
Nutritional Composition of Honey Bee Drones of Two Subspecies Relative to Their Pupal Developmental Stages


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Identification of the Species
2.3. Amino Acid Analysis
2.4. Fatty Acid Analysis
2.5. Mineral Analysis
2.6. Statistical Analysis
3. Results
3.1. Identification of Specimens
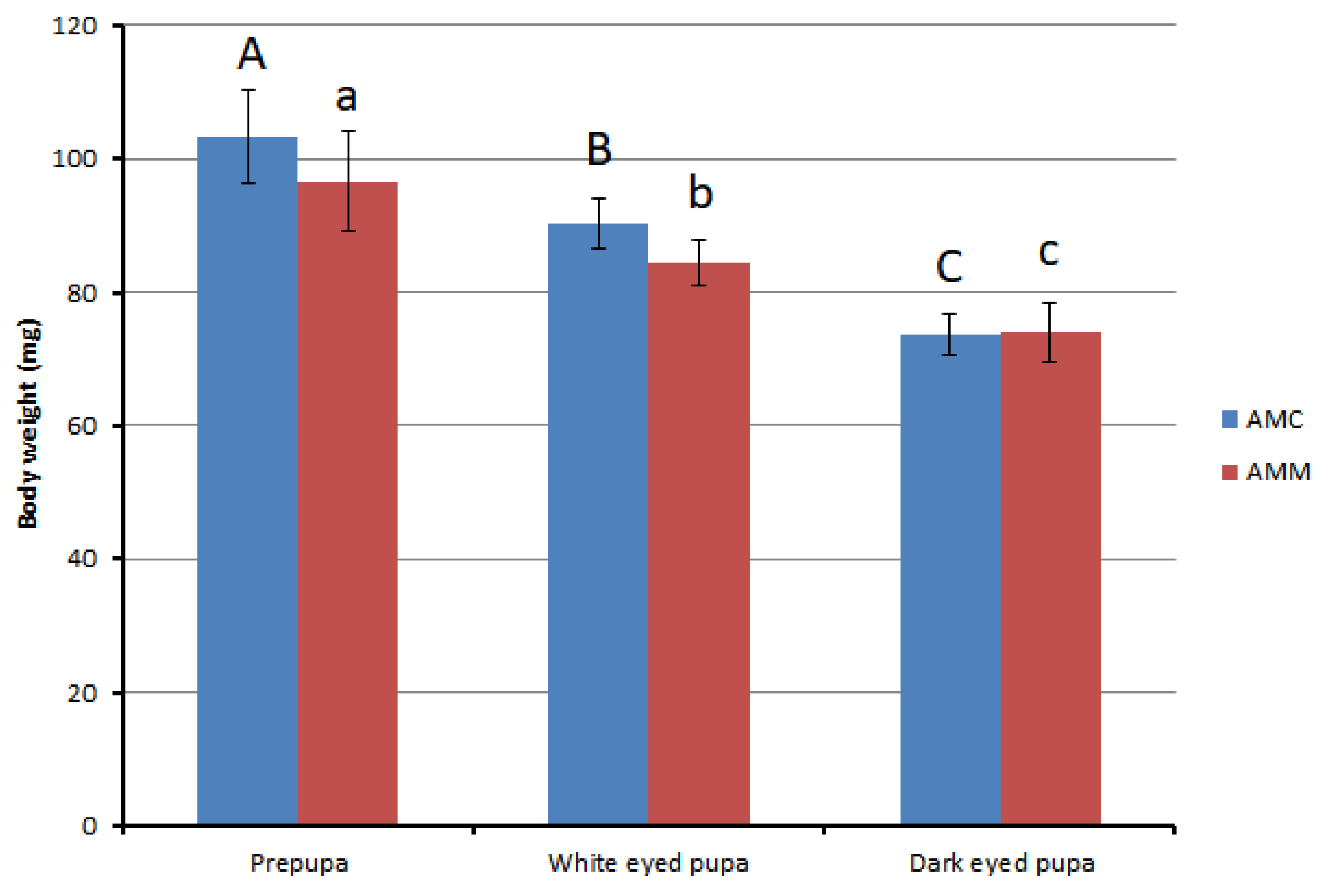
3.2. Body Weights and Sizes
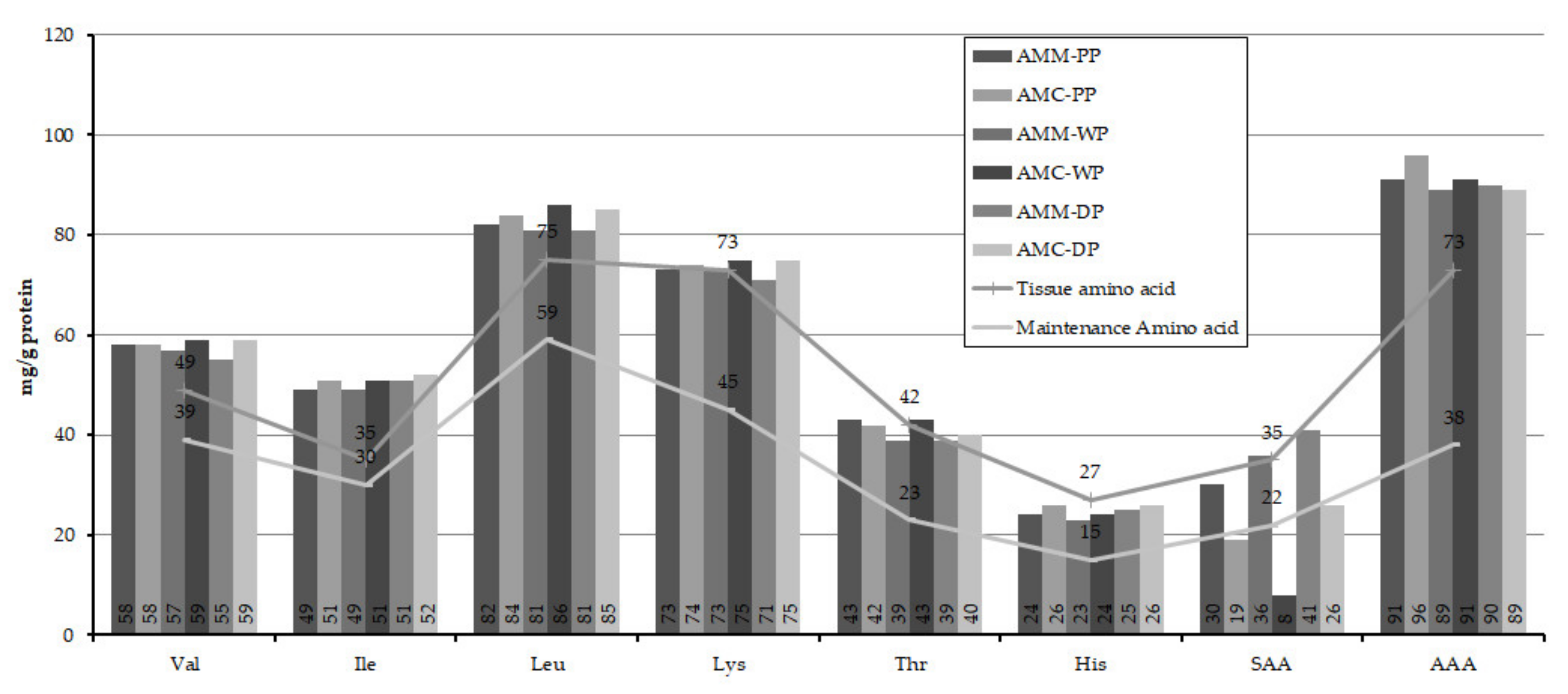
3.3. Amino Acid Composition
3.4. Fatty Acid Composition
3.5. Mineral Content
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- D’Errico, F.; Backwell, L.; Villa, P.; Degano, I.; Lucejko, J.J.; Bamford, M.K.; Higham, T.; Colombini, M.P.; Beaumont, P.B. Early evidence of San material culture represented by organic artifacts from Border Cave, South Africa. Proc. Natl. Acad. Sci. USA 2012, 109, 13214–13219. [Google Scholar] [CrossRef] [Green Version]
- Roffet-Salque, M.; Regert, M.; Evershed, R.P.; Outram, A.K.; Cramp, L.J.E.; Decavallas, O.; Dunne, J.; Gerbault, P.; Mileto, S.; Mirabaud, S.; et al. Widespread exploitation of the honeybee by early Neolithic farmers. Nature 2016, 534, S17–S18. [Google Scholar] [CrossRef] [Green Version]
- Pager, H. Cave Paintings Suggest Honey Hunting Activities in Ice Age Times. Bee World 1976, 57, 9–14. [Google Scholar] [CrossRef]
- Mazar, A.; Panitz-Cohen, N. It is the land of honey: Beekeeping at Tel Hehov. Near East. Archaeol. 2007, 40, 202–219. [Google Scholar] [CrossRef]
- Ediriweera, E.R.H.S.S.; Premarathna, N.Y.S. Medicinal and cosmetic uses of bee’s honey—A review. Ayurveda 2021, 33, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Stangaciu, S. Apitherapy: Holistic healing through the honeybee and bee products in countries with poor healthcare system. In Beekeeping for Poverty Alleviation and Livelihood Security; Gupta, R.K., Reybroek, W., Van Veen, J.W., Gupta, A., Eds.; Springer: Berlin, Germany, 2014; pp. 413–447. [Google Scholar]
- Meyer-Rochow, V.B. Therapeutic arthropods and other, largely terrestrial, folk-medicinally important invertebrates: A comparative survey and review. J. Ethnobiol. Ethnomed. 2017, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherry, R.H. Use of Insects by Australian Aborigines. Am. Èntomol 1991, 37, 8–13. [Google Scholar] [CrossRef]
- Mbata, K.J. Traditional use of arthropods in Zambia. I. The food insects. Food Insects Newslett. 1995, 8, 5–7. [Google Scholar]
