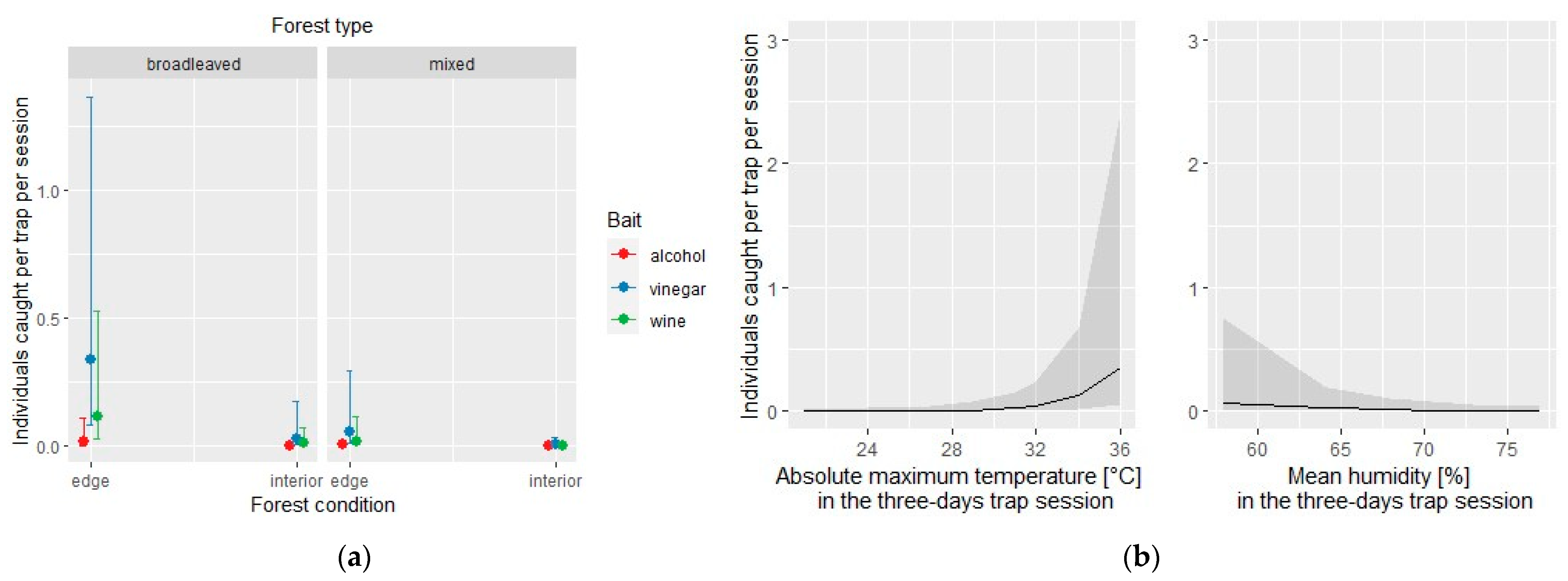
Figure 3.
Exotic species: effect of the covariates of the negative binomial regression model affecting the number of individuals caught by each of the three traps per site and per session; (a) conditional effect of habitat covariates (forest type: broadleaved vs. mixed; forest condition: edge vs. interior); (b) main effect of meteorological covariates.
Figure 3.
Exotic species: effect of the covariates of the negative binomial regression model affecting the number of individuals caught by each of the three traps per site and per session; (a) conditional effect of habitat covariates (forest type: broadleaved vs. mixed; forest condition: edge vs. interior); (b) main effect of meteorological covariates.

Figure 4.
Native species: effect of the covariates of the negative binomial regression model affecting the number of individuals caught by each of the three traps per site and per session; (a) conditional effect of habitat covariates (forest type: broadleaved vs. mixed; forest condition: edge vs. interior); (b) main effect of meteorological covariates.
Figure 4.
Native species: effect of the covariates of the negative binomial regression model affecting the number of individuals caught by each of the three traps per site and per session; (a) conditional effect of habitat covariates (forest type: broadleaved vs. mixed; forest condition: edge vs. interior); (b) main effect of meteorological covariates.

Figure 5.
