
Wolbachia Detection in Field-Collected Mosquitoes from Cameroon

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Samples
2.2. MALDI TOF MS Analyses and Molecular Identification of Mosquitoes
2.3. Mosquito Microbiota Analysis
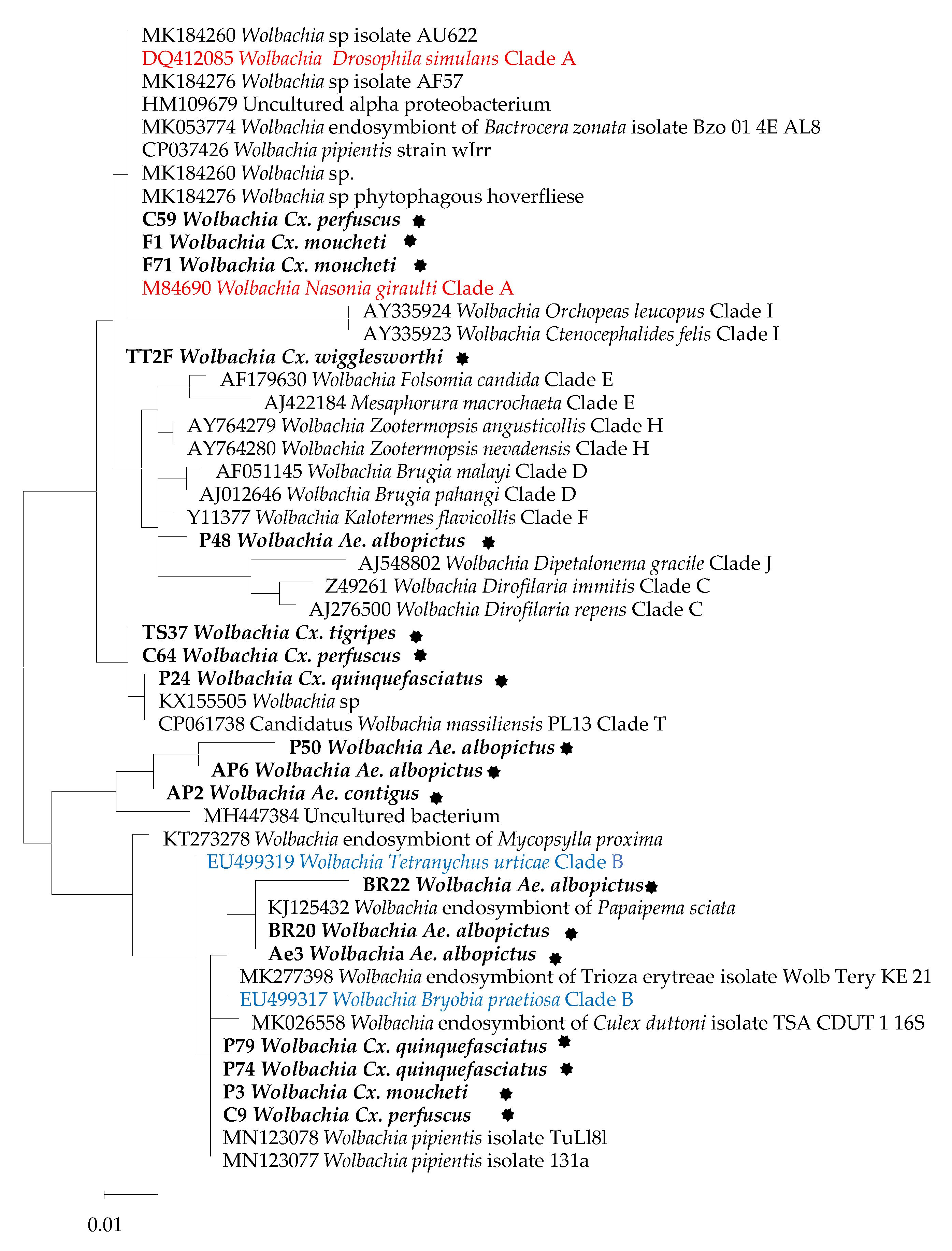
2.4. Sequencing and Phylogenetic Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sinka, M.E.; Bangs, M.J.; Manguin, S.; Rubio-Palis, Y.; Chareonviriyaphap, T.; Coetzee, M.; Mbogo, C.M.; Hemingway, J.; Patil, A.P.; Temperley, W.H.; et al. A Global Map of Dominant Malaria Vectors. Parasites Vectors 2012, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Becker, N.; Petrić, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Medical Importance of Mosquitoes. In Mosquitoes and Their Control; Springer: Berlin/Heidelberg, Germany, 2010; pp. 25–42. [Google Scholar] [CrossRef]
- Azari-Hamidian, S.; Norouzi, B.; Harbach, R.E. A Detailed Review of the Mosquitoes (Diptera: Culicidae) of Iran and Their Medical and Veterinary Importance. Acta Trop. 2019, 194, 106–122. [Google Scholar] [CrossRef]
- Bamou, R.; Mayi, M.P.A.; Djiappi-Tchamen, B.; Nana-Ndjangwo, S.M.; Nchoutpouen, E.; Cornel, A.J.; Awono-Ambene, P.; Parola, P.; Tchuinkam, T.; Antonio-Nkondjio, C. An Update on the Mosquito Fauna and Mosquito-Borne Diseases Distribution in Cameroon. Parasites Vectors 2021, 14, 1–15. [Google Scholar] [CrossRef]
- Barua, S.; Hoque, M.M.; Kelly, P.J.; Poudel, A.; Adekanmbi, F.; Kalalah, A.; Yang, Y.; Wang, C. First Report of Rickettsia felis in Mosquitoes, USA. Emerg. Microbes Infect. 2020, 9, 1008–1010. [Google Scholar] [CrossRef] [Green Version]
- Dieme, C.; Bechah, Y.; Socolovschi, C.; Audoly, G.; Berenger, J.-M.; Faye, O.; Raoult, D.; Parola, P. Transmission Potential of Rickettsia felis Infection by Anopheles gambiae Mosquitoes. Proc. Natl. Acad. Sci. USA 2015, 112, 201413835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. World Malaria Report; World Health Organization: Geneva, Switzerland, 2020; Available online: https://www.who.int/publications/i/item/9789240015791 (accessed on 22 December 2020).
