
Fungal Communities Vectored by Ips sexdentatus in Declining Pinus sylvestris in Ukraine: Focus on Occurrence and Pathogenicity of Ophiostomatoid Species

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
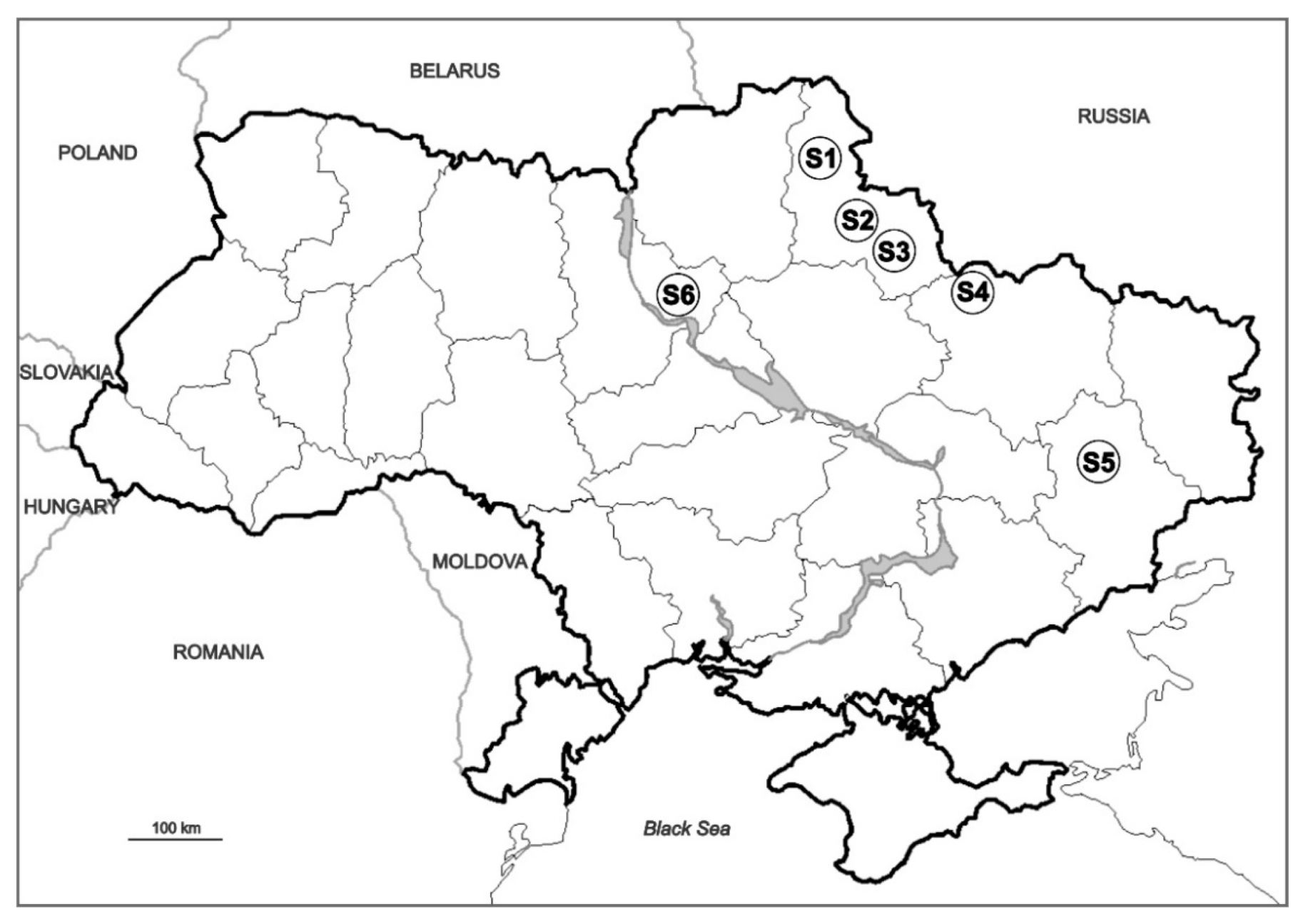
2.1. Collection of Insects
2.2. Sampling from Wood beyond Ips Sexdentatus Galleries
2.3. Isolation and Morphological Grouping of Fungi
2.4. DNA Extraction, Amplification and Sequencing
2.5. Pathogenicity Tests
2.6. Vector Test
2.7. Statistical Analyses
3. Results
3.1. Fungal Communities Associated with Ips Sexdentatus
3.2. Pathogenicity
3.3. Vector Test
4. Discussion
4.1. Ophiostomatoid Fungi
4.2. Pathogenicity
4.3. Other Fungi
4.4. Ips Sexdentatus as a Vector
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Trowbridge, A.M.; Adams, H.D.; Collins, A.; Dickman, L.T.; Grossiord, C.; Hofland, M.; Malone, S.; Weaver, D.K.; Sevanto, S.; Stoy, P.C.; et al. Hotter droughts alter resource allocation to chemical defenses in pinon pine. Oecologia 2021, 197, 921–938. [Google Scholar] [CrossRef] [PubMed]
- Wermelinger, B.; Rigling, A.; Mathis, D.S.; Kenis, M.; Gossner, M.M. Climate change effects on trophic interactions of bark beetles in inner alpine Scots pine forests. Forests 2021, 12, 136. [Google Scholar] [CrossRef]
- Netherer, S.; Panassiti, B.; Pennerstorfer, J.; Matthews, B. Acute drought is an important driver of bark beetle infestation in Austrian Norway spruce stands. Front. For. Glob. Chang. 2019, 2, 39. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, A.; Modlinger, R.; Ashraf, M.Z.; Synek, J.; Schlyter, F.; Roy, A. Core mycobiome and their ecological relevance in the gut of five Ips bark beetles (Coleoptera: Curculionidae: Scolytinae). Front. Microbiol. 2020, 11, 568853. [Google Scholar] [CrossRef] [PubMed]
- Netherer, S.; Kandasamy, D.; Jirosova, A.; Kalinova, B.; Schebeck, M.; Schlyter, F. Interactions among Norway spruce, the bark beetle Ips typographus and its fungal symbionts in times of drought. J. Pest Sci. 2021, 94, 591–614. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, M.M.F. Colonization of fire-damaged trees by Ips sexdentatus (Boerner) as related to the percentage of burnt crown. Entomol. Fennica 2006, 17, 381–386. [Google Scholar] [CrossRef] [Green Version]
- Rossi, J.P.; Samalens, J.C.; Guyon, D.; van Halder, I.; Jactel, H.; Menassieu, P.; Piou, D. Multiscale spatial variation of the bark beetle Ips sexdentatus damage in a pine plantation forest (Landes de Gascogne, Southwestern France). For. Ecol. Manag. 2009, 257, 1551–1557. [Google Scholar] [CrossRef]
- Bueno, A.; Diez, J.J.; Fernandez, M.M. Ophiostomatoid fungi transported by Ips sexdentatus (Coleoptera, Scolytidae) in Pinus pinaster in NW Spain. Silva Fennica 2010, 44, 137. [Google Scholar] [CrossRef] [Green Version]
- Pineau, X.; David, G.; Peter, Z.; Sallé, A.; Baude, M.; Lieutier, F.; Jactel, H. Effect of temperature on the reproductive success, developmental rate and brood characteristics of Ips sexdentatus (Boern.). Agric. For. Entomol. 2016, 19, 23–33. [Google Scholar] [CrossRef]
- Meshkova, V. The lessons of Scots pine forest decline in Ukraine. Environ. Sci. Proc. 2021, 3, 28. [Google Scholar] [CrossRef]
- Toth, E.G.; Köbölkuti, Z.A.; Pedryc, A.; Höhn, M. Evolutionary history and phylogeography of Scots pine (Pinus sylvestris L.) in Europe based on molecular markers. J. For. Res. 2017, 28, 637–651. [Google Scholar] [CrossRef]
- Kirisits, T. Fungal associates of European bark beetles with special emphasis on the Ophiostomatoid fungi. In Bark and Wood Boring Insects in Living Trees in Europe, a Synthesis; Lieutier, F., Day, K.R., Battisti, A., Grégoire, J.-C., Evans, H.F., Eds.; Kluwer: Dordrecht, The Netherlands, 2004; pp. 181–236. [Google Scholar] [CrossRef]
- Jankowiak, R. Ophiostomatoid fungi associated with Ips sexdentatus on Pinus sylvestris in Poland. Dendrobiology 2012, 68, 43–53. [Google Scholar]
- Mathiesen-Käärik, A. Eine Übersicht über die gewönlichsten mit Borkenäfern assoziierten Bläuepilze in Schweden und einige für Schweden neue Bläupilze. Medd. St. Skogsforskningsinst. 1953, 43, 1–74. [Google Scholar]
- Linnakoski, R.; de Beer, Z.W.; Niemelä, P.; Wingfield, M.J. Associations of conifer-infesting bark beetles and fungi in Fennoscandia. Insects 2012, 3, 200–227. [Google Scholar] [CrossRef] [Green Version]
- Lieutier, F.; Yart, A.; Garcia, J.; Ham, M.C.; Morelet, M.; Levieux, J. Phytopathogenic fungi associated with two bark beetles of Scots pine (Pinus sylvestris L.) and preliminary study of their aggressiveness for the host. Ann. For. Sci. 1989, 46, 201–216. [Google Scholar] [CrossRef] [Green Version]
- Lieutier, F.; Garcia, J.; Yart, A.; Vouland, G.; Pettinetti, M.; Morelet, M. Ophiostomatales (Ascomycetes) associated with Ips acuminatus Gyll (Coleoptera, Scolytidae) in Scots pine (Pinus sylvestris L) in South-Eastern France, and comparison with Ips sexdentatus Boern. Agronomie 1991, 11, 807–817. [Google Scholar] [CrossRef] [Green Version]
- Romon, P.; Zhou, X.-D.; Iturrondobeitia, J.C.J.; Wingfield, M.J.; Goldarazena, A. Ophiostoma species (Ascomycetes: Ophiostomatales) associated with bark beetles (Coleoptera: Scolytinae) colonizing Pinus radiata in northern Spain. Can. J. Microbiol. 2007, 53, 756–767. [Google Scholar] [CrossRef]
- Bezos, D.; Martinez-Alvarez, P.; Sanz-Ros, A.; Martin-Garcia, J.; Fernandez, M.; Diez, J. Fungal communities associated with bark beetles in Pinus radiata plantations in northern Spain affected by pine pitch canker, with special focus on Fusarium species. Forests 2018, 9, 698. [Google Scholar] [CrossRef] [Green Version]
- Davydenko, K.; Vasaitis, R.; Meshkova, V.; Menkis, A. Fungi associated with the red-haired bark beetle, Hylurgus ligniperda (Coleoptera: Curculionidae) in the forest-steppe zone in eastern Ukraine. Eur. J. Entomol. 2014, 111, 561–565. [Google Scholar] [CrossRef] [Green Version]
- Davydenko, K.; Vasaitis, R.; Menkis, A. Fungi associated with Ips acuminatus (Coleoptera: Curculionidae) in Ukraine with a special emphasis on pathogenicity of ophiostomatoid species. Eur. J. Entomol. 2017, 114, 77–85. [Google Scholar] [CrossRef]
- Leach, J.G. Insect Transmission of Plant Diseases; McGraw-Hill Book: New York, NY, USA, 1940. [Google Scholar]
- Solheim, H.; Langström, B.; Hellqvist, C. Pathogenicity of the blue-stain fungi Leptographium-wingfieldii and Ophiostoma minus to Scots pine—Effect of tree pruning and inoculum density. Can. J. For. Res. 1993, 23, 1438–1443. [Google Scholar] [CrossRef]
- Menkis, A.; Vasiliauskas, R.; Taylor, A.F.S.; Stenström, E.; Stenlid, J.; Finlay, R. Fungi in decayed roots of conifer seedlings in forest nurseries, afforested clear-cuts and abandoned farmland. Plant Pathol. 2006, 55, 117–129. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for Basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, USA, 1990; pp. 315–321. [Google Scholar]
- Persson, Y.; Vasaitis, R.; Langström, B.; Öhrn, P.; Ihrmark, K.; Stenlid, J. Fungi vectored by the bark beetle Ips typographus following hibernation under the bark of standing trees and in the forest litter. Microb. Ecol. 2009, 58, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Krokene, P.; Solheim, H. Pathogenicity of four blue-stain fungi associated with aggressive and non-aggressive bark beetles. Phytopathol. 1998, 88, 39–44. [Google Scholar] [CrossRef] [Green Version]
- Bezos, D.; Martinez-Alvarez, P.; Diez, J.; Fernandez, M. The pine shoot beetle Tomicus piniperda as a plausible vector of Fusarium circinatum in northern Spain. Ann. For. Sci. 2015, 72, 1079–1088. [Google Scholar] [CrossRef] [Green Version]
- Ploetz, R.C.; Hulcr, J.; Wingfield, M.J.; de Beer, Z.W. Destructive tree diseases associated with ambrosia and bark beetles: Black swan events in tree pathology? Plant Dis. 2013, 97, 856–872. [Google Scholar] [CrossRef] [Green Version]
- Linnakoski, R.; Jankowiak, R.; Villari, C.; Kirisits, T.; Solheim, H.; de Beer, Z.W.; Wingfield, M.J. The Ophiostoma clavatum species complex: A newly defined group in the Ophiostomatales including three novel taxa. Antonie van Leeuwenhoek 2016, 109, 987–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, M.M.F.; Garcia, A.E.; Lieutier, F. Effects of various densities of Ophiostoma ips inoculations on Pinus sylvestris in north-western Spain. For. Pathol. 2004, 34, 213–223. [Google Scholar] [CrossRef]
- Levieux, J.; Lieutier, F.; Moser, C.; Perry, T.J. Transportation of phytopathogenic fungi by the bark beetle Ips sexdentatus Boerner and associated mites. J. Appl. Entomol. 1989, 108, 1–11. [Google Scholar] [CrossRef]
- Jankowiak, R.; Solheim, H.; Bilanski, P.; Marincowitz, S.; Wingfield, M.J. Seven new species of Graphilbum from conifers in Norway, Poland, and Russia. Mycologia 2020, 112, 1240–1262. [Google Scholar] [CrossRef] [PubMed]
- Jankowiak, R.; Szewczyk, G.; Bilanski, P.; Jazlowiecka, D.; Harabin, B.; Linnakoski, R. Blue-stain fungi isolated from freshly felled Scots pine logs in Poland, including Leptographium sosnaicola sp. nov. For. Pathol. 2021, 51, e12672. [Google Scholar] [CrossRef]
- Bracewell, R.R.; Six, D.L. Experimental evidence of bark beetle adaptation to a fungal symbiont. Ecol. Evol. 2015, 5, 5109–5119. [Google Scholar] [CrossRef] [PubMed]
- Kolarik, M.; Jankowiak, R. Vector affinity and diversity of Geosmithia fungi living on subcortical insects inhabiting Pinaceae species in Central and Northeastern Europe. Microbial Ecol. 2013, 66, 682–700. [Google Scholar] [CrossRef] [PubMed]
- Hofstetter, R.W.; Gandhi, K.J. Interactions among climate, disturbance, and bark beetles affect forest landscapes of the future. In Bark Beetle Management, Ecology, and Climate Change; Gandhi, K.J., Hofstetter, R.W., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 395–404. [Google Scholar] [CrossRef]


{kind=link}
| Taxa | Genbank | Detected in Beetles, % (No. Examined) | ||
|---|---|---|---|---|
| Accession No. | Direct Sequencing (288) | Isolations (288) | All (576) | |
| Ophiostomatoid (species sensu lato) | ||||
| Graphium sp. KD5 | OK576216 | 1.7 | 3.5 | 2.6 |
| Grosmannia penicillata | OK576218 | 1.7 | 2.8 | 2.3 |
| Leptographium olivaceum | OK576217 | 2.8 | 3.8 | 3.3 |
| Leptographium sosnaicola | OK576219 | 4.9 | 4.9 | 4.9 |
| Ophiostoma bicolor | OK576220 | - | 1.7 | 0.9 |
| Ophiostoma canum | OK576221 | 5.9 | 2.4 | 4.2 |
| Ophiostoma ips | OK576222 | 17.0 | 1.0 | 9.0 |
| Ophiostoma minus | OK576223 | 7.6 | 1.4 | 4.5 |
| Ophiostoma piceae | OK576224 | 1.0 | 0.7 | 0.9 |
| Other Ascomycota | ||||
| Alternaria alternata | OK576225 | 1.4 | 1.4 | 1.4 |
| Anthostomella pinea | OK576226 | 2.4 | 2.4 | 2.4 |
| Aspergillus versicolor | OK576227 | 4.2 | 1.0 | 2.6 |
| Aureobasidium pullulans | OK576228 | 2.8 | 2.4 | 2.6 |
| Beauveria bassiana | OK576229 | 2.1 | 9.0 | 5.6 |
| Beauveria pseudobassiana | OK576230 | - | 2.1 | 1.0 |
| Bionectria ochroleuca | OK576231 | 6.6 | 2.8 | 4.7 |
| Botryotinia fuckeliana | OK576232 | 9.4 | 3.1 | 6.3 |
| Chaetomium sp. | OK576233 | 5.2 | - | 2.6 |
| Chalara sp. | OK576234 | 2.1 | 2.1 | 2.1 |
| Cladobotryum mycophilum | OK576235 | 5.9 | - | 3.0 |
| Cladosporium cladosporioides | OK576236 | 12.5 | - | 6.3 |
| Cladosporium sp. | OK576237 | 22.2 | - | 11.1 |
| Clavispora lusitaniae | OK576238 | 2.1 | - | 1.0 |
| Cordyceps farinose | OK576239 | 0.7 | 1.7 | 1.2 |
| Cyclaneusma minus | OK576240 | 2.8 | 6.3 | 4.5 |
| Dactylonectria macrodidyma | OK576241 | 3.8 | 2.1 | 3.0 |
| Diplodia sapinea | OK576242 | 2.4 | 5.2 | 3.8 |
| Fusarium avenaceum | OK576243 | 3.1 | 10.1 | 6.6 |
| Fusarium sp. | OK576244 | - | 0.7 | 0.4 |
| Leptodontidium beauverioides | OK576245 | 11.1 | - | 5.6 |
| Lophodermium seditiosum | OK576246 | 2.1 | 11.1 | 6.6 |
| Mariannaea elegans | OK576247 | 1.4 | 6.3 | 3.8 |
| Metapochonia bulbilosa | OK576248 | 1.4 | 9.4 | 5.4 |
| Nectria sp. | OK576249 | 2.8 | - | 1.4 |
| Penicillium citreonigrum | OK576250 | - | 4.5 | 2.3 |
| Pezicula eucrita | OK576251 | 0.7 | 1.4 | 1.0 |
| Phomopsis sp. | OK576252 | 1.4 | - | 0.7 |
| Sydowia polyspora | OK576253 | 1.4 | 3.5 | 2.4 |
| Talaromyces ruber | OK576254 | 5.2 | - | 2.6 |
| Trichoderma asperellum | OK576255 | 2.4 | 3.5 | 3.0 |
| Trichoderma sp. | - | - | 2.8 | 1.4 |
| Basidiomycota | ||||
| Entomocorticium sp. | OK576256 | 24.7 | - | 12.3 |
| Filobasidium magnum | OK576257 | 5.2 | - | 2.6 |
| Fomitopsis pinicola | OK576258 | 0.7 | 8.3 | 4.5 |
| Heterobasidion annosum | OK576259 | 5.6 | 0.3 | 3.0 |
| Phlebiopsis gigantea | OK576260 | 3.1 | 1.0 | 2.1 |
| Mucoromycotina | ||||
| Mortierella gemmifera | OK576261 | - | 1.7 | 0.9 |
| Mucor fragilis | OK576262 | 5.9 | - | 3.0 |
| Mucor sp. | - | - | 1.7 | 0.9 |
| Umbelopsis isabellina | OK576263 | 3.8 | 6.9 | 5.4 |
| Unidentified fungi | ||||
| Fungal sp. A | OK576264 | 0.3 | 5.2 | 2.8 |
| Fungal sp. HH78_19 | OK576265 | 1.0 | - | 0.5 |
| Fungal sp. K11 | OK576266 | 1.7 | 3.8 | 2.8 |
| Fungal sp. K21 | OK576267 | 1.7 | - | 0.9 |
| Fungal sp. K23 | OK576268 | 0.3 | 2.1 | 1.2 |
| Fungal sp. K27 | OK576269 | - | 3.1 | 1.6 |
| Total, no. (detected exclusively by the method) | 48 (13) | 43 (8) | 56 | |
| Site | S2 | S3 | S4 | S5 | S6 |
|---|---|---|---|---|---|
| S1 | 0.37 | 0.38 | 0.37 | 0.37 | 0.38 |
| S2 | 0.28 | 0.28 | 0.28 | 0.28 | |
| S3 | 0.26 | 0.26 | 0.26 | ||
| S4 | 0.24 | 0.25 | |||
| S5 | 0.23 |
| Fungus | Symptomatic Saplings, % (12 Tested per Fungus & Control) | Lesion Length, mm | Re-Isolation Frequency, | ||
|---|---|---|---|---|---|
| Dead | Dieback Symptoms a | All | (Mean ± SE) b | % | |
| Graphium sp. KD5 | 58.3 | 41.7 | 100 | 14.7 ± 1.2 | 87.5 |
| Grosmannia penicillata | 33.3 | 16.7 | 50 | 9.6 ± 1.9 A | 87.5 |
| Leptographiumolivaceum | 16.7 | 16.7 | 33.4 | 6.5 ± 0.2 | 91.7 |
| Leptographium sosnaicola | 75.0 | 25.0 | 100 | 25.9 ± 1.2 | 91.7 |
| Ophiostoma bicolor | 8.3 | 25.0 | 33.3 | 10.2 ± 0.3 A | 100 |
| Ophiostoma canum | 0 | 0 | 0 | 3.4 ± 0.1 | 62.5 |
| Control | 0 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davydenko, K.; Vasaitis, R.; Elfstrand, M.; Baturkin, D.; Meshkova, V.; Menkis, A. Fungal Communities Vectored by Ips sexdentatus in Declining Pinus sylvestris in Ukraine: Focus on Occurrence and Pathogenicity of Ophiostomatoid Species. Insects 2021, 12, 1119. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121119
Davydenko K, Vasaitis R, Elfstrand M, Baturkin D, Meshkova V, Menkis A. Fungal Communities Vectored by Ips sexdentatus in Declining Pinus sylvestris in Ukraine: Focus on Occurrence and Pathogenicity of Ophiostomatoid Species. Insects. 2021; 12(12):1119. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121119
Chicago/Turabian StyleDavydenko, Kateryna, Rimvydas Vasaitis, Malin Elfstrand, Denys Baturkin, Valentyna Meshkova, and Audrius Menkis. 2021. "Fungal Communities Vectored by Ips sexdentatus in Declining Pinus sylvestris in Ukraine: Focus on Occurrence and Pathogenicity of Ophiostomatoid Species" Insects 12, no. 12: 1119. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121119