
Short-Term Selection to Diflubenzuron and Bacillus thuringiensis Var. Israelensis Differentially Affects the Winter Survival of Culex pipiens f. Pipiens and Culex pipiens f. Molestus (Diptera: Culicidae)


 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mosquito Colonies and Rearing Methods
2.2. Larval Bioassays
2.3. Larval Selection
2.4. Winter Survival
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farajollahi, A.; Fonseca, D.M.; Kramer, L.D.; Kilpatrick, A.M. “Bird biting” mosquitoes and human disease: A review of the role of Culex pipiens complex mosquitoes in epidemiology. Infect. Genet. Evol. 2011, 11, 1577–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugman, V.A.; Hernández-Triana, L.M.; Medlock, J.M.; Fooks, A.R.; Carpenter, S.; Johnson, N. The Role of Culex pipiens L. (Diptera: Culicidae) in Virus Transmission in Europe. Int. J. Environ. Res. Public Health 2018, 15, 389. [Google Scholar] [CrossRef] [Green Version]
- Vinogradova, E.B. Culex Pipiens Pipiens Mosquitoes: Taxonomy, Distribution, Ecology, Physiology, Genetics, Applied Importance and Control. Pensoft Publishers: Sofia, Bulgaria; Moscow, Russia, 2000; ISBN 954-642-103-0. [Google Scholar]
- Becker, N.; Jöst, A.; Weitzel, T. The Culex pipiens Complex in Europe. J. Am. Mosq. Control. Assoc. 2012, 28, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Vogels, C.B.; Van De Peppel, L.J.; Van Vliet, A.J.; Westenberg, M.; Ibañez-Justicia, A.; Stroo, A.; Buijs, J.A.; Visser, T.M.; Koenraadt, C.J. Winter Activity and Aboveground Hybridization Between the Two Biotypes of the West Nile Virus Vector Culex pipiens. Vector-Borne Zoonotic Dis. 2015, 15, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.J.; Briegel, H. Inability of diapausing Culex pipiens (Diptera: Culicidae) to use blood for producing lipid reserves for overwitrer survival. J. Med. Entomol. 1989, 26, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Sim, C.; Denlinger, D.L. Insulin signaling and the regulation of insect diapause. Front. Physiol. 2013, 4, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robich, R.M.; Denlinger, D.L. Diapause in the mosquito Culex pipiens evokes a metabolic switch from blood feeding to sugar gluttony. Proc. Natl. Acad. Sci. USA 2005, 102, 15912–15917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, H.; Neill, S.L.O. Controlling vector-borne diseases by releasing modified mosquitoes. Nat. Rev. Microbiol. 2018, 16, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Bellini, R.; Zeller, H.; Van Bortel, W. A review of the vector management methods to prevent and control outbreaks of West Nile virus infection and the challenge for Europe. Parasites Vectors 2014, 7, 323. [Google Scholar] [CrossRef] [Green Version]
- Merzendorfer, H. Chitin synthesis inhibitors: Old molecules and new developments. Insect Sci. 2012, 20, 121–138. [Google Scholar] [CrossRef]
- Ben-Dov, E. Bacillus thuringiensis subsp. israelensis and Its Dipteran-Specific Toxins. Toxins 2014, 6, 1222–1243. [Google Scholar] [CrossRef]