- Gomes, F.M.; Hixson, B.L.; Tyner, M.D.W.; Ramirez, J.L.; Canepa, G.E.; Alves e Silva, T.L.; Molina-Cruz, A.; Keita, M.; Kane, F.; Traoré, B.; et al. Effect of Naturally Occurring Wolbachia in Anopheles gambiae s.l. Mosquitoes from Mali on Plasmodium falciparum Malaria Transmission. Proc. Natl. Acad. Sci. USA 2017, 114, 12566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeffries, C.L.; Lawrence, G.G.; Golovko, G.; Kristan, M.; Orsborne, J.; Spence, K.; Hurn, E.; Bandibabone, J.; Tantely, L.M.; Raharimalala, F.N.; et al. Novel Wolbachia Strains in Anopheles Malaria Vectors from Sub-Saharan Africa. Wellcome Open Res. 2018, 3, 113. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.; Quek, S.; Jeffries, C.L.; Bandibabone, J.; Dhokiya, V.; Bamou, R.; Kristan, M.; Messenger, L.A.; Gidley, A.; Hornett, E.A.; et al. Stable High-Density and Maternally Inherited Wolbachia Infections in Anopheles moucheti and Anopheles demeilloni Mosquitoes. Curr. Biol. 2021, 31, 2310–2320.e5. [Google Scholar] [CrossRef]
- Wong, M.L.; Liew, J.W.K.; Wong, W.K.; Pramasivan, S.; Mohamed Hassan, N.; Wan Sulaiman, W.Y.; Jeyaprakasam, N.K.; Leong, C.S.; Low, V.L.; Vythilingam, I. Natural Wolbachia Infection in Field-Collected Anopheles and Other Mosquito Species from Malaysia. Parasites Vectors 2020, 13, 414. [Google Scholar] [CrossRef] [PubMed]
- Kittayapong, P.; Milne, J.R.; Tigvattananont, S.; Baimai, V. Distribution of the Reproduction-Modifying Bacteria, Wolbachia, in Natural Populations of Tephritid Fruit Flies in Thailand. Sci. Asia 2000, 26, 93–103. [Google Scholar] [CrossRef]
- Nugapola, N.W.N.P.; De Silva, W.A.P.P.; Karunaratne, S.H.P.P. Distribution and Phylogeny of Wolbachia Strains in Wild Mosquito Populations in Sri Lanka. Parasites Vectors 2017, 10, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master Manipulators of Invertebrate Biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Zug, R.; Hammerstein, P. Still a Host of Hosts for Wolbachia: Analysis of Recent Data Suggests That 40% of Terrestrial Arthropod Species Are Infected. PLoS ONE 2012, 7, e38544. [Google Scholar] [CrossRef] [Green Version]
- Ayala, D.; Akone-Ella, O.; Rahola, N.; Kengne, P.; Ngangue, M.F.; Mezeme, F.; Makanga, B.K.; Nigg, M.; Costantini, C.; Simard, F.; et al. Natural Wolbachia Infections Are Common in the Major Malaria Vectors in Central Africa. Evol. Appl. 2019, 12, 1583–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrostek, E.; Pelz-Stelinski, K.; Hurst, G.D.D.; Hughes, G.L. Horizontal Transmission of Intracellular Insect Symbionts via Plants. Front. Microbiol. 