- Murray, S.S.; Schoeninger, M.J.; Bunn, H.T.; Pickering, T.R.; Marlett, J.A. Nutritional Composition of Some Wild Plant Foods and Honey Used by Hadza Foragers of Tanzania. J. Food Compos. Anal 2001, 14, 3–13. [Google Scholar] [CrossRef]
- Wongsiri, S.; Lekprayoon, C.; Thapa, R.; Thirakupt, K.; Rinderer, E.T.; Sylvester, A.H.; Oldroyd, B.P.; Booncham, U. Comparative biology of Apis andreniformis and Apis floreain Thailand. Bee World 1997, 78, 23–35. [Google Scholar] [CrossRef]
- Ramos-Elorduy, J.; Moreno, J.M.P.; Prado, E.E.; Perez, M.A.; Otero, J.L.; de Guevara, O.L. Nutritional Value of Edible Insects from the State of Oaxaca, Mexico. J. Food Compos. Anal. 1997, 10, 142–157. [Google Scholar] [CrossRef]
- Onore, G. A brief note on edible insects in Ecuador. Ecol. Food Nutr. 1997, 36, 277–285. [Google Scholar] [CrossRef]
- Bukkens, S.G.F. The nutrient value of edible insects. Ecol. Food Nutr. 1997, 36, 287–319. [Google Scholar] [CrossRef]
- Banjo, A.D.; Songonuga, E.A. The nutritional value of fourteen species of edible insects in southwestern Nigeria. Afr. J. Bio-technol. 2006, 5, 298–301. [Google Scholar]
- Rumpold, B.A.; Schlüter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef]
- Chakravorty, J.; Ghosh, S.; Megu, K.; Jung, C.; Meyer-Rochow, V.B. Nutritional and anti-nutritional composition of Oecophylla smaragdina (Hymenoptera: Formicidae) and Odontotermes sp. (Isoptera: Termitidae): Two preferred edible insects of Arunachal Pradesh, India. J. Asia-Pac. Èntomol. 2016, 19, 711–720. [Google Scholar] [CrossRef]
- Ghosh, S.; Lee, S.-M.; Jung, C.; Meyer-Rochow, V. Nutritional composition of five commercial edible insects in South Korea. J. Asia-Pac. Èntomol 2017, 20, 686–694. [Google Scholar] [CrossRef]
- Kulma, M.; Kouřimská, L.; Homolková, D.; Božik, M.; Plachý, V.; Vrabec, V. Effect of developmental stage on the nutritional value of edible insects. A case study with Blaberus craniifer and Zophobas morio. J. Food Compos. Anal. 2020, 92, 103570. [Google Scholar] [CrossRef]
- Kouřimská, L.; Adámková, A. Nutritional and sensory quality of edible insects. NFS J. 2016, 4, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Payne, C.; Scarborough, P.; Rayner, M.; Nonaka, K. A systematic review of nutrient composition data available for twelve commercially available edible insects, and comparison with reference values. Trends Food Sci. Technol. 2016, 47, 69–77. [Google Scholar] [CrossRef]
- Nowak, V.; Persijn, D.; Rittenschober, D.; Charrondiere, U.R. Review of food composition data for edible insects. Food Chem. 2016, 193, 39–46. [Google Scholar] [CrossRef]