- Cantón, P.E.; Reyes, E.Z.; De Escudero, I.R.; Bravo, A.; Soberón, M. Binding of Bacillus thuringiensis subsp. israelensis Cry4Ba to Cyt1Aa has an important role in synergism. Peptides 2011, 32, 595–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, C.; Fernandez, L.E.; Sun, J.; Folch, J.L.; Gill, S.S.; Soberón, M.; Bravo, A. Bacillus thuringiensis subsp. israelensis Cyt1Aa synergizes Cry11Aa toxin by functioning as a membrane-bound receptor. Proc. Natl. Acad. Sci. USA 2005, 102, 18303–18308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirth, M.C.; Park, H.-W.; Walton, W.E.; Federici, B.A. Cyt1A of Bacillus thuringiensis Delays Evolution of Resistance to Cry11A in the Mosquito Culex quinquefasciatus. Appl. Environ. Microbiol. 2005, 71, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Grigoraki, L.; Puggioli, A.; Mavridis, K.; Douris, V.; Montanari, M.; Bellini, R.; Vontas, J. Striking diflubenzuron resistance in Culex pipiens, the prime vector of West Nile Virus. Sci. Rep. 2017, 7, 11699. [Google Scholar] [CrossRef] [Green Version]
- Porretta, D.; Fotakis, E.; Mastrantonio, V.; Chaskopoulou, A.; Michaelakis, A.; Kioulos, I.; Weill, M.; Urbanelli, S.; Vontas, J.; Bellini, R. Focal distribution of diflubenzuron resistance mutations in Culex pipiens mosquitoes from Northern Italy. Acta Trop. 2019, 193, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Fotakis, E.A.; Mastrantonio, V.; Grigoraki, L.; Porretta, D.; Puggioli, A.; Chaskopoulou, A.; Osório, H.; Weill, M.; Bellini, R.; Urbanelli, S.; et al. Identification and detection of a novel point mutation in the Chitin Synthase gene of Culex pipiens associated with diflubenzuron resistance. PLoS Negl. Trop. Dis. 2020, 14, e0008284. [Google Scholar] [CrossRef] [PubMed]
- Guz, N.; Cagatay, N.S.; Fotakis, E.A.; Durmusoglu, E.; Vontas, J. Detection of diflubenzuron and pyrethroid resistance mutations in Culex pipiens from Mugla, Turkey. Acta Trop. 2020, 203, 105294. [Google Scholar] [CrossRef]
- Paul, A.; Harrington, L.C.; Zhang, L.; Scott, J.G. Insecticide resistance in Culex pipiens from New York. J. Am. Mosq. Control. Assoc. 2005, 21, 305–309. [Google Scholar] [CrossRef]
- Ferreira, L.M.; Silva-Filha, M.H.L.N. Bacterial larvicides for vector control: Mode of action of toxins and implications for resistance. Biocontrol. Sci. Technol. 2013, 23, 1137–1168. [Google Scholar] [CrossRef]
- Liu, N. Insecticide Resistance in Mosquitoes: Impact, Mechanisms, and Research Directions. Annu. Rev. Ѐntomol. 2015, 60, 537–559. [Google Scholar] [CrossRef]
- Kliot, A.; Ghanim, M. Fitness costs associated with insecticide resistance. Pest Manag. Sci. 2012, 68, 1431–1437. [Google Scholar] [CrossRef] [PubMed]
- Gazave, É.; Chevillon, C.; Lenormand, T.; Marquine, M.; Raymond, M. Dissecting the cost of insecticide resistance genes during the overwintering period of the mosquito Culex pipiens. Heredity 2001, 87, 441–448. [Google Scholar] [CrossRef] [Green Version]
- Peppe, P. Molecular Identification of Species of Mosquitoes for the Detection of Vectors of the West Nile Virus. Master’s Thesis, University of Thessaly, Volos, Greece, 2015. (Abstract in English). [Google Scholar]
- West Nile Virus in Greece. Annual Epidemiological Report for 2019. National Public Health Organization. H.C.D.C.P. 2019. Available online: https://eody.gov.gr/wp-content/uploads/2019/01/WNV_annual_report_2019_ENG.pdf (accessed on 8 April 2021).
- Kioulos, I.; Kampouraki, A.; Morou, E.; Skavdis, G.; Vontas, J. Insecticide resistance status in the major West Nile virus vector Culex pipiens from Greece. Pest Manag. Sci. 2014, 70, 623–627. [Google Scholar] [CrossRef]
- Dieme, C.; Yssouf, A.; Vega-Rúa, A.; Berenger, J.-M.; Failloux, A.-B.; Raoult, D.; Parola, P.; Almeras, L. Accurate identification of Culicidae at aquatic developmental stages by MALDI-TOF MS profiling. Parasites Vectors 2014, 7, 544. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for Laboratory and Field Testing of Mosquito Larvicides; World Health Organisation: Geneva, Switzerland, 2005. [Google Scholar]
- Finney, D.J. Probit Analysis; Cambridge University Press: London, UK, 1971. [Google Scholar]
- Spielman, A.; Wong, J. Environmental Control of Ovarian Diapause in Culex pipiens. Ann. Entomol. Soc. Am. 1973, 66, 905–907. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Yang, C.; Huang, J.Y.; Lü, L. Susceptibility of field populations of Anopheles sinensis (Diptera: Culicidae) to Bacillus thuringiensis subsp israelensis. Biocontrol. Sci. Technol. 2004, 14, 321–325. [Google Scholar]
- Boyer, S.; Tilquin, M.; Ravanel, P. Differential sensitivity to Bacillus thuringiensis var. israelensis and temephos in field mosquito populations of Ochlerotatus cataphylla (Diptera: Culicidae): Toward resistance? Environ. Toxicol. Chem. 2007, 26, 157–162. [Google Scholar] [CrossRef]
- Boyer, S.; Paris, M.; Jego, S.; Lempérière, G.; Ravanel, P. Influence of insecticide Bacillus thuringiensis subsp. israelensis treatments on resistance and enzyme activities in Aedes rusticus larvae (Diptera: Culicidae). Biol. Control. 2012, 62, 75–81. [Google Scholar] [CrossRef]
- Georghiou, G.P.; Wirth, M.C. Influence of exposure to single versus multiple toxins of Bacillus thuringiensis subsp. israelensis on development of resistance in the mosquito Culex quinquefasciatus (Diptera: Culicidae). Appl. Environ. Microbiol. 1997, 63, 1095–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, M.S.; Kelada, N.L.; Zahran, H.M.; El-Meniawi, F.A. Resistance development in mosquito larvae Culex pipiens to the bacterial agent Bacillus thuringiensis var. israelensis. J. Appl. Ѐntomol. 2003, 127, 29–32. [Google Scholar] [CrossRef]
- da Silva Carvalho, K.; Crespo, M.M.; Araújo, A.P.; da Silva, R.S.; de Melo-Santos, M.A.V.; de Oliveira, C.M.F.; Silva-Filha, M.H.N.L. Long-term exposure of Aedes aegypti to Bacillus thuringiensis svar. israelensis did not involve altered susceptibility to this microbial larvicide or to other control agents. Parasites Vectors 2018, 11, 673. [Google Scholar] [CrossRef] [PubMed]
- Paris, M.; David, J.-P.; Despres, L. Fitness costs of resistance to Bti toxins in the dengue vector Aedes aegypti. Ecotoxicology 2011, 20, 1184–1194. [Google Scholar] [CrossRef]
- Becker, N. Microbial control of mosquitoes: Management of the upper rhine mosquito population as a model programme. Parasitol. Today 1997, 13, 485–487. [Google Scholar] [CrossRef]
- Mittal, P.K.; Adak, T.; Subbarao, S.K. Laboratory selection to investigate the development of resistance to Bacillus thuringiensis var. israelensis H-14 in Culex quinquefasciatus Say (Diptera: Culicidae). Natl. Acad. Sci. Lett. 2005, 28, 281–283. [Google Scholar]
- Becker, N.; Ludwig, M.; Su, T. Lack of Resistance in Aedes vexans Field Populations After 36 Years of Bacillus thuringiensis subsp. israelensis Applications in the Upper Rhine Valley, Germany. J. Am. Mosq. Control. Assoc. 2018, 34, 154–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadavid-Restrepo, G.; Sahaza, J.; Orduz, S. Treatment of an Aedes aegypti colony with the Cry11Aa toxin for 54 generations results in the development of resistance. Memórias Inst. Oswaldo Cruz 2012, 107, 74–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stalinski, R.; Tetreau, G.; Gaude, T.; Després, L. Pre-selecting resistance against individual Bti Cry toxins facilitates the development of resistance to the Bti toxins cocktail. J. Invertebr. Pathol. 2014, 119, 50–53. [Google Scholar] [CrossRef]
- Spanoudis, C.G.; Andreadis, S.S.; Tsaknis, N.K.; Petrou, A.P.; Gkeka, C.D.; Savopoulou–Soultani, M. Effect of Temperature on Biological Parameters of the West Nile Virus Vector Culex pipiens form ‘molestus’ (Diptera: Culicidae) in Greece: Constant vs Fluctuating Temperatures. J. Med. Ѐntomol. 2018, 56, 641–650. [Google Scholar] [CrossRef]
- Bourguet, D.; Guillemaud, T.; Chevillon, C.; Raymond, M. Fitness costs of insecticide resistance in natural breeding sites of the mosquito Culex pipiens. Evolution 2004, 58, 128–135. [Google Scholar] [CrossRef]
- Tetreau, G.; Grizard, S.; Patil, C.D.; Tran, F.-H.; Van, V.T.; Stalinski, R.; Laporte, F.; Mavingui, P.; Després, L.; Moro, C.V. Bacterial microbiota of Aedes aegypti mosquito larvae is altered by intoxication with Bacillus thuringiensis israelensis. Parasites Vectors 2018, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takken, W.; Smallegange, R.C.; Vigneau, A.J.; Johnston, V.; Brown, M.; Mordue-Luntz, A.J.; Billingsley, P.F. Larval nutrition differentially affects adult fitness and Plasmodium development in the malaria vectors Anopheles gambiae and Anopheles stephensi. Parasites Vectors 2013, 6, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivero, A.; Magaud, A.; Nicot, A.; Vézilier, J. Energetic cost of insecticide resistance in Culex pipiens mosquitoes. J. Med. Ѐntomol. 2011, 48, 694–700. [Google Scholar] [CrossRef]
- Fotakis, E.A.; Chaskopoulou, A.; Grigoraki, L.; Tsiamantas, A.; Kounadi, S.; Georgiou, L.; Vontas, J. Analysis of population structure and insecticide resistance in mosquitoes of the genus Culex, Anopheles and Aedes from different environments of Greece with a history of mosquito borne disease transmission. Acta Trop. 2017, 174, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Koenraadt, C.J.M.; Möhlmann, T.W.R.; Verhulst, N.O.; Spitzen, J.; Vogels, C.B.F. Effect of overwintering on survival and vector competence of the West Nile virus vector Culex pipiens. Parasites Vectors 2019, 12, 147. [Google Scholar] [CrossRef]
- Rinehart, J.P.; Yocum, G.D.; Leopold, R.A.; Robich, R.M. Cold storage of Culex pipiens in the absence of diapause. J. Med. Ѐntomol. 2010, 47, 1071–1076. [Google Scholar] [CrossRef] [Green Version]
- Faraji, A.; Gaugler, R. Experimental host preference of diapause and non-diapause induced Culex pipiens pipiens (Diptera: Culicidae). Parasites Vectors 2015, 8, 389. [Google Scholar] [CrossRef] [Green Version]
- Belinato, T.A.; Valle, D. The Impact of Selection with Diflubenzuron, a Chitin Synthesis Inhibitor, on the Fitness of Two Brazilian Aedes aegypti Field Populations. PLoS ONE 2015, 10, e0130719. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, N.T.; Carey, J.R.; Ioannou, C.S.; Ji, H.; Muller, H.-G.; Wang, J.-L.; Luckhart, S.; Lewis, E.E. Seasonality of Post-capture Longevity in a Medically-Important Mosquito (Culex pipiens). Front. Ecol. Evol. 2016, 4, 63. [Google Scholar] [CrossRef] [Green Version]



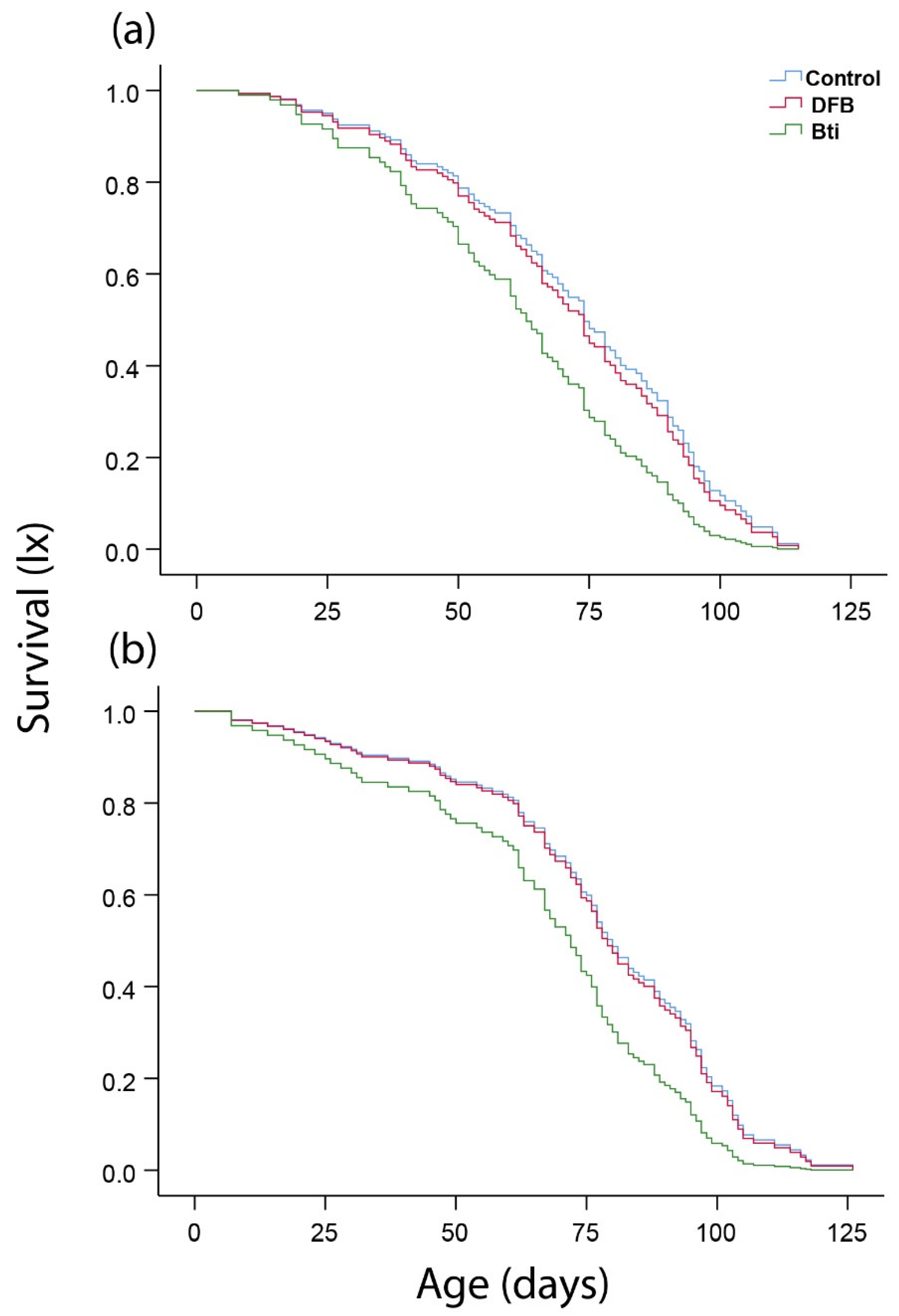{kind=link}
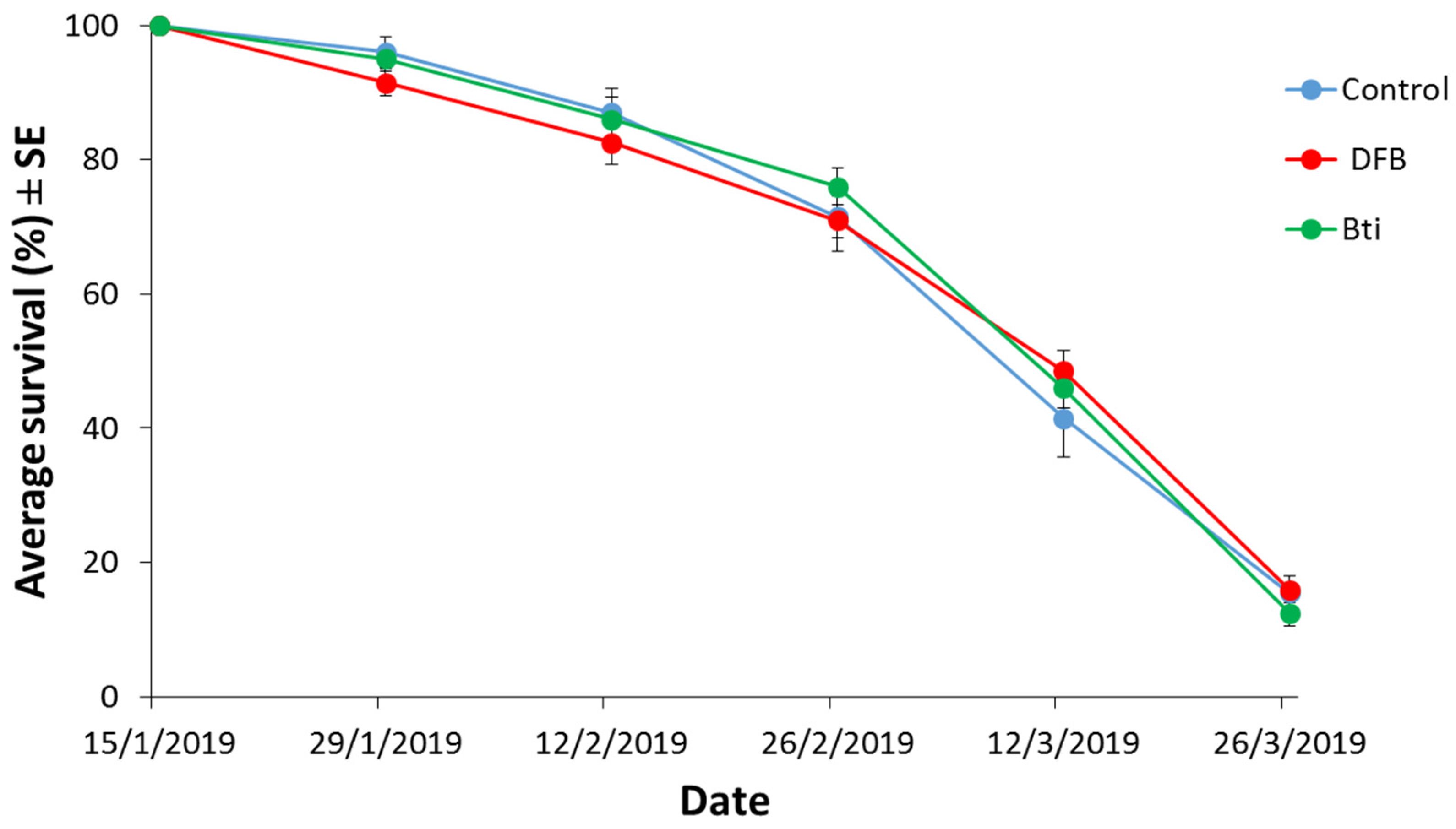{kind=link}
| Population | N * | EI50 (95% CL) a | RR50 | EI90 (95% CL) a | RR90 | Slope | X2 (df) | |
|---|---|---|---|---|---|---|---|---|
| Cx. pipiens f. pipiens | Control | 3150 | 0.0025 (0.0014–0.0035) | - | 0.0081 (0.0065–0.0097) | - | 2.47 | 177.57 b (105) |
| Selected | 2700 | 0.0093 (0.0076–0.0106) | 3.7 | 0.0252 (0.0218–0.0321) | 3.1 | 2.96 | 70.98 (87) | |
| Cx. pipiens f. molestus | Control | 3150 | 0.0022 (0.0016–0.0026) | - | 0.0040 (0.0037–0.0044) | - | 4.49 | 201.99 b (105) |
| Selected | 2700 | 0.0037 (0.0028–0.0046) | 1.7 | 0.0116 (0.0101–0.0132) | 2.9 | 2.60 | 57.94 (87) | |
| Population | N * | LC50 (95% CL) a | RR50 | LC90 (95% CL) a | RR90 | Slope | X2 (df) | |
|---|---|---|---|---|---|---|---|---|
| Cx. pipiens f. pipiens | Control | 2700 | 0.031 (0.029–0.033) | - | 0.047 (0.044–0.051) | - | 7.52 | 120.73 b (87) |
| Selected | 2700 | 0.042 (0.035–0.048) | 1.3 | 0.069 (0.065–0.074) | 1.5 | 6.12 | 47.99 (87) | |
| Cx. pipiens f. molestus | Control | 2700 | 0.032 (0.029–0.034) | - | 0.047 (0.044–0.051) | - | 7.62 | 126.68 b (87) |
| Selected | 2700 | 0.045 (0.034–0.059) | 1.4 | 0.081 (0.065–0.089) | 1.7 | 4.98 | 117.21 b (87) | |
| Population | Larval Survival * a (% ± SE) | Larval Developmental Duration b (Days ± SE) | Pupal Survival c (%) | Pupal Developmental Duration b (Days ± SE) | |
|---|---|---|---|---|---|
| Males | Females | ||||
| Control | 35.1 ± 1.77 a | 48.54 ± 0.17 c (351) | 50.7 a | 11.75 ± 0.09 a (85) | 11.89 ± 0.08 a (93) |
| F3 DFB | 16.0 ± 1.58 c | 49.22 ± 0.20 b (160) | 45.0 a | 11.50 ± 0.10 a (30) | 11.64 ± 0.11 a (40) |
| F3 Bti | 23.2 ± 2.31 b | 50.49 ± 0.32 a (232) | 44.4 a | 11.57 ± 0.10 a (59) | 11.87 ± 0.11 a (47) |
| F | 25.26 | - | - | - | - |
| X2 | - | 30.26 | 2.10 | 3.38 | 1.42 |
| df | 2, 12 | 2 | 2 | 2 | 2 |
| p | <0.001 | <0.001 | 0.272 | 0.184 | 0.490 |
| Longevity Parameters in Days ± SE | ||||
|---|---|---|---|---|
| Average | Quartiles | |||
| 25 | 50 | 75 | ||
| Males | ||||
| Control (n = 49) | 72.88 ± 3.46 a | 92 ± 3.51 | 75 ± 6.29 | 60 ± 6.85 |
| F3 DFB (n = 30) | 69.13 ± 4.48 ab | 93 ± 6.95 | 66 ± 1.36 | 50 ± 9.68 |
| F3 Bti (n = 47) | 61.23 ± 3.69 b | 80 ± 4.83 | 68 ± 5.38 | 46 ± 11.45 |
| Females | ||||
| Control (n = 49) | 73.38 ± 4.37 a | 97 ± 1.72 | 77 ± 2.78 | 62 ± 13.39 |
| F3 DFB (n = 30) | 77.23 ± 4.50 a | 95 ± 6.17 | 76 ± 5.46 | 62 ± 3.22 |
| F3 Bti (n = 47) | 70.08 ± 3.33 b | 88 ± 3.59 | 77 ± 2.56 | 62 ± 7.44 |
| Population | N * | Blood Meal Acceptance a (%) | Preoviposition Period b (Days ± SE) | Eggs Per Raft c (±SE) | Larval Hatch Rate Per Egg Raft c (% ± SE) |
|---|---|---|---|---|---|
| Control | 31 | 35.71 a | 10.00 ± 0.76 a | 61.10 ± 5.43 a | 89.10 ± 3.42 a |
| F3 DFB | 32 | 34.38 a | 11.72 ± 0.61 a | 51.54 ± 4.50 a | 92.58 ± 3.04 a |
| F3 Bti | 25 | 46.43 a | 11.53 ± 0.50 a | 51.46 ± 3.70 a | 90.68 ± 3.81 a |
| F | - | - | - | 1.43 | 0.22 |
| X2 | - | 1.06 | 2.64 | - | - |
| df | - | 2 | 2 | 2, 31 | 2, 31 |
| p | - | 0.587 | 0.266 | 0.254 | 0.798 |
| Longevity Parameters in Days ± SE | ||||
|---|---|---|---|---|
| Population | Average | Quartiles | ||
| 25 | 50 | 75 | ||
| Control (n = 31) | 81.45 ± 6.02 a | 109 ± 6.59 | 89 ± 5.00 | 68 ± 15.02 |
| F3 DFB (n = 32) | 92.40 ± 5.84 a | 115 ± 1.23 | 105 ± 2.80 | 83 ± 22.04 |
| F3 Bti (n = 25) | 99.84 ± 3.19 a | 115 ± 1.62 | 97 ± 1.48 | 92 ± 2.91 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ioannou, C.S.; Hadjichristodoulou, C.; Kyritsi, M.A.; Papadopoulos, N.T. Short-Term Selection to Diflubenzuron and Bacillus thuringiensis Var. Israelensis Differentially Affects the Winter Survival of Culex pipiens f. Pipiens and Culex pipiens f. Molestus (Diptera: Culicidae). Insects 2021, 12, 527. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12060527
Ioannou CS, Hadjichristodoulou C, Kyritsi MA, Papadopoulos NT. Short-Term Selection to Diflubenzuron and Bacillus thuringiensis Var. Israelensis Differentially Affects the Winter Survival of Culex pipiens f. Pipiens and Culex pipiens f. Molestus (Diptera: Culicidae). Insects. 2021; 12(6):527. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12060527
Chicago/Turabian StyleIoannou, Charalampos S., Christos Hadjichristodoulou, Maria A. Kyritsi, and Nikos T. Papadopoulos. 2021. "Short-Term Selection to Diflubenzuron and Bacillus thuringiensis Var. Israelensis Differentially Affects the Winter Survival of Culex pipiens f. Pipiens and Culex pipiens f. Molestus (Diptera: Culicidae)" Insects 12, no. 6: 527. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12060527