2017, 8, 2237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.Z.; Breinholt, J.W.; Kawahara, A.Y. Evidence for Common Horizontal Transmission of Wolbachia among Butterflies and Moths. BMC Evol. Biol. 2016, 16, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caputo, B.; Moretti, R.; Manica, M.; Serini, P.; Lampazzi, E.; Bonanni, M.; Fabbri, G.; Pichler, V.; Torre, A.; Calvitti, M. A Bacterium against the Tiger: Preliminary Evidence of Fertility Reduction after Release of Aedes albopictus Males with Manipulated Wolbachia Infection in an Italian Urban Area. Pest Manag. Sci. 2020, 76, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Dutra, H.L.C.; Rocha, M.N.; Dias, F.B.S.; Mansur, S.B.; Caragata, E.P.; Moreira, L.A. Wolbachia Blocks Currently Circulating Zika Virus Isolates in Brazilian Aedes aegypti Mosquitoes. Cell Host Microbe 2016, 19, 771–774. [Google Scholar] [CrossRef] [Green Version]
- Caragata, E.P.; Dutra, H.L.C.; Moreira, L.A. Inhibition of Zika Virus by Wolbachia in Aedes aegypti. Microb. Cell 2016, 3, 293–295. [Google Scholar] [CrossRef]
- Pinto, S.B.; Riback, T.I.S.; Sylvestre, G.; Costa, G.; Peixoto, J.; Dias, F.B.S.; Tanamas, S.K.; Simmons, C.P.; Dufault, S.M.; Ryan, P.A.; et al. Effectiveness of Wolbachia-Infected Mosquito Deployments in Reducing the Incidence of Dengue and Other Aedes-Borne Diseases in Niterói, Brazil: A Quasi-Experimental Study. PLoS Negl. Trop. Dis. 2021, 15, e0009556. [Google Scholar] [CrossRef]
- Frentiu, F.D.; Zakir, T.; Walker, T.; Popovici, J.; Pyke, A.T.; van den Hurk, A.; McGraw, E.A.; O’Neill, S.L. Limited Dengue Virus Replication in Field-Collected Aedes aegypti Mosquitoes Infected with Wolbachia. PLoS Negl. Trop. Dis. 2014, 8, e2688. [Google Scholar] [CrossRef] [Green Version]
- Walker, T.; Johnson, P.H.; Moreira, L.A.; Iturbe-Ormaetxe, I.; Frentiu, F.D.; McMeniman, C.J.; Leong, Y.S.; Dong, Y.; Axford, J.; Kriesner, P.; et al. The WMel Wolbachia Strain Blocks Dengue and Invades Caged Aedes aegypti Populations. Nature 2011, 476, 450–453. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, M. Key to the Females of Afrotropical Anopheles Mosquitoes (Diptera: Culicidae). Malar. J. 2020, 19, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillies, M.T. A Supplement to the Anophelinae of Africa South of the Sahara (Afrotropical Region). Publ. S. Afr. Inst. Med. