
Morphometric Analysis of Coptotermes spp. Soldier Caste (Blattodea: Rhinotermitidae) in Indonesia and Evidence of Coptotermes gestroi Extreme Head-Capsule Shapes

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Collection and Identification
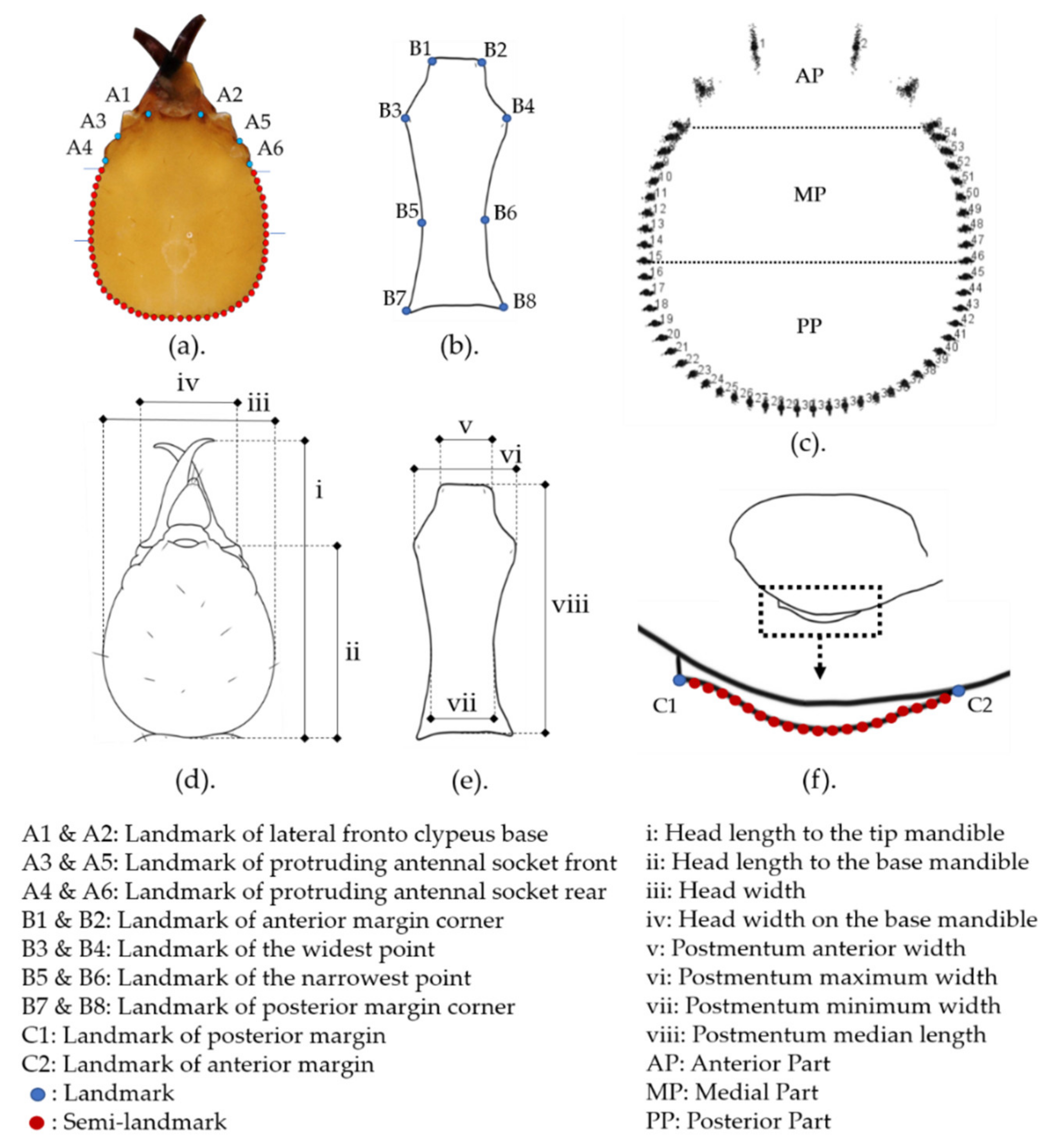
2.2. Linear Morphometric Analysis
2.3. Geometric Morphometric Analysis
2.4. Statistical Analysis
2.5. Phylogenetic Analysis
3. Results
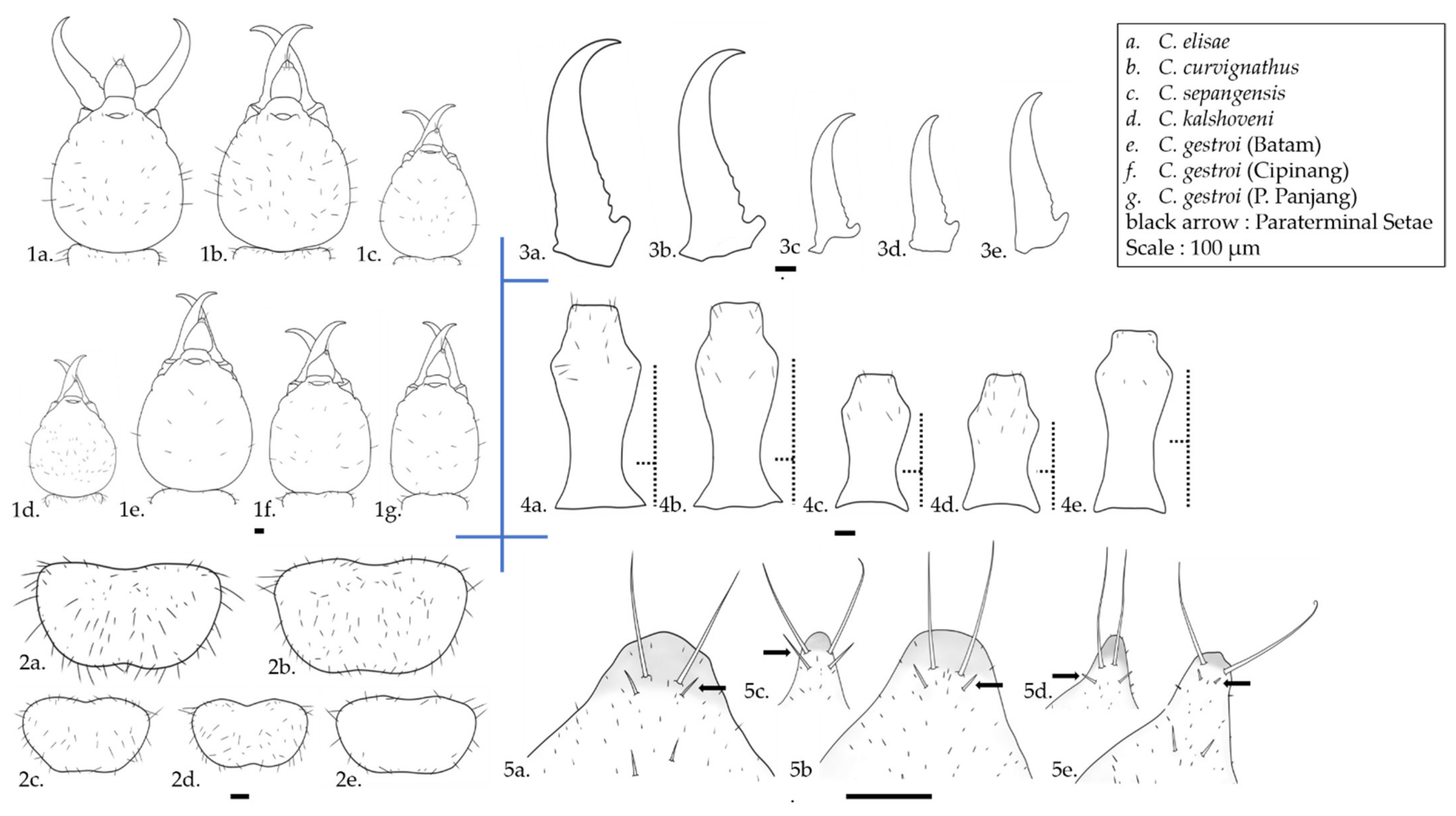
3.1. Soldier Morphology Characteristics of Coptotermes (Wasmann)
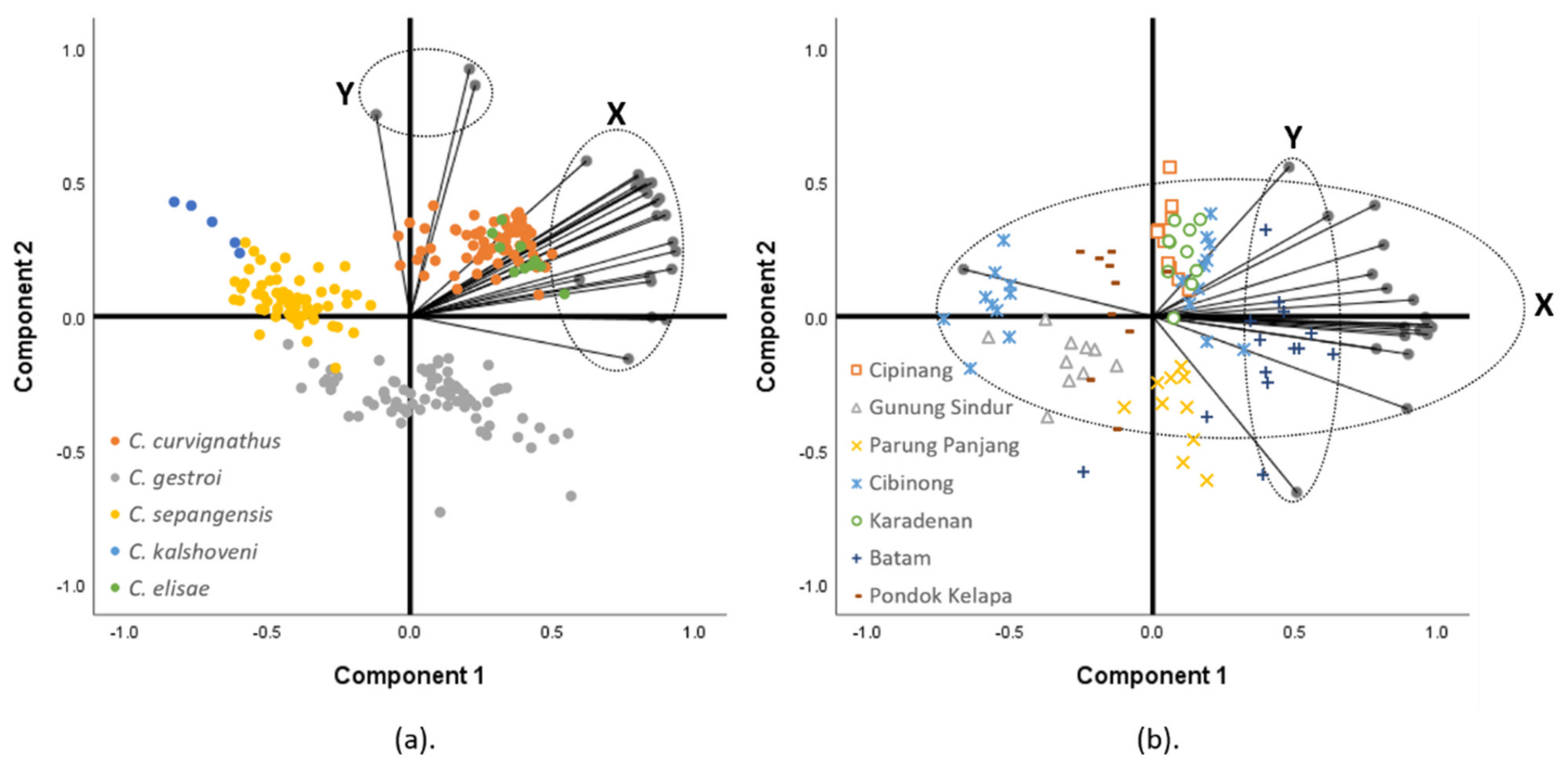
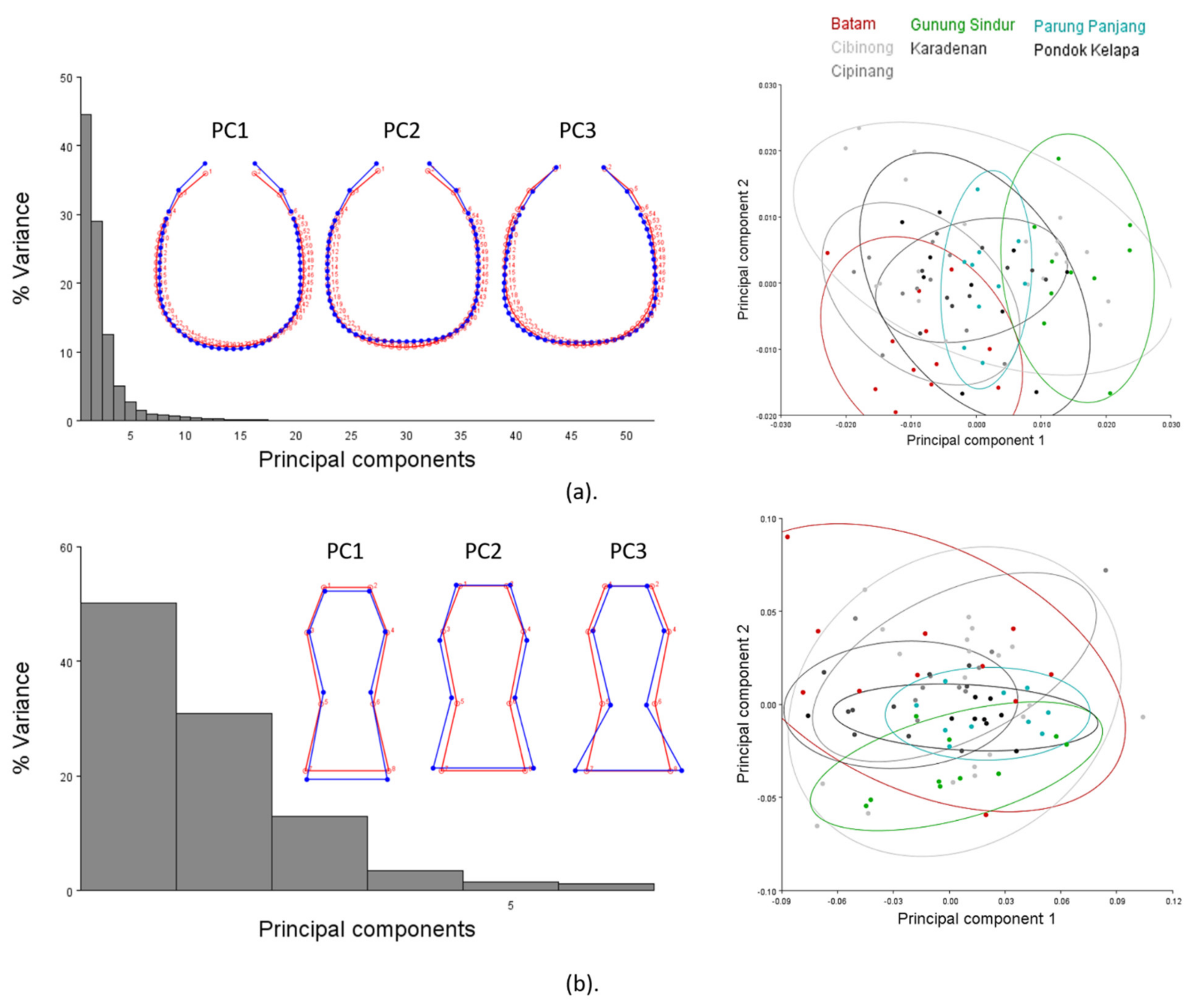
3.2. Linear Morphometrics
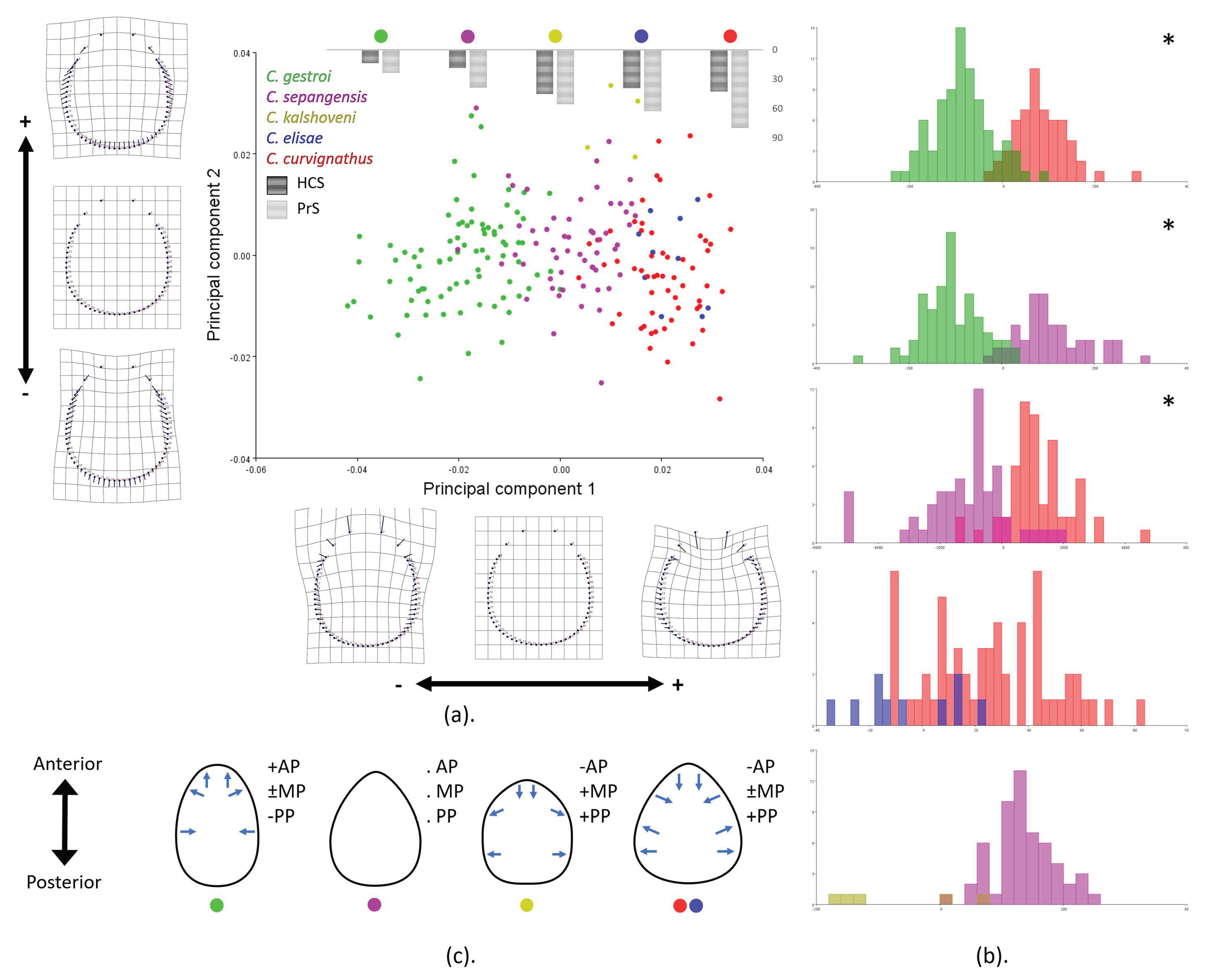
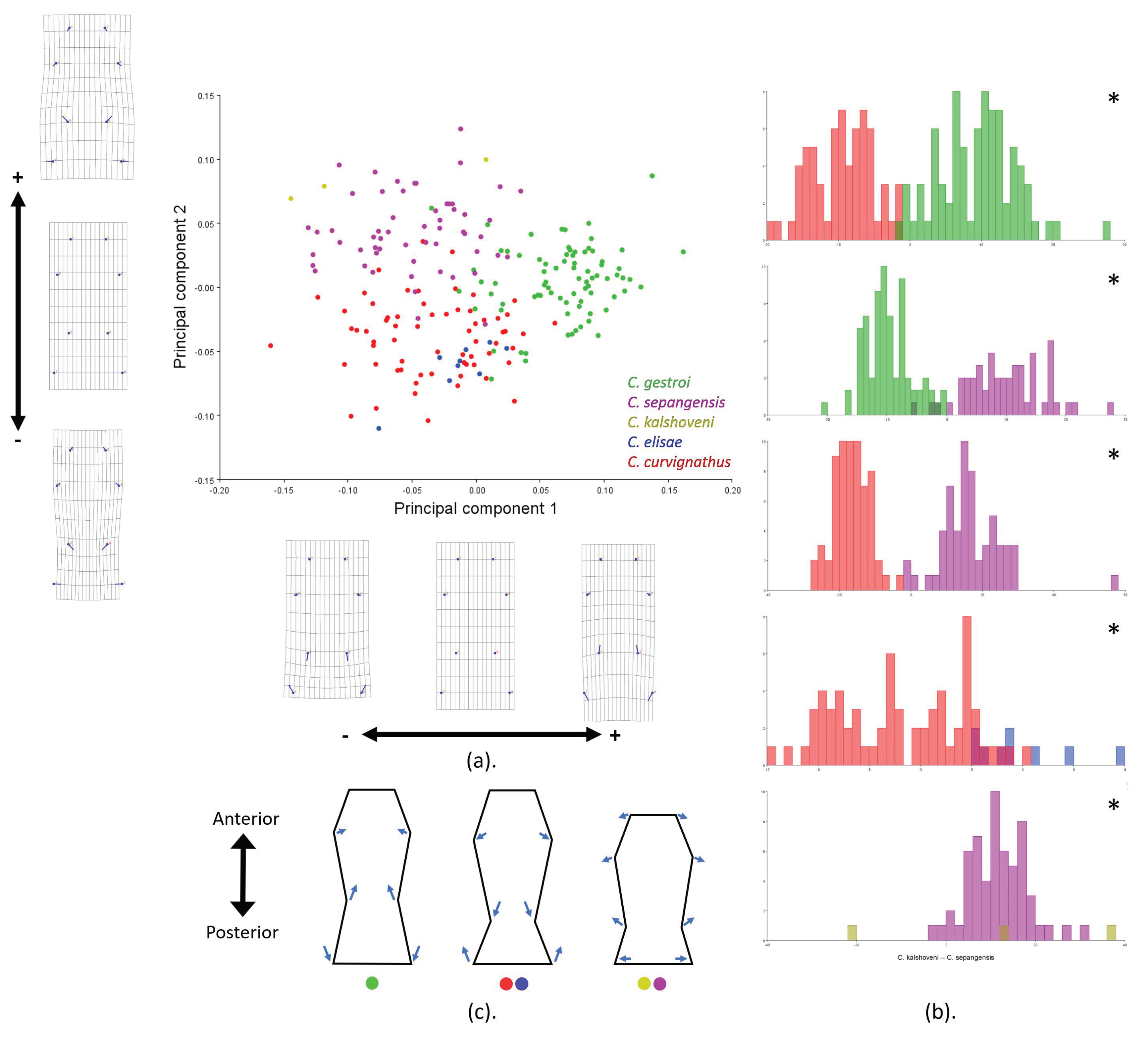
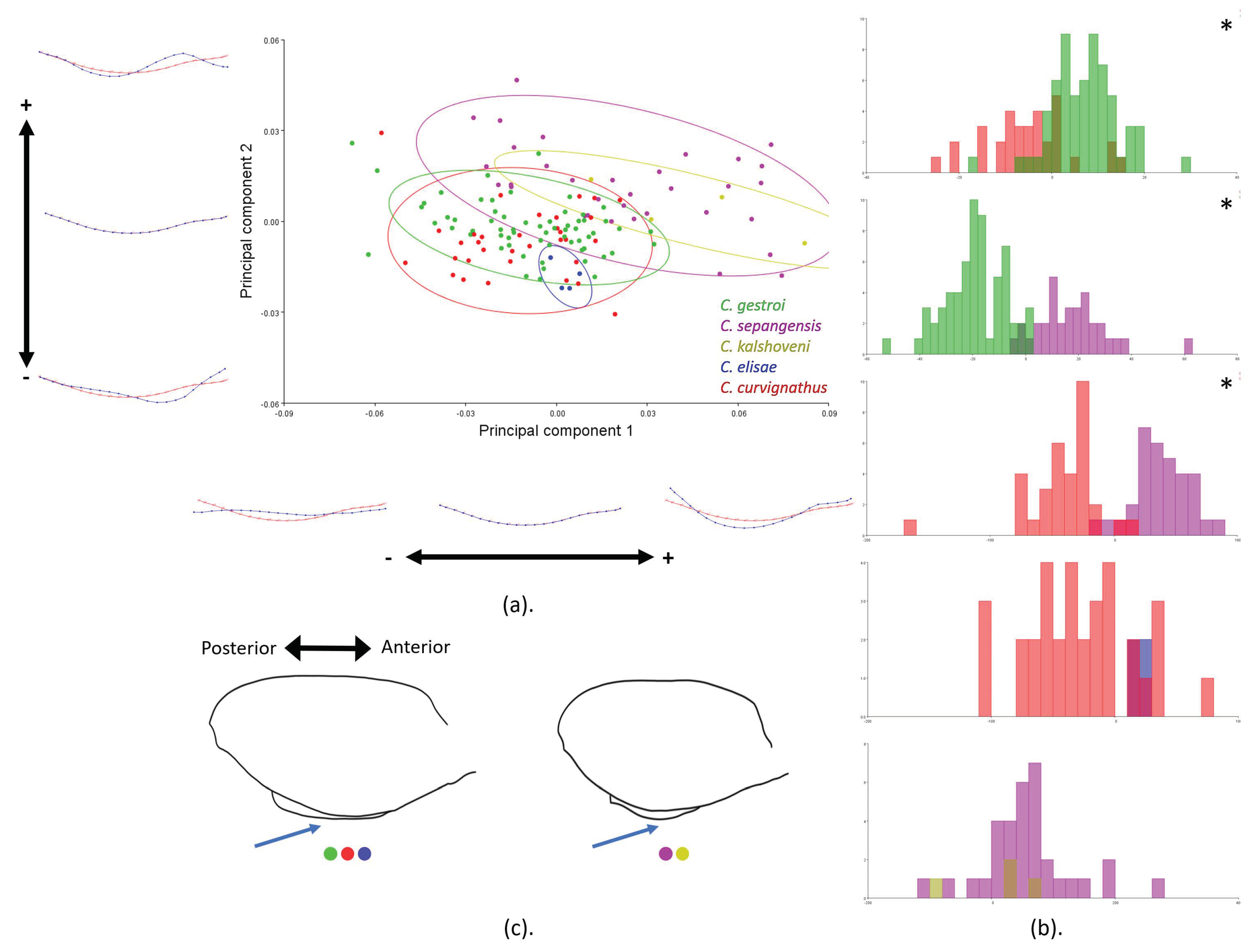
3.3. Geometric Morphometrics
3.4. Pairwise Distance of 12 s and 16 s Genes
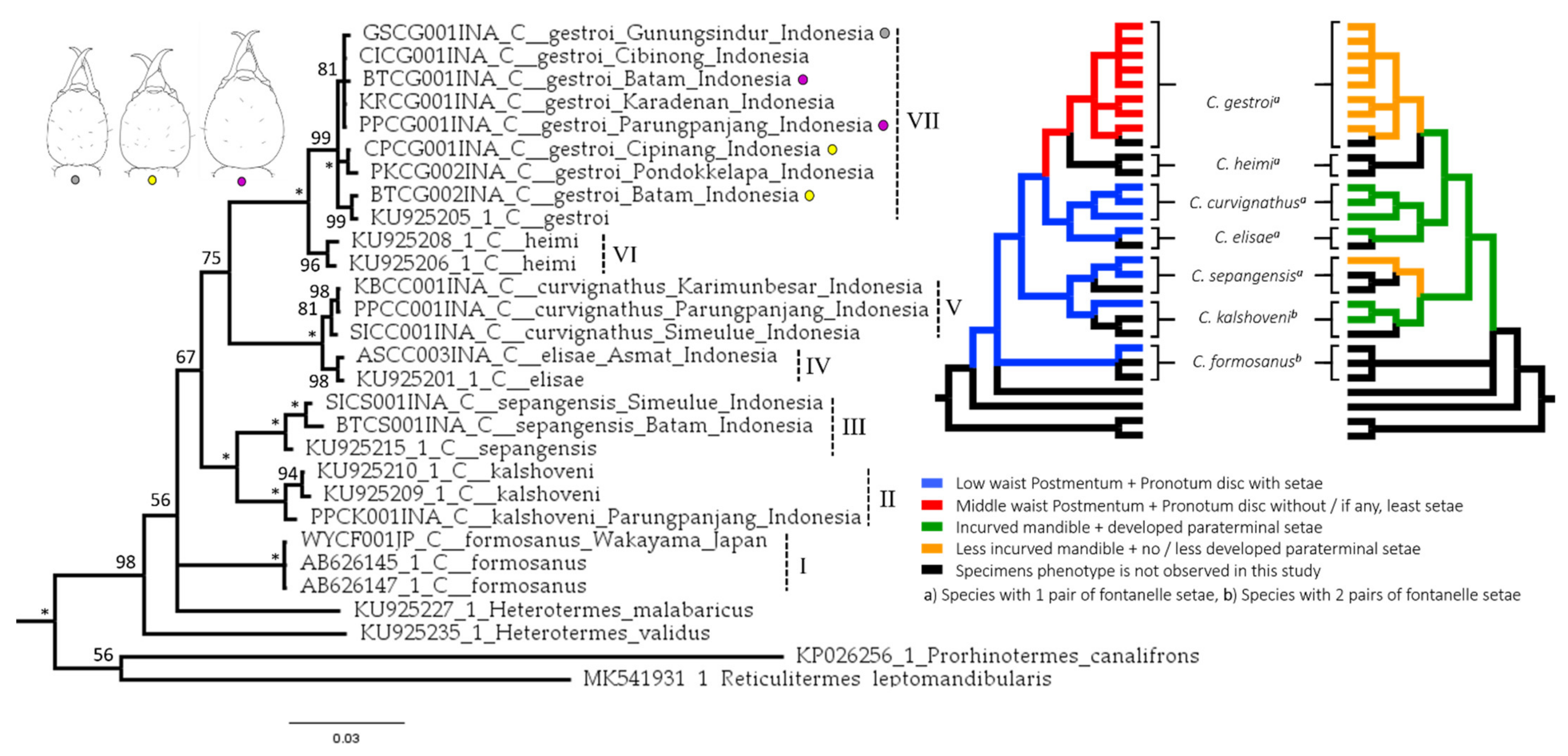
3.5. Phylogenetic Relationship Inferred from Mitochondrial Genes
4. Discussion
4.1. Important Noticeable Characteristics
4.2. Putative Function-Related Shape and Size Variation of HC and PS
4.3. Impact of Coptotermes Head Shape Perplexity on Indonesia Termite Pest Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abe, T.; Higashi, M. Cellulose Centered Perspective on Terrestrial Community Structure. Oikos 1991, 60, 127–133. [Google Scholar] [CrossRef]
- Lee, K.E.; Wood, T.G. Termites and Soils; Academic Press: London, UK, 1971; p. 279. [Google Scholar]
- Li, H.-F.; Fujisaki, I.; Su, N.-Y. Predicting Habitat Suitability of Coptotermes gestroi (Isoptera: Rhinotermitidae) with Species Distribution Models. J. Econ. Entomol. 2013, 106, 311–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, L.M.; Brown, J.L.; Yoder, A.D. Integrating Statistical Genetic and Geospatial Methods Brings New Power to Phylogeography. Mol. Phylogenet. Evol. 2011, 59, 523–537. [Google Scholar] [CrossRef]
- Constantino, R. The Pest Termites of South America: Taxonomy, Distribution and Status. J. Appl. Entomol. 2002, 126, 355–365. [Google Scholar] [CrossRef]
- Hapukotuwa, N.K.; Grace, J.K. Preferences of Coptotermes formosanus Shiraki and Coptotermes gestroi (Wasmann) (Blattodea: Rhinotermitidae) Among Three Commercial Wood Species. Insects 2011, 2, 499–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.-Y. Control of Foraging Colonies of Subterranean Termites, Coptotermes travians (Isoptera: Rhinotermitidae) in Malaysia Using Hexaflumuron Baits. Sociobiology 2002, 39, 411–416. [Google Scholar]
- Kirton, Y.P.; Kirton, L.G. A Note on a Survey of Termite Attack in Bahau Conifer Plantation. J. Trop. For. Sci. 1998, 10, 564–567. [Google Scholar]
- Jones, D.T.; Prasetyo, A.H. A Survey of the Termites (Insecta: Isoptera) of Tabalong District, South Kalimantan, Indonesia. Raffles Bull. Zool. 2002, 50, 117–128. [Google Scholar]
- Chouvenc, T.; Li, H.-F.; Austin, J.; Bordereau, C.; Bourguignon, T.; Cameron, S.L.; Cancello, E.M.; Constantino, R.; Costa-Leonardo, A.M.; Eggleton, P.; et al. Revisiting Coptotermes (Isoptera: Rhinotermitidae): A Global Taxonomic Road Map for Species Validity and Distribution of an Economically Important Subterranean Termite Genus. Syst. Entomol. 2016, 41, 299–306. [Google Scholar] [CrossRef]
- Lee, C.-Y. Subterranean Termite Pests and Their Control in the Urban Environment in Malaysia. Sociobiology 2002, 40, 3–10. [Google Scholar]