- Ghosh, S.; Namin, S.M.; Meyer-Rochow, V.B.; Jung, C. Chemical composition and nutritional value of different species of Vespa hornets. Foods 2021, 10, 418. [Google Scholar] [CrossRef]
- Longvah, T.; Mangthya, K.; Ramulu, P. Nutrient composition and protein quality evaluation of eri silkworm (Samiaricini) prepupae and pupae. Food Chem. 2011, 128, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Finke, M.D. Nutrient Composition of Bee Brood and its Potential as Human Food. Ecol. Food Nutr. 2005, 44, 257–270. [Google Scholar] [CrossRef]
- Ghosh, S.; Jung, C.; Meyer-Rochow, V.B. Nutritional value and chemical composition of larvae, pupae, and adults of worker honey bee, Apis mellifera ligustica as a sustainable food source. J. Asia-Pac. Èntomol. 2016, 19, 487–495. [Google Scholar] [CrossRef]
- Ghosh, S.; Chuttong, B.; Burgett, M.; Meyer-Rochow, V.B. Nutritional value of brood and adult workers of the Asia honeybee species Apis cerana and Apis dorsata. In African Edible Insects as Alternative Source of Food, Oil, Protein and Bioactive Compo-Nents; Mariod, A.A., Ed.; Springer Nature: Cham, Switzerland, 2020; pp. 265–273. [Google Scholar]
- Filipiak, M.; Woyciechowski, M.; Czarnoleski, M. Stoichiometric niche, nutrient partitioning and resource allocation in a solitary bee are sex-specific and phosphorous is allocated mainly to the cocoon. Sci. Rep. 2021, 11, 652. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.B.; Evans, J.; Jonas-Levi, A.; Benjamin, O.; Martinez, I.; Dahle, B.; Roos, N.; Lecocq, A.; Foley, K. Standard methods for Apis mellifera brood for human food. J. Apic. Res. 2019, 58, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c ox-idase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Hebert, P.D.; Ratnasingham, S.; De Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Boil. Sci 2003, 270, S96–S99. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis of the AOAC, 15th ed; Association of Official Analytical Chemists: Washington, DC, USA, 1990. [Google Scholar]
- Ministry of Food and Drug Safety. Korean Food Standard Codex; Ministry of Food and Drug Safety: Cheongju, Korea, 2010.
- FAO (Food and Agriculture Organization of the United Nations). Dietary Protein Quality Evaluation in Human Nutrition; FAO: Rome, Italy, 2013. [Google Scholar]
- Linus Pauling Institute, Micronutrient Information Center, Oregon State University. Available online: http://www.lpi.oregonstate.edu/mic (accessed on 15 February 2021).
- USDA Database on Nutrient Content of Food. Available online: http://www.data.nal.usda.gov (accessed on 2 April 2021).
- Van Huis, A.; Van Itterbeeck, J.; Klunder, H.; Mertens, E.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects: Future Prospects for Food and Feed Ecurity; FAO of the United Nations: Rome, Italy, 2013. [Google Scholar]
- Kinyuru, J.N.; Konyole, S.O.; Roos, N.; Onyango, C.A.; Owino, V.O.; Owuor, B.O.; Estambale, B.B.; Friis, H.; Aagaard-Hansen, J.; Kenji, G.M. Nutrient composition of four species of winged termites consumed in western Kenya. J. Food Compos. Anal. 2013, 30, 120–124. [Google Scholar] [CrossRef]
- Mozhui, L.; Kakati, L.; Kiewhuo, P.; Changkija, S. Traditional Knowledge of the Utilization of Edible Insects in Nagaland, North-East India. Foods 2020, 9, 852. [Google Scholar] [CrossRef]
- Gahukar, R. Entomophagy and human food security. Int. J. Trop. Insect Sci. 2011, 31, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Menozzi, D.; Sogari, G.; Veneziani, M.; Simoni, E.; Mora, C. Eating novel foods: An application of the Theory of Planned Behaviour to predict the consumption of an insect-based product. Food Qual. Prefer. 2017, 59, 27–34. [Google Scholar] [CrossRef]
- Gahukar, R.T. Edible insects collected from forests for family livelihood and wellness of rural communities: A review. Glob. Food Secur. 2020, 25. [Google Scholar] [CrossRef]
- Paul, A.; Frederich, M.; Uyttenbroeck, R.; Hatt, S.; Malik, P.; Lebecque, S.; Hamaidia, M.; Miazek, K.; Goffin, D.; Willems, L.; et al. Grasshopper s a food resource? A review. Biotechnol. Agron. Soc. Environ. 2016, 20, 337–352. [Google Scholar]
- Meyer-Rochow, V.B. Can insects help to ease the problem of world food shortage? Search 1975, 6, 261–262. [Google Scholar]
- Hedenus, F.; Wirsenius, S.; Johansson, D.J.A. The importance of reduced meat and dairy consumption for meeting stringent climate change targets. Clim. Chang. 2014, 124, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Rojas-Downing, M.M.; Nejadhashemi, A.P.; Harrigan, T.; Woznicki, S. Climate change and livestock: Impacts, adaptation, and mitigation. Clim. Risk Manag. 2017, 16, 145–163. [Google Scholar] [CrossRef]
- Belluco, S.; Losasso, C.; Maggioletti, M.; Alonzi, C.C.; Paoletti, M.G.; Ricci, A. Edible Insects in a Food Safety and Nutritional Perspective: A Critical Review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 296–313. [Google Scholar] [CrossRef]
- Raheem, D.; Raposo, A.; Oluwole, O.B.; Nieuwland, M.; Saraiva, A.; Carrascosa, C. Entomophagy: Nutritional, ecological, safety and legislation aspects. Food Res. Int. 2019, 126, 108672. [Google Scholar] [CrossRef]
- Gorbunova, N.A.; Zakharov, A.N. Edible insects as a source of alternative protein. A review. Theory Pract. Meat Process. 2021, 6, 23–32. [Google Scholar] [CrossRef]
- Patel, S.; Suleria, H.A.R.; Rauf, A. Edible insects as innovative foods: Nutritional and functional assessments. Trends Food Sci. Technol. 2019, 86, 352–359. [Google Scholar] [CrossRef]
- Jongema, Y. List of Edible Insects of the World; Wageningen UR: Wageningen, The Netherlands, 2015; Available online: http://https://tinyurl.com/mestm6p (accessed on 22 June 2020).
- Ghosh, S.; Jung, C.; Meyer-Rochow, V.B. What governs selection and acceptance of edible insect species? In Edible Insects in Sustainable Food Systems; Halloran, A., Vantomme, P., Roos, N., Eds.; Springer: Cham, Switzerland, 2018; pp. 331–351. [Google Scholar]
- Wendin, K.M.; Nyberg, M. Factors influencing consumer perception and acceptability of insect-based foods. Curr. Opin. Food Sci. 2021, 40, 67–71. [Google Scholar] [CrossRef]
- Elhassan, M.; Wendin, K.; Olsson, V.; Langton, M. Quality Aspects of Insects as Food—Nutritional, Sensory, and Related Concepts. Foods 2019, 8, 95. [Google Scholar] [CrossRef] [Green Version]
- Melgar-Lalanne, G.; Hernández-Álvarez, A.; Salinas-Castro, A. Edible Insects Processing: Traditional and Innovative Technologies. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1166–1191. [Google Scholar] [CrossRef] [Green Version]
- Megido, R.C.; Gierts, C.; Blecker, C.; Brostaux, Y.; Haubruge, É.; Alabi, T.; Francis, F. Consumer acceptance of insect-based alternative meat products in Western countries. Food Qual. Prefer. 2016, 52, 237–243. [Google Scholar] [CrossRef]
- Orkusz, A. Edible Insects versus Meat—Nutritional Comparison: Knowledge of Their Composition Is the Key to Good Health. Nutrients 2021, 13, 1207. [Google Scholar] [CrossRef] [PubMed]
- Nyberg, M.; Olsson, V.; Wendlin, K. Reasons for eating insects? Responses and reactions among Swedish consumers. Int. J. Gastron. Food Sci. 2020, 22, 10068. [Google Scholar] [CrossRef]
- Ghosh, S.; Jung, C.; Meyer-Rochow, V.; Dekebo, A. Perception of entomophagy by residents of Korea and Ethiopia revealed through structured questionnaire. J. Insects Food Feed. 2020, 6, 59–64. [Google Scholar] [CrossRef]
- Marcucci, C. Food frontiers: Insects as food, is the future already here? Mediterr. J. Nutr. Metab. 2020, 13, 43–52. [Google Scholar] [CrossRef]
- Klunder, H.; Wolkers-Rooijackers, J.; Korpela, J.; Nout, M. Microbiological aspects of processing and storage of edible insects. Food Control 2012, 26, 628–631. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B.; Jung, C. Insects Used as Food and Feed: Isn’t That What We All Need? Foods 2020, 9, 1003. [Google Scholar] [CrossRef]
- Ghosh, S.; Sohn, H.-Y.; Pyo, S.-J.; Jensen, A.B.; Meyer-Rochow, V.B.; Jung, C. Nutritional Composition of Apis mellifera Drones from Korea and Denmark as a Potential Sustainable Alternative Food Source: Comparison Between Developmental Stages. Foods 2020, 9, 389. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Jung, C.; Chuttong, B.; Burgett, M. Nutritional aspects of the dwarf honeybee (Apis florea L.) for human consumption. In The Future Role of Dwarf Honeybees in Natural and Agricultural Systems; Abrol, D.P., Ed.; CRC Press: Boca Raton, FL, USA, 2020; pp. 137–145. [Google Scholar]
- Vasal, S.K. The role of high lysine cereals in animal and human nutrition in Asia. In Protein Sources for the Animal Feed Industry: Expert Consultation and Workshop, Bangkok, 29 April–3 May 2002; FAO Animal Production and Health Proceedings; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004; pp. 167–183. [Google Scholar]
- Leinonen, I.; Iannetta, P.P.M.; Rees, R.; Russell, W.; Watson, C.; Barnes, A. Lysine Supply Is a Critical Factor in Achieving Sustainable Global Protein Economy. Front. Sustain. Food Syst. 2019, 3. [Google Scholar] [CrossRef]
- Melegh, B.; Pap, M.; Bock, I.; Rebouche, C.J. Relationship of Carnitine and Carnitine Precursors Lysine, ε-N-Trimethyllysine, and γ-Butyrobetaine in Drug-Induced Carnitine Depletion. Pediatr. Res. 1993, 34, 460–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, T.; Nakamura, T.; Shibakusa, T.; Sugita, M.; Naganuma, F.; Iida, T.; Miura, Y.; Mohsen, A.; Harada, R.; Yanai, K. Insufficient Intake of L-Histidine Reduces Brain Histamine and Causes Anxiety-Like Behaviors in Male Mice. J. Nutr. 2014, 144, 1637–1641. [Google Scholar] [CrossRef] [PubMed]
- Fernstrom, J.D.; Fernstrom, M.H. Tyrosine, Phenylalanine, and Catecholamine Synthesis and Function in the Brain. J. Nutr. 2007, 137, 1539 S–1547 S. [Google Scholar] [CrossRef] [PubMed]
- Petroff, O.A.C. Book Review: GABA and Glutamate in the Human Brain. Neuroscientist 2002, 8, 562–573. [Google Scholar] [CrossRef]
- Mlcek, J.; Rop, O.; Borkovcova, M.; Bednarova, M. A Comprehensive Look at the Possibilities of Edible Insects as Food in Europe –A Review. Pol. J. Food Nutr. Sci. 2014, 64, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Rochow, V.B.; Gahukar, R.; Ghosh, S.; Jung, C. Chemical Composition, Nutrient Quality and Acceptability of Edible Insects Are Affected by Species, Developmental Stage, Gender, Diet, and Processing Method. Foods 2021, 10, 1036. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect Fat Body: Energy, Metabolism, and Regulation. Annu. Rev. Èntomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [Green Version]
- Cripps, C.; Blomquist, G.J.; De Renobales, M. De novo biosynthesis of linoleic acid in insects. Biochim. et Biophys. Acta (BBA) Lipids Lipid Metab. 1986, 876, 572–580. [Google Scholar] [CrossRef]
- De Renobales, M.; Cripps, C.; Stanley-Samuelson, D.W.; Jurenka, R.A.; Blomquist, G.J. Biosynthesis of linoleic acid in insects. Trends Biochem. Sci. 1987, 12, 364–366. [Google Scholar] [CrossRef]
- Malcicka, M.; Visser, B.; Ellers, J. An Evolutionary Perspective on Linoleic Acid Synthesis in Animals. Evol. Biol. 2017, 45, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Perez, V.; Chang, E.T. Sodium-to-Potassium Ratio and Blood Pressure, Hypertension, and Related Factors. Adv. Nutr. 2014, 5, 712–741. [Google Scholar] [CrossRef]
- Morrissey, E.; Giltinan, M.; Kehoe, L.; Nugent, A.P.; McNulty, B.A.; Flynn, A.; Walton, J. Sodium and Potassium Intakes and Their Ratio in Adults (18–90 y): Findings from the Irish National Adult Nutrition Survey. Nutrients 2020, 12, 938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronaghy, H.A. The Role of Zinc in Human Nutrition. World Rev. Nutr. Diet. 1987, 54, 237–254. [Google Scholar] [CrossRef] [PubMed]
- Sidor, E.; Dżugan, M. Drone Brood Homogenate as Natural Remedy for Treating Health Care Problem: A Scientific and Practical Approach. Molecules 2020, 25, 5699. [Google Scholar] [CrossRef] [PubMed]
- Lecocq, A.; Foley, K.; Jensen, A.B. Drone brood production in Danish apiaries and its potential for human consumption. J. Apic. Res. 2018, 57, 331–336. [Google Scholar] [CrossRef]
- Herren, P.; Fieseler, L.; Ambuehl, D.; Grunder, J. Foodborne bacteria in raw drone brood of Apis mellifera–a preliminary survey. J. Insects Food Feed. 2021, 7, 281–285. [Google Scholar] [CrossRef]
- Ambühl, D. Beeza—The Honey Bee Cook Book; Skyfood: Zürich, Switzerland, 2016. [Google Scholar]
- Evans, J.D.; Müller, A.; Jensen, A.B.; Dahle, B.; Flore, R.; Eilenberg, J.; Frøst, M.B. A descriptive sensory analysis of honeybee drone brood from Denmark and Norway. J. Insects Food Feed 2016, 2, 277–283. [Google Scholar] [CrossRef]
- National Institute of Agriculral Sciences, Rural Development Administration. Development Policy and Plans for Korean Insect Industry; Rural Development Administration: Jeonju-si, Korea, 2020; p. 51. [Google Scholar]
- Ulmer, M.; Smetana, S.; Heinz, V. Utilizing honeybee drone brood as a protein source for food products: Life cycle assessment of apiculture in Germany. Resour. Conserv. Recycl. 2019, 154, 104576. [Google Scholar] [CrossRef]
- Govorushko, S. Global status of insects as food and feed source: A review. Trends Food Sci. Technol. 2019, 91, 436–445. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Amino Acid | Apis mellifera carnica | Apis mellifera mellifera | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Prepupa | White Eyed Pupa | Dark Eyed Pupa | F | p | Prepupa | White Eyed Pupa | Dark Eyed Pupa | F | p | |
| Asp | 2.4 ± 0.01 a,A | 2.8 ± 0.02 b,A | 2.8 ± 0.06 b,A | 100.5 | 0.002 | 2.4 ± 0.00 a,A | 2.8 ± 0.01 b,A | 3.0 ± 0.00 c,A | 2338.0 | 0.000 |
| Thr * | 1.3 ± 0.03 a,A | 1.6 ± 0.00 b,A | 1.7 ± 0.02 c,A | 259.8 | 0.000 | 1.4 ± 0.01 a,A | 1.5 ± 0.01 b,A | 1.7 ± 0.00 c,A | 781.5 | 0.000 |
| Ser | 1.4 ± 0.02 a,A | 1.6 ± 0.02 b,A | 1.9 ± 0.01 c,A | 757.6 | 0.000 | 1.4 ± 0.00 a,A | 1.7 ± 0.02 b,A | 2.0 ± 0.00 c,A | 1024.0 | 0.000 |
| Glu | 6.3 ± 0.04 a,A | 7.7 ± 0.12 b,A | 7.4 ± 0.07 b,A | 159.1 | 0.001 | 6.6 ± 0.01 a,A | 7.6 ± 0.03 b,A | 8.1 ± 0.02 c,B | 2598.0 | 0.000 |
| Pro | 2.4 ± 0.08 a,A | 3.0 ± 0.03 b,A | 3.7 ± 0.03 c,A | 321.6 | 0.000 | 2.8 ± 0.01 a,A | 3.3 ± 0.03 b,B | 3.6 ± 0.00 c,A | 945.1 | 0.000 |
| Gly | 1.5 ± 0.00 a,A | 1.9 ± 0.02 b,A | 2.6 ± 0.01 c,A | 4588.0 | 0.000 | 1.6 ± 0.11 a,A | 1.9 ± 0.02 b,A | 2.4 ± 0.01 c,B | 61.4 | 0.000 |
| Ala | 1.5 ± 0.01 a,A | 2.0 ± 0.02 b,A | 2.9 ± 0.02 c,A | 2769.0 | 0.000 | 1.5 ± 0.19 a,A | 2.0 ± 0.01 b,A | 2.5 ± 0.01 b,B | 43.2 | 0.006 |
| Val * | 1.8 ± 0.02 a,A | 2.2 ± 0.02 b,A | 2.5 ± 0.01 c,A | 1044.0 | 0.000 | 1.9 ± 0.00 a,A | 2.2 ± 0.00 b,A | 2.4 ± 0.01 c,A | 4646.0 | 0.000 |
| Cys | 0.2 ± 0.01 a,A | 0.1 ± 0.01 b,A | 0.7 ± 0.01 c,A | 2063.0 | 0.000 | 0.5 ± 0.01 a,B | 0.6 ± 0.01 a,B | 1.0 ± 0.05 b,A | 157.2 | 0.001 |
| Met * | 0.4 ± 0.01 a,A | 0.2 ± 0.00 b,A | 0.4 ± 0.04 a,A | 61.8 | 0.004 | 0.5 ± 0.03 a,A | 0.8 ± 0.02 b,B | 0.8 ± 0.02 b,B | 89.5 | 0.002 |
| Ile * | 1.6 ± 0.02 a,A | 1.9 ± 0.02 b,A | 2.2 ± 0.04 c,A | 201.8 | 0.001 | 1.6 ± 0.00 a,A | 1.9 ± 0.04 b,A | 2.2 ± 0.01 c,A | 218.6 | 0.001 |
| Leu * | 2.6 ± 0.01 a,A | 3.2 ± 0.03 b,A | 3.6 ± 0.04 c,A | 551.9 | 0.000 | 2.7 ± 0.02 a,A | 3.1 ± 0.01 b,A | 3.5 ± 0.02 c,A | 1503.0 | 0.000 |
| Tyr | 1.6 ± 0.01 a,A | 1.7 ± 0.03 b,A | 2.0 ± 0.02 c,A | 256.7 | 0.000 | 1.5 ± 0.01 a,B | 1.8 ± 0.00 b,A | 2.1 ± 0.04 c,A | 335.8 | 0.000 |
| Phe * | 1.4 ± 0.00 a,A | 1.7 ± 0.01 b,A | 1.8 ± 0.00 c,A | 2392.0 | 0.000 | 1.5 ± 0.02 a,A | 1.6 ± 0.02 b,A | 1.8 ± 0.01 c,A | 227.3 | 0.001 |
| His * | 0.8 ± 0.06 a,A | 0.9 ± 0.01 b,A | 1.1 ± 0.00 b,A | 28.6 | 0.011 | 0.8 ± 0.00 a,A | 0.9 ± 0.01 a,A | 1.1 ± 0.06 c,A | 30.0 | 0.010 |
| Lys * | 2.3 ± 0.05 a,A | 2.8 ± 0.04 b,A | 3.2 ± 0.03 c,A | 218.7 | 0.001 | 2.4 ± 0.03 a,A | 2.8 ± 0.02 b,A | 3.1 ± 0.00 c,A | 958.6 | 0.000 |
| Arg | 1.7 ± 0.01 a,A | 2.1 ± 0.01 b,A | 2.3 ± 0.01 c,A | 1212.0 | 0.000 | 1.7 ± 0.04 a,A | 2.0 ± 0.01 b,B | 2.3 ± 0.02 c,A | 290.1 | 0.000 |
| Total | 31.1 a,A | 37.4 b,A | 42.5 c,A | 448.8 | 0.000 | 32.8 a,A | 38.4 b,A | 43.4 c,A | 2817.0 | 0.000 |
| Fatty Acid | Apis mellifera carnica | Apis mellifera mellifera | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Prepupa | White Eyed Pupa | Dark Eyed Pupa | F | p | Prepupa | White Eyed Pupa | Dark Eyed Pupa | F | p | |
| Saturated fatty acids | ||||||||||
| Capric acid (C10:0) | 2.0 ± 0.10 | 1.9 ± 0.03 | 2.0 ± 0.03 | 1.5 | 0.289 | ND | ND | 1.8 ± 0.03 | --- | |
| Lauric acid (C12:0) | 28.2 ± 0.84 a,A | 29.8 ± 0.27 b,A | 27.6 ± 0.20 a,A | 14.5 | 0.005 | 20.9 ± 0.21 a,B | 24.9 ± 0.30 b,B | 26.0 ± 0.12 c,B | 431.5 | 0.000 |
| Myristic acid (C14:0) | 379.3 ± 8.63 a,A | 355.0 ± 3.51 b,A | 234.7 ± 1.87 c,A | 597.9 | 0.000 | 341.7 ± 2.501 a,B | 354.0 ± 2.64 b,A | 284.1 ± 1.34 c,B | 833.8 | 0.000 |
| Palmitic acid (C16:0) | 4699.2 ± 94.42 a,A | 4640.6 ± 36.65 a,A | 3307.0 ± 32.03 b,A | 494.5 | 0.000 | 4847.7 ± 28.24 a,A | 4726.6 ± 24.57 b,B | 3803.9 ± 26.28 c,B | 1402.0 | 0.000 |
| Heptadecanoic acid (C17:0) | 4.2 ± 0.09 a,A | 4.1 ± 0.05 a,A | 4.1 ± 0.07 a,A | 1.5 | 0.300 | 4.3 ± 0.02 a,A | 4.7 ± 0.05 b,B | 4.5 ± 0.03 c,B | 89.1 | 0.000 |
| Stearic acid (C18:0) | 1277.9 ± 20.17 a,A | 1362.7 ± 10.37 b,A | 1207.3 ± 15.90 c,A | 71.0 | 0.000 | 1207.0 ± 7.19 a,B | 1260.0 ± 5.74 b,B | 1181.4 ± 11.29 c,A | 68.2 | 0.000 |
| Arachidic acid (C20:0) | 46.8 ± 0.35 a,A | 60.9 ± 0.84 b,A | 72.4 ± 1.54 c,A | 463.0 | 0.000 | 45.1 ± 0.59 a,B | 58.9 ± 0.69 b,B | 67.7 ± 0.82 c,B | 779.4 | 0.000 |
| Behenic acid (C22:0) | 16.0 ± 0.53 a,A | 20.9 ± 0.52 b,A | 30.3 ± 0.60 c,A | 515.9 | 0.000 | 16.9 ± 0.24 a,A | 21.2 ± 0.52 b,A | 27.6 ± 0.44 c,B | 490.0 | 0.000 |
| Subtotal | 6453.7 a,A | 6475.8 a,A | 4885.4 b,A | 369.6 | 0.000 | 6483.5 a,A | 6450.3 a,A | 5396.9 b,B | 877.1 | 0.000 |
| Monounsaturated fatty acids | ||||||||||
| Myristoleic acid (C14:1) | 2.4 ± 0.07 A | 2.0 ± 0.03 A | ND | 3124.0 | 0.000 | 3.1 ± 0.02 a,B | 3.0 ± 0.02 b,B | 2.4 ± 0.04 c | 472.6 | 0.000 |
| Palmitoleic acid (C16:1) | 55.4 ± 1.21 a,A | 50.8 ± 0.60 b,A | 47.9 ± 0.96 c,A | 47.3 | 0.000 | 72.3 ± 0.21 a,B | 65.6 ± 0.34 b,B | 56.1 ± 0.41 c,B | 1815.0 | 0.000 |
| Oleic acid (C18:1 n-9, Cis) | 4701.8 ± 81.72 a,A | 4771.3 ± 39.85 a,A | 4316.3 ± 25.28 b,A | 60.7 | 0.000 | 4439.6 ± 21.05 a,B | 4578.5 ± 33.87 b,B | 4197.3 ± 38.45 c,B | 109.1 | 0.000 |
| cis11-Eicosenic acid (C20:1 n-9) | 7.3 ± 0.09 a,A | 7.6 ± 0.08 b,A | 9.1 ± 0.13 c,A | 247.4 | 0.000 | 6.6 ± 0.10 a,B | 7.6 ± 0.06 b,A | 8.5 ± 0.11 c,B | 312.0 | 0.000 |
| Subtotal | 4766.9 a,A | 4831.7 a,A | 4373.3 b,A | 60.1 | 0.000 | 4521.6 a,B | 4654.7 b,B | 4264.2 c,B | 113.3 | 0.000 |
| Polyunsaturated fatty acids | ||||||||||
| Linolelaidic acid (C18:2 n-6, trans) | 10.2 ± 0.55 a,A | 13.3 ± 0.96 b,A | 17.3 ± 0.56 c,A | 73.1 | 0.000 | 21.3 ± 4.51 a,B | 22.0 ± 2.40 a,B | 22.2 ± 0.94 a,B | 0.1 | 0.920 |
| Linoleic acid (C18:2 n-6, Cis) | 46.6 ± 0.99 a,A | 49.0 ± 0.52 b,A | 36.3 ± 0.60 c,A | 255.4 | 0.000 | 31.3 ± 0.73 a,B | 53.2 ± 0.44 b,B | 56.8 ± 0.63 c,B | 1533.0 | 0.000 |
| Linolenic acid (C18:3 n-3) | 153.0 ± 3.51 a,A | 151.9 ± 1.63 a,A | 154.1 ± 2.27 a,A | 0.5 | 0.615 | 77.4 ± 0.37 a,B | 98.4 ± 1.14 b,B | 118.7 ± 2.74 c,B | 428.9 | 0.000 |
| cis-11,14,17-Eicosatrienoic acid (C20:3 n-3) | ND | ND | 1.8 ± 0.08 | --- | ND | ND | ND | --- | ||
| cis-13,16-Docosadienoic acid (C22:2) | 14.9 ± 0.27 a,A | 18.8 ± 0.12 b,A | 26.2 ± 0.12 c,A | 2808.0 | 0.000 | 13.0 ± 0.49 a,B | 16.8 ± 0.48 b,B | 19.4 ± 0.09 c,B | 197.7 | 0.000 |
| cis-5,8,11,14,17-Eicosapentaenoic acid (C20:5 n-3) | 3.9 ± 0.60 a,A | 6.0 ± 1.06 a b,A | 7.3 ± 0.42 b,c,A | 16.0 | 0.004 | 6.5 ± 3.39 a,A | 7.4 ± 1.81 a,A | 6.6 ± 0.83 a,A | 0.1 | 0.879 |
| Subtotal | 228.6 a,A | 239.0 b,A | 242.8 b,A | 11.7 | 0.009 | 149.4 a,B | 197.7 b,B | 223.8 c,B | 116.8 | 0.000 |
| Total | 11449.2 a,A | 11546.5 a,A | 9501.5 b,A | 204.4 | 0.000 | 11154.6 a,A | 11302.7 a,B | 9884.9 b,B | 413.9 | 0.000 |
| Apis mellifera carnica | Apis mellifera mellifera | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Prepupa | White Eyed Pupa | Dark Eyed Pupa | F | P | Prepupa | White Eyed Pupa | Dark Eyed Pupa | F | p | |
| Ca | 34.0 ± 0.68 a,A | 37.9 ± 0.47 b,A | 46.1 ± 0.38 c,A | 415.9 | 0.000 | 39.3 ± 0.36 a,B | 40.1 ± 0.33 a,B | 43.3 ± 0.82 b,B | 45.9 | 0.000 |
| Mg | 65.9 ± 1.42 a,A | 74.3 ± 0.63 b,A | 88.4 ± 0.25 c,A | 465.4 | 0.000 | 70.2 ± 0.54 a,B | 75.3 ± 0.84 b,A | 85.8 ± 1.01 c,A | 279.4 | 0.000 |
| Na | 7.8 ± 0.59 a,A | 7.0 ± 0.01 a,A | 10.3 ± 0.42 b,A | 49.6 | 0.000 | 8.1 ± 0.25 a,A | 8.7 ± 0.05 b,B | 9.9 ± 0.05 c,A | 111.7 | 0.000 |
| K | 1048.9 ± 23.41 a,A | 1219.8 ± 7.26 b,A | 1401.2 ± 3.88 c,A | 453.7 | 0.000 | 1079.9 ± 4.64 a,A | 1205.2 ± 18.92 b,A | 1341.6 ± 12.48 c,B | 287.9 | 0.000 |
| P | 651.7 ± 14.92 a,A | 734.7 ± 3.39 b,A | 869.2 ± 4.62 c,A | 424.0 | 0.000 | 673.5 ± 3.21 a,A | 731.3 ± 4.18 b,A | 812.3 ± 12.30 c,B | 244.3 | 0.000 |
| Fe | 5.6 ± 0.65 a,A | 5.7 ± 0.07 a,A | 6.1 ± 0.04 a b,A | 1.0 | 0.407 | 4.7 ± 0.09 a,A | 5.2 ± 0.09 b,B | 5.7 ± 0.06 c,B | 119.8 | 0.000 |
| Zn | 4.8 ± 0.08 a,A | 5.3 ± 0.02 b,A | 6.0 ± 0.04 c,A | 436.6 | 0.000 | 4.4 ± 0.02 a,B | 4.9 ± 0.05 b,B | 5.5 ± 0.04 c,B | 597.1 | 0.000 |
| Cu | 1.6 ± 0.05 a,A | 1.8 ± 0.01 b,A | 2.0 ± 0.02 c,A | 111.6 | 0.000 | 1.5 ± 0.01 a,B | 1.6 ± 0.03 b,B | 1.9 ± 0.06 c,A | 76.9 | 0.000 |
| Apis mellifera carnica | Apis mellifera mellifera | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Prepupa | White Eyed Pupa | Dark Eyed Pupa | Prepupa | White Eyed Pupa | Dark Eye Pupa | |||||||
| Male | Female | Male | Female | Male | Female | Male | Female | Male | Female | Male | Female | |
| Ca | 3.4 | 3.4 | 3.8 | 3.8 | 4.6 | 4.6 | 3.9 | 3.9 | 4.0 | 4.0 | 4.3 | 4.3 |
| Mg | 15.7 | 20.6 | 17.7 | 23.2 | 21.0 | 27.6 | 16.7 | 21.9 | 17.9 | 23.5 | 20.4 | 26.8 |
| Na | 0.5 | 0.5 | 0.5 | 0.5 | 0.7 | 0.7 | 0.5 | 0.5 | 0.6 | 0.6 | 0.7 | 0.7 |
| K | 30.9 | 40.3 | 35.9 | 46.9 | 41.2 | 53.9 | 31.8 | 41.5 | 35.4 | 46.4 | 39.5 | 51.6 |
| P | 93.1 | 93.1 | 105.0 | 105.0 | 124.2 | 124.2 | 96.2 | 96.2 | 104.5 | 104.5 | 116.0 | 116.0 |
| Fe | 70.0 | 31.1 | 71.3 | 31.7 | 76.3 | 33.9 | 58.8 | 26.1 | 65.0 | 28.9 | 71.3 | 31.7 |
| Zn | 43.6 | 60.0 | 48.2 | 66.3 | 54.5 | 75.0 | 40.0 | 55.0 | 44.5 | 61.3 | 50.0 | 68.8 |
| Cu | 177.8 | 177.8 | 200.0 | 200.0 | 222.2 | 222.2 | 166.7 | 166.7 | 177.8 | 177.8 | 211.1 | 211.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh, S.; Herren, P.; Meyer-Rochow, V.B.; Jung, C. Nutritional Composition of Honey Bee Drones of Two Subspecies Relative to Their Pupal Developmental Stages. Insects 2021, 12, 759. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080759
Ghosh S, Herren P, Meyer-Rochow VB, Jung C. Nutritional Composition of Honey Bee Drones of Two Subspecies Relative to Their Pupal Developmental Stages. Insects. 2021; 12(8):759. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080759
Chicago/Turabian StyleGhosh, Sampat, Pascal Herren, Victor Benno Meyer-Rochow, and Chuleui Jung. 2021. "Nutritional Composition of Honey Bee Drones of Two Subspecies Relative to Their Pupal Developmental Stages" Insects 12, no. 8: 759. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12080759