Popillia japonica: effect of the covariates of the negative binomial regression model affecting the number of individuals caught by each of the three traps per site and per session; (a) conditional effect of habitat covariates (forest type: broadleaved vs. mixed; forest condition: edge vs. interior); (b) main effect of meteorological covariates.
Figure 5.
Popillia japonica: effect of the covariates of the negative binomial regression model affecting the number of individuals caught by each of the three traps per site and per session; (a) conditional effect of habitat covariates (forest type: broadleaved vs. mixed; forest condition: edge vs. interior); (b) main effect of meteorological covariates.

Figure 6.
Epuraea ocularis: effect of the covariates of the negative binomial regression model affecting the number of individuals caught by each of the three traps per site and per session; (a) conditional effect of habitat covariates (forest type: broadleaved vs. mixed; forest condition: edge vs. interior); (b) main effect of meteorological covariates.
Figure 6.
Epuraea ocularis: effect of the covariates of the negative binomial regression model affecting the number of individuals caught by each of the three traps per site and per session; (a) conditional effect of habitat covariates (forest type: broadleaved vs. mixed; forest condition: edge vs. interior); (b) main effect of meteorological covariates.

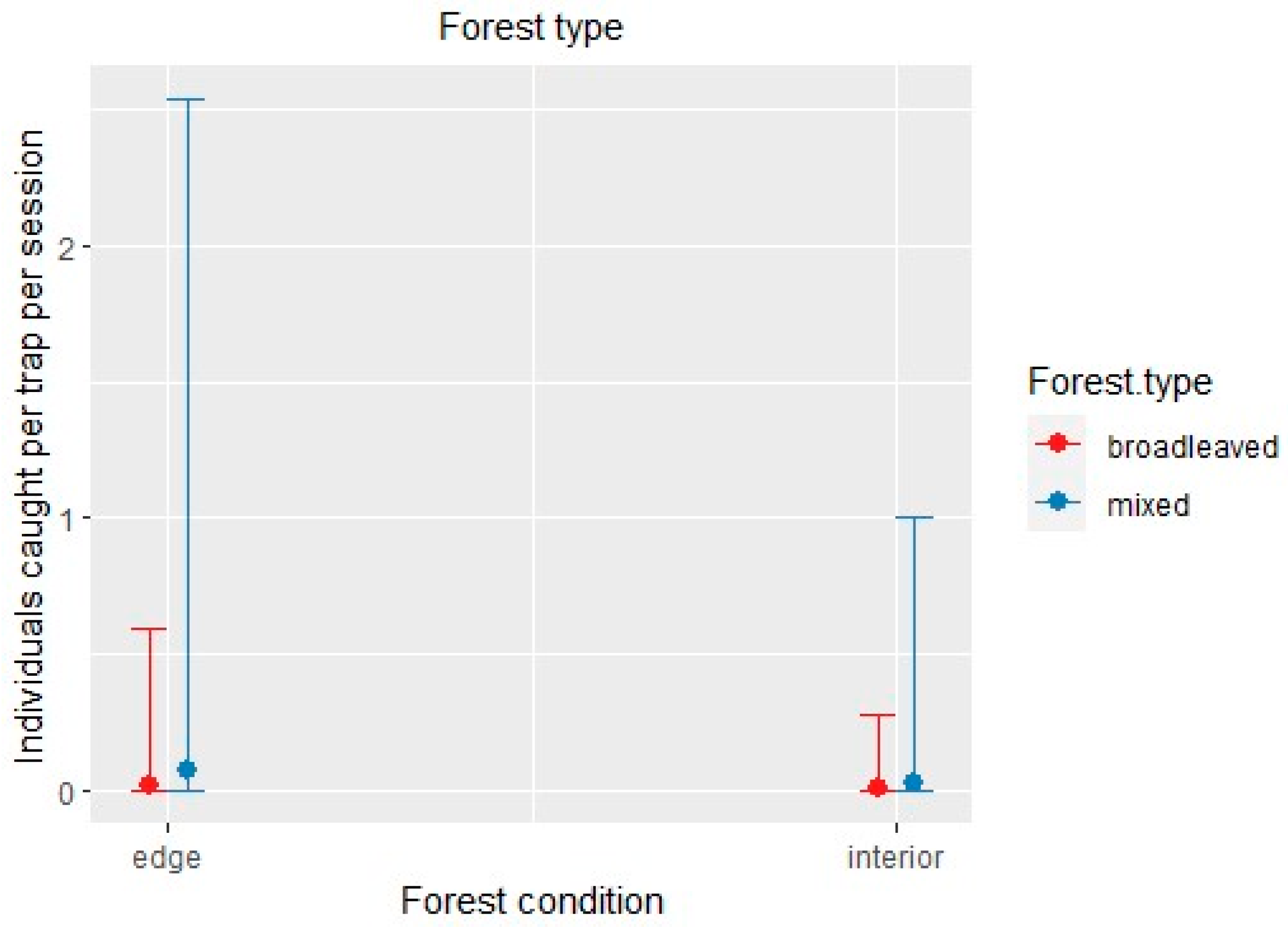
Figure 7.
Xyleborinus saxesenii: effect of the negative binomial regression model’s covariates affecting the number of individuals caught by the trap triggered with alcohol per site and session. Only the conditional effect of habitat covariates (forest type: broadleaved vs. mixed; forest condition: edge vs. interior) is shown, as neither of the two meteorological variables showed significant effects.
Figure 7.
Xyleborinus saxesenii: effect of the negative binomial regression model’s covariates affecting the number of individuals caught by the trap triggered with alcohol per site and session. Only the conditional effect of habitat covariates (forest type: broadleaved vs. mixed; forest condition: edge vs. interior) is shown, as neither of the two meteorological variables showed significant effects.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Location of the sampling sites in which the group of three traps was triggered with different baits. The forest covers in a buffer of 250 m around the sampling site used to define the forest conditions (interior condition: forest cover >90%) and forest type.
Table 1.
Location of the sampling sites in which the group of three traps was triggered with different baits. The forest covers in a buffer of 250 m around the sampling site used to define the forest conditions (interior condition: forest cover >90%) and forest type.
| Site Number | Nord (WGS84/UTM Zone 32N) | East (WGS84/UTM Zone 32N) | Forest Cover (250-m Buffer) | Forest Condition | Forest Type |
|---|---|---|---|---|---|
| 1 | 5,053,409 | 476,170 | 98% | interior | broadleaved |
| 2 | 5,055,451 | 476,779 | 100% | interior | mixed |
| 3 | 5,057,292 | 477,621 | 58% | edge | broadleaved |
| 4 | 5,056,847 | 478,549 | 52% | edge | mixed |
| 5 | 5,055,783 | 478,335 | 87% | edge | mixed |
| 6 | 5,052,549 | 480,498 | 79% | edge | broadleaved |
| 7 | 5,052,204 | 480,113 | 88% | edge | broadleaved |
| 8 | 5,053,714 | 480,070 | 76% | edge | mixed |
| 9 | 5,054,249 | 480,724 | 99% | interior | mixed |
| 10 | 5,050,105 | 478,906 | 82% | edge | broadleaved |
| 11 | 5,049,577 | 478,796 | 100% | interior | broadleaved |
| 12 | 5,056,516 | 479,151 | 100% | interior | mixed |
| 13 | 5,054,071 | 479,998 | 99% | interior | mixed |
Table 2.
Meteorological data obtained from the airport station for each of the three-days trapping sessions.
Table 2.
Meteorological data obtained from the airport station for each of the three-days trapping sessions.
| Session Number | Placement Date | Control Date | Abs. Min. Temp. (°C) 3-Day Session | Abs. Max. Temp. (°C) 3-Day Session | Mean Temp. (°C) 3-Day Session | Mean Humidity3-Days Session |
|---|---|---|---|---|---|---|
| 1 | 31 July 2020 | 3 August 2020 | 18 | 36 | 26.3 | 64.0 |
| 2 | 7 August 2020 | 10 August 2020 | 17 | 34 | 26.5 | 58.0 |
| 3 | 14 August 2020 | 17 August 2020 | 18 | 32 | 24.5 | 68.0 |
| 4 | 28 August 2020 | 31 August 2020 | 13 | 27 | 21.0 | 77.0 |
| 5 | 4 September 2020 | 7 September 2020 | 14 | 29 | 20.8 | 73.0 |
| 6 | 11 September 2020 | 14 September 2020 | 17 | 31 | 23.3 | 70.5 |
| 7 | 25 September 2020 | 28 September 2020 | 4 | 21 | 13.0 | 64.3 |
Table 3.
Coleoptera species collected during the survey. (V—Vinegar; W—wine; E—Alcohol.)
| Species | Family/Subfamily | Origin | Status | Attractant | N° of Individuals |
|---|---|---|---|---|---|
| Carpophilus lugubris Murray 1864 | Nitidulidae | Nearctic | Invasive | V(3); W(2) | 5 |
| Cryptarcha strigata (Fabricius, 1787) | Nitidulidae | W-Palaearctic | Native | W(1) | 1 |
| Epuraea guttata (Olivier, 1811) | Nitidulidae | W-Palaearctic | Native | V(6); W(1) | 7 |
| Epuraea luteola (Erichson, 1843) | Nitidulidae | E-Palaearctic | Invasive | V(5) | 5 |
| Epuraea ocularis (Fairmaire, 1849) | Nitidulidae | E-Palaearctic | Invasive | V(176); W(28); E(1) | 205 |
| Epuraea unicolor (Olivier, 1790) | Nitidulidae | Palaearctic | Native | V(5) | 5 |
| Glischrochilus quadrisignatus (Say, 1835) | Nitidulidae | Nearctic | Invasive | V(3); W(1) | 4 |
| Soronia grisea (Linnaeus, 1758) | Nitidulidae | Palaearctic | Native | V(2); W(9) | 11 |
| Anisandrus dispar Fabricius, 1792 | Scolytinae | Palaearctic | Native | E(4) | 4 |
| Xyleborinus saxesenii (Ratzeburg, 1837) | Scolytinae | Palaearctic | Native | W(1) E(581) | 59 |
| Cetonia aurata (Linnaeus, 1758) | Scarabaeidae | W-Palaearctic | Native | W(1) | 1 |
| Popillia japonica (Newman, 1838) | Scarabaeidae | E-Palaearctic | Invasive | V(161); W(51); E(6) | 218 |
| Potosia cuprea (Fabricius, 1775) | Scarabaeidae | W-Palaearctic | Native | W(1) | 1 |
| Protaetia speciosa (Adams, 1817) | Scarabaeidae | W-Palaearctic | Native | W(1) | 5 |
Table 4.
Exotic species. Effectiveness of baits in attracting individuals of all species pooled together, accounting for the effects of habitat and meteorological covariates considered in the negative binomial regression model.
Table 4.
Exotic species. Effectiveness of baits in attracting individuals of all species pooled together, accounting for the effects of habitat and meteorological covariates considered in the negative binomial regression model.
| Covariates | Estimate | SE of Estimate | z-Value | p |
|---|---|---|---|---|
| (intercept) | −1.657 | 2.998 | −0.553 | 0.581 |
| Bait: vinegar | 4.311 | 0.584 | 7.379 | <0.001 |
| Bait: wine | 2.705 | 0.594 | 4.552 | <0.001 |
| Forest type: mixed forests | 0.201 | 0.367 | 0.549 | 0.583 |
| Forest cover condition: interior | −0.579 | 0.371 | −1.559 | 0.119 |
| Absolute maximum temperature | 0.127 | 0.044 | 2.901 | 0.004 |
| Mean humidity | −0.075 | 0.033 | −2.296 | 0.022 |
Table 5.
Native species. Effectiveness of baits in attracting individuals of all species pooled together, accounting for the effects of habitat and meteorological covariates considered in the negative binomial regression model.
Table 5.