- Rahman, H.; Fernandez, K.; Arumugam, N. Termites Infesting Malaysian Forests: Case Study from Bornean Forest, Sabah, Malaysia. In Termites and Sustainable Management: Volume 2—Economic Losses and Management; Khan, M.A., Ahmad, W., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 97–118. [Google Scholar] [CrossRef]
- Gray, B.; Buchter, J. Termite Eradication in Araucaria Plantations in New Guinea. Commonw. For. Rev. 1969, 48, 201–207. [Google Scholar]
- Lemmens, R.H.M.J.; Soerianegara, I.; Wong, W.C. Plant Resources of South-East Asia. No. 5(2). Timber Trees: Minor Commercial Timbers; Backhuys Publisher: Leiden, The Netherlands, 1995; Volume 5, p. 651. [Google Scholar]
- Wikantyoso, B.; Himmi, S.K.; Yusuf, S. A Case Report of Termite Attack on Mango Fruit: Plasticity in Feeding? In Proceedings of the National Seminar of Entomological Society of Indonesia, Sumedang, Indonesia, 10 October 2019; pp. 1–6. [Google Scholar]
- Bourguignon, T.; Roisin, Y. Revision of the Termite Family Rhinotermitidae (Isoptera) in New Guinea. ZooKeys 2011, 148, 55–103. [Google Scholar] [CrossRef]
- Li, H.-F.; Kanzaki, N.; Su, N.-Y. Redescription of the Drywood Termite Incisitermes inamurae (Isoptera: Kalotermitidae) from Southern Taiwan. Ann. Entomol. Soc. Am. 2009, 102, 759–765. [Google Scholar] [CrossRef] [Green Version]
- Marquina, D.; Buczek, M.; Ronquist, F.; Łukasik, P. The Effect of Ethanol Concentration on the Morphological and Molecular Preservation of Insects for Biodiversity Studies. bioRxiv 2020. [Google Scholar] [CrossRef]
- Koshikawa, S.; Matsumoto, T.; Miura, T. Morphometric Changes During Soldier Differentiation of the Damp-wood Termite Hodotermopsis japonica (Isoptera, Termopsidae). Insectes Sociaux 2002, 49, 245–250. [Google Scholar] [CrossRef]
- Kaji, T.; Keiler, J.; Bourguignon, T.; Miura, T. Functional Transformation Series and the Evolutionary Origin of Novel Forms: Evidence from a Remarkable Termite Defensive Organ. Evol. Dev. 2016, 18, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Hanh, T.V.; Hien, N.T.; Yen, N.; Hu-Án, T.T. Applying Molecular Technology to Identify Termite Species of the Genus Coptotermes in the Hanoi Old Quarter. In Proceedings of the 10th Pacific-Termite Research Group Conference, Kuala Lumpur, Malaysia, 26–28 February 2014. S1-1. [Google Scholar]
- Kirton, L.G.; Brown, V.K. The Taxonomic Status of Pest Species of Coptotermes in Southeast Asia: Resolving the Paradox in the Pest Status of the Termites, Coptotermes gestroi, C. havilandi and C. travians (Isoptera: Rhinotermitidae). Sociobiology 2003, 42, 43–63. [Google Scholar]
- Ahmad, M. Key to the Indomalayan Termites. Biologia 1958, 4, 1–193. [Google Scholar]
- Roonwal, M.L.; Chhotani, O.B. Termite Fauna of Assam Region, Eastern India; Zoological Survey of India: Calcutta, India, 1962; Volume 28, p. 406. [Google Scholar]
- Roonwal, M.L.; Maiti, P.K. Termite from Indonesia Including West Irian. Treubia 1966, 27, 63–140. [Google Scholar]
- Roonwal, M.L.; Chhotani, O.B. Fauna of India; the Director of Zoological Survey of India, Ed.; Zoological Survey of India: Calcutta, India, 1989; Volume 1, p. 665. [Google Scholar]
- Tho, Y.P. Termites of Peninsular Malaysia. Malay. For. Rec. 1992, 36, 1–224. [Google Scholar]
- Kushwaha, K.S. External Morphology of the Termite, Odontotermes obesus (Rambur) (Isoptera: Termitidae). Part 3. Chaetotaxy of the Soldier, Worker and Alate Castes. Rec. Indian Mus. 1960, 58, 71–114. [Google Scholar]
- Kushwaha, K.S. Chaetotaxy of the Termite, Odontotermes assmuthi Holmgren (Isoptera: Termitidae). Proc. Indian Acad. Sci. Sect. B 1961, 54, 130–137. [Google Scholar] [CrossRef]