Native species. Effectiveness of baits in attracting individuals of all species pooled together, accounting for the effects of habitat and meteorological covariates considered in the negative binomial regression model.
| Covariates | Estimate | SE of Estimate | z-Value | p |
|---|---|---|---|---|
| (intercept) | −6.772 | 3.344 | −2.025 | 0.042 |
| Bait: vinegar | −1. 187 | 0.479 | −2.479 | 0.013 |
| Bait: wine | −0.986 | 0.467 | −2.110 | 0.035 |
| Forest type: mixed forests | 0.345 | 0.403 | 0.857 | 0.391 |
| Forest cover condition: interior | −0.411 | 0.406 | −1.012 | 0.312 |
| Absolute maximum temperature | 0.039 | 0.050 | 0.786 | 0.431 |
| Mean humidity | 0.073 | 0.036 | 2.038 | 0.042 |
Table 6.
Popillia japonica. Effectiveness of baits in attracting individuals of the species, accounting for the effects of habitat and meteorological covariates considered in the negative binomial regression model.
Table 6.
Popillia japonica. Effectiveness of baits in attracting individuals of the species, accounting for the effects of habitat and meteorological covariates considered in the negative binomial regression model.
| Covariates | Estimate | SE of Estimate | z-Value | p |
|---|---|---|---|---|
| (intercept) | −9.751 | 9.453 | −1.032 | 0.302 |
| Bait: vinegar | 3.136 | 0.922 | 3.401 | 0.001 |
| Bait: wine | 2.047 | 0.940 | 2.178 | 0.029 |
| Forest type: mixed forests | −1.827 | 0.680 | −2.685 | 0.007 |
| Forest cover condition: interior | −2.574 | 0.795 | −3.237 | 0.001 |
| Absolute maximum temperature | 0.519 | 0.172 | 3.012 | 0.003 |
| Mean humidity | −0.147 | 0.078 | −1.893 | 0.058 |
Table 7.
Epuraea ocularis. Effectiveness of baits in attracting individuals of the species, accounting for the effects of habitat and meteorological covariates considered in the negative binomial regression model.
Table 7.
Epuraea ocularis. Effectiveness of baits in attracting individuals of the species, accounting for the effects of habitat and meteorological covariates considered in the negative binomial regression model.
| Covariates | Estimate | SE of Estimate | z-Value | p |
|---|---|---|---|---|
| (intercept) | −8.111 | 3.705 | −2.189 | 0.029 |
| Bait: vinegar | 5.382 | 1.115 | 4.829 | <0.001 |
| Bait: wine | 3.410 | 1.129 | 3.019 | 0.003 |
| Forest type: mixed forests | 1.083 | 0.466 | 2.327 | 0.020 |
| Forest cover condition: interior | −0.392 | 0.461 | −0.851 | 0.395 |
| Absolute maximum temperature | −0.016 | 0.052 | −0.311 | 0.756 |
| Mean humidity | 0.051 | 0.040 | 1.292 | 0.196 |
Table 8.
Xyleborinus saxesenii. Effectiveness of alcohol in attracting individuals of the species, accounting for the effects of habitat and meteorological covariates considered in the negative binomial regression model.
Table 8.
Xyleborinus saxesenii. Effectiveness of alcohol in attracting individuals of the species, accounting for the effects of habitat and meteorological covariates considered in the negative binomial regression model.
| Covariates | Estimate | SE of Estimate | z-Value | p |
|---|---|---|---|---|
| (intercept) | −64.815 | 45.690 | −1.419 | 0.156 |
| Forest type: mixed forests | 1.544 | 0.860 | 1.796 | 0.073 |
| Forest cover condition: interior | −0.972 | 0.861 | −1.129 | 0.259 |
| Absolute maximum temperature | 0.711 | 0.596 | 1.194 | 0.233 |
| Mean humidity | 0.597 | 0.390 | 1.530 | 0.126 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ruzzier, E.; Galli, A.; Bani, L. Monitoring Exotic Beetles with Inexpensive Attractants: A Case Study. Insects 2021, 12, 462. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050462
AMA Style
Ruzzier E, Galli A, Bani L. Monitoring Exotic Beetles with Inexpensive Attractants: A Case Study. Insects. 2021; 12(5):462. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050462
Chicago/Turabian StyleRuzzier, Enrico, Andrea Galli, and Luciano Bani. 2021. "Monitoring Exotic Beetles with Inexpensive Attractants: A Case Study" Insects 12, no. 5: 462. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050462
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.