- Takematsu, Y. The Genus Reticulitermes (Isoptera: Rhinotermitidae) in Japan, with Description of a New Species. Entomol. Sci. 1999, 2, 231–243. [Google Scholar]
- Constantino, R. Key to the Soldiers of South American Heterotermes with a New Species from Brazil (Isoptera: Rhinotermitidae). Insect Syst. Evol. 2000, 31, 463–472. [Google Scholar] [CrossRef]
- Kushwaha, K.S. Taxonomic Differentiation in Three Species of Odontotermes (Isoptera:Termitidae) as Based on Chaetotaxy. In Termites in the Humid Tropics: Proceedings of the New Delhi Symposium; Sharma, S.K., Ed.; UNESCO: Delhi, India, 1962; pp. 52–55. [Google Scholar]
- Gathorne-Hardy, F.J.; Collins, N.; Buxton, R.D.; Eggleton, P. A Faunistic Review of the Termites (Insecta: Isoptera) of Sulawesi, Including an Updated Checklist of the Species. Malay. Nat. J. 2000, 54, 347–353. [Google Scholar]
- Gathorne-Hardy, F.J.; Jones, D.T.; Mawdsley, N.A. The Recolonization of the Krakatau Islands by Termites (Isoptera), and Their Biogeographical Origins. Biol. J. Linn. Soc. 2000, 71, 251–267. [Google Scholar] [CrossRef]
- Krishna, K.; Grimaldi, D.A.; Krishna, V.; Engel, M.S. Treatise on the Isoptera of the World. Bull. Am. Mus. Nat. Hist. 2013, 377, 627–971. [Google Scholar]
- Mutanen, M.; Pretorius, E. Subjective Visual Evaluation vs. Traditional and Geometric Morphometrics in Species Delimitation: A Comparison of Moth Genitalia. Syst. Entomol. 2007, 32, 371–386. [Google Scholar] [CrossRef]
- Jost, J. Relations and Dependencies between Morphological Characters. Theory Biosci. 2017, 136, 69–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seilacher, A.; Gishlick, A.D. Morphodynamics; CRC Press: Boca Raton, FL, USA, 2014; p. 551. [Google Scholar] [CrossRef]
- Añez, N.; Valenta, D.; Cazorla, D.; Quicke, D.; Feliciangeli, M. Multivariate Analysis To Discriminate Species of Phlebotomine Sand Flies (Diptera: Psychodidae): Lutzomyia townsendi, L. spinicrassa, and L. youngi. J. Med. Entomol. 1997, 34, 312–316. [Google Scholar] [CrossRef]
- Yeap, B.-K.; Dugal, F.M.; Othman1, A.S.; Lee, C.-Y. Genetic Relationship Between Coptotermes heimi and Coptotermes gestroi (Isoptera: Rhinotermitidae). Sociobiology 2010, 56, 291–311. [Google Scholar]
- Atmaja, V.; Hamidy, A.; Arisuryanti, T.; Matsui, M.; Smith, E. A New Species of Microhyla (Anura: Microhylidae) from Sumatra, Indonesia. Treubia 2019, 45, 25–46. [Google Scholar] [CrossRef]
- Tatsuta, H.; Takahashi, K.H.; Sakamaki, Y. Geometric Morphometrics in Entomology: Basics and Applications. Entomol. Sci. 2018, 21, 164–184. [Google Scholar] [CrossRef] [Green Version]
- Cooke, S.B.; Terhune, C.E. Form, Function, and Geometric Morphometrics. Anat. Rec. 2015, 298, 5–28. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P. MorphoJ: An Integrated Software Package for Geometric Morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.P.; Barluenga, M.; Meyer, A. Shape Analysis of Symmetric Structures: Quantifying Variation Among Individuals and Asymmetry. Evolution 2002, 56, 1909–1920. [Google Scholar] [CrossRef] [Green Version]
- James Rohlf, F.; Marcus, L.F. A Revolution Morphometrics. Trends Ecol. Evol. 1993, 8, 129–132. [Google Scholar] [CrossRef]
- Savriama, Y. A Step-by-Step Guide for Geometric Morphometrics of Floral Symmetry. Front Plant Sci 2018, 9, 1433. [Google Scholar] [CrossRef] [Green Version]
- Lo, N.; Kitade, O.; Miura, T.; Constantino, R.; Matsumoto, T. Molecular Phylogeny of the Rhinotermitidae. Insectes Sociaux 2004, 51, 365–371. [Google Scholar] [CrossRef]
- Yeap, B.-K.; Othman, A.S.; Lee, V.S.; Lee, C.-Y. Genetic Relationship Between Coptotermes gestroi and Coptotermes vastator (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2007, 100, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Hill, G.F. Termites (Isoptera) from the Australian Region; Daw, H.E., Ed.; Government Printer: Melbourne, Australia, 1942.
- Ahmad, M. Termites (Isoptera) of Thailand; Bulletin of the American Museum of National History: New York, NY, USA, 1965; Volume 131. [Google Scholar]
- Thapa, R.S. Termites of Sabah (East Malaysia); Forestry Department: Sabah, Malaysia, 1982.
- Maiti, P.K. A Taxonomic Monograph on the World Species of Termites of the Family Rhinotermitidae (Isoptera: Insecta). In Memoirs of the Zoological Survey of India; Zoological Survey of India: Calcutta, India, 2006; Volume 20, p. 272. [Google Scholar]
- Parés-Casanova, P.M.; Salamanca-Carreño, A.; Crosby-Granados, R.A.; Bentez-Molano, J. A Comparison of Traditional and Geometric Morphometric Techniques for the Study of Basicranial Morphology in Horses: A Case Study of the Araucanian Horse from Colombia. Animals 2020, 10, 118. [Google Scholar] [CrossRef] [Green Version]
- Syaukani. A Guide to The Nasus Termites (Nasutitermitinae, Termitidae) of Kerinci Seblat Nasional Park, Sumatra; Yamane, S., Ed.; Nagao Natural Environment Foundation: Tokyo, Japan, 2010; p. 139. [Google Scholar]
- Li, H.-F.; Lan, Y.-C.; Su, N.-Y. Redescription of Prorhinotermes japonicus (Isoptera: Rhinotermitidae) From Taiwan. Ann. Entomol. Soc. Am. 2011, 104, 878–885. [Google Scholar] [CrossRef] [Green Version]
- Takematsu, Y.; Vongkulang, C. A Taxonomic Review of the Rhinotermitidae (Isoptera) of Thailand. J. Nat. Hist. 2012, 46, 1079–1109. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Choi, D.-S.; Ji, J.-Y.; Kim, N.; Han, J.M.; Park, S.-H.; Lee, S.; Seo, M.S.; Hwang, W.-J.; Forschler, B.T.; et al. A New Record of Reticulitermes kanmonensis Takematsu, 1999 (Isoptera: Rhinotermitidae) from Korea. J. Asia Pac. Entomol. 2015, 18, 351–359. [Google Scholar] [CrossRef]
- Smith, D.K. A Morphometric Analysis of Allosaurus. J. Vertebr. Paleontol. 1998, 18, 126–142. [Google Scholar] [CrossRef]
- Marramà, G.; Kriwet, J. Principal Component and Discriminant Analyses as Powerful Tools to Support Taxonomic Identification and Their Use for Functional and Phylogenetic Signal Detection of Isolated Fossil Shark Teeth. PLoS ONE 2017, 12, e0188806. [Google Scholar] [CrossRef] [PubMed]
- Rohlf, F.J. Tps Utility Program, 1.78; Ecology & Evolution and Anthropology, Stony Brook University: New York, NY, USA, 2019. [Google Scholar]
- Bookstein, F.L. Size and Shape Spaces for Landmark Data in Two Dimensions. Stat. Sci. 1986, 1, 181–222. [Google Scholar] [CrossRef]
- Bookstein, F.L. Landmark Methods for Forms Without Landmarks: Morphometrics of Group Differences in Outline Shape. Med. Image Anal. 1997, 1, 225–243. [Google Scholar] [CrossRef]
- Zelditch, M.L.; Swiderski, D.L.; Sheets, H.D.; Fink, W.L. Beyond Two-dimensional Configurations of Landmarks. In Geometric Morphometrics for Biologists; Zelditch, M.L., Swiderski, D.L., Sheets, H.D., Fink, W.L., Eds.; Academic Press: San Diego, CA, USA, 2004; pp. 385–407. [Google Scholar] [CrossRef]
- Rohlf, F.J. TpsDig2, 2.31; Ecology & Evolution and Anthropology, Stony Brook University: New York, NY, USA, 2017. [Google Scholar]
- Pietrusewsky, M.; Lauer, A.; Tsang, C.H.; Li, K.T.; Douglas, M.T. A Biodistance Analysis of Mandibles From Taiwan, Asia, and the Pacific: A Search for Polynesian Origins. In Biological Distance Analysis; Pilloud, M.A., Hefner, J.T., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 447–461. [Google Scholar] [CrossRef]
- Ramos, S.D.S.; Rickard Liow, S.J. Discriminant Function Analysis. In The Encyclopedia of Applied Linguistics; Chapelle, C.A., Ed.; John Wiley & Sons: Chichester, UK, 2012. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Size, Shape, and Form: Concepts of Allometry in Geometric Morphometrics. Dev. Genes Evol. 2016, 226, 113–137. [Google Scholar] [CrossRef]
- Kambhampati, S.; Smith, P.T. PCR Primers for the Amplification of Four Insect Mitochondrial Gene Fragments. Insect Mol. Biol. 1995, 4, 233–236. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X Windows Interface: Flexible Strategies for Multiple Sequence Alignment Aided by Quality Analysis Tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions Through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanabe, A.S. Kakusan: A Computer Program to Automate the Selection of a Nucleotide Substitution Model and the Configuration of a Mixed Model on Multilocus Data. Mol. Ecol. Notes 2007, 7, 962–964. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, And Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Hillis, D.M.; Bull, J.J. An Empirical Test of Bootstrapping as a Method for Assessing Confidence in Phylogenetic Analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Rannala, B. Frequentist Properties of Bayesian Posterior Probabilities of Phylogenetic Trees Under Simple and Complex Substitution Models. Syst. Biol. 2004, 53, 904–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leache, A.; Reeder, T. Molecular Systematics of the Eastern Fence Lizard (Sceloporus undulatus): A Comparison of Parsimony, Likelihood, and Bayesian Approaches. Syst. Biol. 2002, 51, 44–68. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drake, A.G.; Klingenberg, C.P. The Pace of Morphological Change: Historical Transformation of Skull Shape in St Bernard Dogs. Proc. R. Soc. B Biol. Sci. 2008, 275, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Tokuda, G.; Isagawa, H.; Sugio, K. The Complete Mitogenome of the Formosan Termite, Coptotermes formosanus Shiraki. Insectes Sociaux 2012, 59, 17–24. [Google Scholar] [CrossRef]
- Bourguignon, T.; Lo, N.; Šobotník, J.; Sillam-Dussès, D.; Roisin, Y.; Evans, T.A. Oceanic Dispersal, Vicariance and Human Introduction Shaped the Modern Distribution of the Termites Reticulitermes, Heterotermes and Coptotermes. Proc. R. Soc. B Biol. Sci. 2016, 283, 20160179. [Google Scholar] [CrossRef] [Green Version]
- Inward, D.J.G.; Vogler, A.P.; Eggleton, P. A Comprehensive Phylogenetic Analysis of Termites (Isoptera) Illuminates Key Aspects of Their Evolutionary Biology. Mol. Phylogenet. Evol. 2007, 44, 953–967. [Google Scholar] [CrossRef] [PubMed]
- Deligne, J.; Quennedey, A.; Blum, M.S. The Enemies and Defense Mechanisms of Termites. In Social Insects; Hermann, H.R., Ed.; Academic Press: London, UK, 1981; pp. 1–76. [Google Scholar] [CrossRef]
- Wasmann, E. Viaggio di Leonardo Fea in Birmania e Regioni Vicine, LXXII: Neue Termitophilen und Termiten Aus Indien. Ann. Mus. Civ. Stor. Nat. Genova 1896, 16, 613–630. [Google Scholar]
- Scheffrahn, R.; Carrijo, T.; Krecek, J.; Su, N.-Y.; Szalanski, A.; Austin, J.; Chase, J.; Mangold, J. A Single Endemic and Three Exotic Species of the Termite Genus Coptotermes (Isoptera, Rhinotermitidae) in the New World. Arthropod Syst. Phylogeny 2015, 73, 333–348. [Google Scholar]
- Scheffrahn, R.; Su, N.-Y. Asian Subterranean Termite, Coptotermes gestroi (=havilandi) (Wasmann) (Insecta: Blattodea: Rhinotermitidae). Available online: https://edis.ifas.ufl.edu/publication/IN285 (accessed on 18 May 2021).
- Li, H.F.; Yang, R.L.; Su, N.Y. Interspecific Competition and Territory Defense Mechanisms of Coptotermes formosanus and Coptotermes gestroi (Isoptera: Rhinotermitidae). Environ. Entomol. 2010, 39, 1601–1607. [Google Scholar] [CrossRef] [Green Version]
- Li, H.-F.; Ye, W.; Su, N.-Y.; Kanzaki, N. Phylogeography of Coptotermes gestroi and Coptotermes formosanus (Isoptera: Rhinotermitidae) in Taiwan. Ann. Entomol. Soc. Am. 2009, 102, 684–693. [Google Scholar] [CrossRef] [Green Version]
- Chouvenc, T.; Basille, M.; Li, H.F.; Su, N.Y. Developmental Instability in Incipient Colonies of Social Insects. PLoS ONE 2014, 9, e113949. [Google Scholar] [CrossRef] [Green Version]
- Quennedey, A.; Deligne, J. L’arme Frontale des Soldats de Termites. I. Rhinotermitidae. Insectes Sociaux 1975, 22, 243–267. [Google Scholar] [CrossRef]
- Chhotani, O.B. Termite Pest of Agriculture in the Indian Region and Their Control; Zoological Survey of India: Calcutta, India, 1980; Volume 4, pp. 29–33. [Google Scholar]
- Chen, G.-Y.; Ke, Y.-L.; Liang, W.-R.; Li, H.-F. Redescription of Formosan Subterranean Termite, Coptotermes formosanus (Blattodea: Rhinotermitidae), with Three New Synonyms from China. Acta Entomol. Musei Natl. Pragae 2020, 60, 599–608. [Google Scholar] [CrossRef]
- Kalshoven, L.G.E. Observations on Coptotermes havilandi Holmgr. (javanicus Kemn.) (Isoptera). Beaufortia 1962, 9, 121–137. [Google Scholar]
- Jamil, N.; Singham, V.; Wan Ismail, W.N. Morphometric Variation and Genetic Relationship of Coptotermes spp. (Blattodea: Rhinotermitidae) in Sarawak, Malaysia. Malays. Appl. Biol. J. 2018, 47, 113–125. [Google Scholar]
- Matsuura, K. Colony-Level Stabilization of Soldier Head Width for Head-Plug Defense in the Termite Reticulitermes speratus (Isoptera: Rhinotermitidae). Behav. Ecol. Sociobiol. 2002, 51, 172–179. [Google Scholar] [CrossRef]
- Hao, Y.-N.; Sun, Y.-X.; Liu, C.-Z. Functional Morphology of Antennae and Sensilla of Hippodamia variegata (Coleoptera: Coccinellidae). PLoS ONE 2020, 15, e0237452. [Google Scholar] [CrossRef] [PubMed]
- Parashar, B.; Chauhan, R.; Prakash, S.; Rao, K. Mechanotactile and Olfactory Antennal Sensilla in Four Species of Female Tabanids (Diptera). Ital. J. Zool. 1994, 61, 121–128. [Google Scholar] [CrossRef]
- Liu, Z.; Hua, B.-Z.; Liu, L. Ultrastructure of the Sensilla on Larval Antennae and Mouthparts in the Peach Fruit Moth, Carposina sasakii Matsumura (Lepidoptera: Carposinidae). Micron 2011, 42, 478–483. [Google Scholar] [CrossRef]
- Nowińska, A.; Brożek, J. Morphological Study of the Antennal Sensilla in Gerromorpha (Insecta: Hemiptera: Heteroptera). Zoomorphology 2017, 136, 327–347. [Google Scholar] [CrossRef] [Green Version]
- Hallberg, E.; Hansson, B.S. Arthropod Sensilla: Morphology and Phylogenetic Considerations. Microsc. Res. Tech. 1999, 47, 428–439. [Google Scholar] [CrossRef]
- Yanagawa, A.; Shimizu, S.; Noma, K.; Nishikawa, M.; Kazumasa, O.; Yokohari, F. Classification and Distribution of Antennal Sensilla of the Termite Coptotermes formosanus (Isoptera: Rhinotermitidae). Sociobiology 2009, 54, 327–349. [Google Scholar]
- Oberst, S.; Bann, G.; Lai, J.C.S.; Evans, T.A. Cryptic Termites Avoid Predatory Ants by Eavesdropping on Vibrational Cues from Their Footsteps. Ecol. Lett. 2017, 20, 212–221. [Google Scholar] [CrossRef]
- Evans, T.A.; Inta, R.; Lai, J.C.; Prueger, S.; Foo, N.W.; Fu, E.W.E.; Lenz, M. Termites Eavesdrop to Avoid Competitors. Proc. R. Soc. B Biol. Sci. 2009, 276, 4035–4041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romiti, F.; Redolfi DeZan, L.; Piras, P.; Carpaneto, G.M. Shape Variation of Mandible and Head in Lucanus cervus (Coleoptera: Lucanidae): A Comparison of Morphometric Approaches. Biol. J. Linn. Soc. 2016, 120, 1–16. [Google Scholar] [CrossRef]
- David, S.; Funken, J.; Potthast, W.; Blanke, A. Musculoskeletal Modelling Under an Evolutionary Perspective: Deciphering the Role of Single Muscle Regions in Closely Related Insects. J. R. Soc. Interface 2016, 13. [Google Scholar] [CrossRef] [PubMed]
- Blanke, A.; Pinheiro, M.; Watson, P.J.; Fagan, M.J. A Biomechanical Analysis of Prognathous and Orthognathous Insect Head Capsules: Evidence for a Many-to-one Mapping of Form to Function. J. Evol. Biol. 2018, 31, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Weihmann, T.; Wipfler, B. The Generalized Feeding Apparatus of Cockroaches: A Model for Biting and Chewing Insects; Springer: Cham, Switzerland, 2019; pp. 203–262. [Google Scholar] [CrossRef]
- Wipfler, B.; Weißing, K.; Klass, K.-D.; Weihmann, T. The Cephalic Morphology of the American Cockroach Periplaneta americana (Blattodea). Arthropod Syst. Phylogeny 2016, 74, 267–297. [Google Scholar]
- Mizuno, T.; Kojima, Y. A Blindsnake that Decapitates Its Termite Prey. J. Zool. 2015, 297, 220–224. [Google Scholar] [CrossRef]
- Arif, A.; Putri, G.; Lestari, P.I.; Widawati; Nurqalbi, M.; Saira, A. Diversity of Termite Rhinotermitidae (Isoptera, Insecta) in Educational Forest of Hasanuddin University. Perennial 2020, 16, 59–67. [Google Scholar] [CrossRef]
- Arif, A.; Putri, G.; Muin, M. Hazard Mapping of Subterranean Termite Attacks in Makassar City, South Sulawesi, Indonesia. Insects 2019, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Retmadhona, I.Y. Species Diversity, Spatial Distribution, and Intensity of Soil Termite Attack on Acacia Crassicarpa Stand on Peatlands; Institut Pertanian Bogor: Bogor, Indonesia, 2017. [Google Scholar]
- Subekti, N. Population Characteristics of Subterranean Termite Coptotermes spp (Blattodea: Rhinotermitidae) and the Impact of the Attack. Biosaintifika J. Biol. Biol. Educ. 2010, 2. [Google Scholar] [CrossRef]
- Arinana, A.; Aldina, R.; Nandika, D.; Rauf, A.; Harahap, I.S.; Sumertajaya, I.M.; Bahtiar, E.T. Termite Diversity in Urban Landscape, South Jakarta, Indonesia. Insects 2016, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Indrayani, Y. Short Communication: Diversity and distribution of termites in buildings in Pontianak City, West Kalimantan, Indonesia. Biodivers. J. Biol. Divers. 2017, 18. [Google Scholar] [CrossRef]
- Gallego-Abenza, M.; Mathevon, N.; Wheatcroft, D. Experience Modulates an Insect’s Response to Anthropogenic Noise. Behav. Ecol. 2019, 31, 90–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tricot, M.-C.C.; Cammaerts, R. Impact of Environmental Noise on Insects’ Physiology and Ethology-A Study on Ants as Models. Bioelectromagnetics 2018, 3, 1–8. [Google Scholar]
- Bowen, A.E.; Gurule-Small, G.A.; Tinghitella, R.M. Anthropogenic Noise Reduces Male Reproductive Investment in an Acoustically Signaling Insect. Behav. Ecol. Sociobiol. 2020, 74, 103. [Google Scholar] [CrossRef]
- Dinakaran, S.; Anbalagan, S. Anthropogenic Impacts on Aquatic Insects in Six Streams of South Western Ghats. J. Insect Sci. 2007, 7. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Code | Species | Collecting Site | GenBank Accession Number | ||
|---|---|---|---|---|---|
| 12S | 16S | CMG * | |||
| Samples from This Study | |||||
| CPCG001INA | C. gestroi | Cipinang, Indonesia | MW765243 | MW765223 | |
| GSCG001INA | C. gestroi | Gunung Sindur, Indonesia | MW765244 | MW765224 | |
| KRCG001INA | C. gestroi | Karadenan, Indonesia | MW765246 | MW765226 | |
| PPCG001INA | C. gestroi | Parung Panjang, Indonesia | MW765247 | MW765227 | |
| CICG001INA | C. gestroi | Cibinong, Indonesia | MW765248 | MW765228 | |
| PKCG002INA | C. gestroi | Pondok Kelapa, Indonesia | MW765250 | MW765230 | |
| BTCG001INA | C. gestroi | Batam, Indonesia Col. 1 | MW765245 | MW765225 | |
| BTCG002INA | C. gestroi | Batam, Indonesia Col. 2 | MW765249 | MW765229 | |
| BTCS001INA | C. sepangensis | Batam, Indonesia | MW765251 | MW765231 | |
| SICS001INA | C. sepangensis | Alafan Simeulue, Indonesia | MW765252 | MW765232 | |
| PPCK001INA | C. kalshoveni | Parung Panjang, Indonesia | MW765254 | MW765234 | |
| PPCC001INA | C. curvignathus | Parung Panjang, Indonesia | MW765255 | MW765235 | |
| SICC001INA | C. curvignathus | East Simeulue, Indonesia | MW765256 | MW765236 | |
| KBCC001INA | C. curvignathus | Karimunbesar, Indonesia | MW765257 | MW765237 | |
| ASCC003INA | C. elisae | Asmat, Papua, Indonesia | MW765260 | MW765240 | |
| WYCF001JP | C. formosanus | Wakayama, Japan | MW765261 | MW765241 | |
| CIOJ001INA | Odontotermes sp. | Cibinong, Indonesia | MW765262 | MW765242 | |
| Samples from other studies | |||||
| C. gestroi | Singapore | KU925205 | |||
| C. heimi | Pakistan | KU925206 | |||
| C. heimi | Pakistan | KU925208 | |||
| C. sepangensis | Brunei | KU925215 | |||
| C. kalshoveni | Singapore | KU925209 | |||
| C. kalshoveni | Thailand | KU925210 | |||
| C. formosanus | Okinawa Island, Okinawa | AB626145 | |||
| C. formosanus | Iriomote Island, Okinawa | AB626147 | |||
| H. malabaricus | India | KU925227 | |||
| H. validus | Australia | KU925235 | |||
| P. canalifrons | Réunion Island, France | KP026256 | |||
| R. leptomandibularis | Zhejiang, China | MK41931 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wikantyoso, B.; Tseng, S.-P.; Himmi, S.K.; Yusuf, S.; Yoshimura, T. Morphometric Analysis of Coptotermes spp. Soldier Caste (Blattodea: Rhinotermitidae) in Indonesia and Evidence of Coptotermes gestroi Extreme Head-Capsule Shapes. Insects 2021, 12, 477. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050477
Wikantyoso B, Tseng S-P, Himmi SK, Yusuf S, Yoshimura T. Morphometric Analysis of Coptotermes spp. Soldier Caste (Blattodea: Rhinotermitidae) in Indonesia and Evidence of Coptotermes gestroi Extreme Head-Capsule Shapes. Insects. 2021; 12(5):477. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050477
Chicago/Turabian StyleWikantyoso, Bramantyo, Shu-Ping Tseng, Setiawan Khoirul Himmi, Sulaeman Yusuf, and Tsuyoshi Yoshimura. 2021. "Morphometric Analysis of Coptotermes spp. Soldier Caste (Blattodea: Rhinotermitidae) in Indonesia and Evidence of Coptotermes gestroi Extreme Head-Capsule Shapes" Insects 12, no. 5: 477. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12050477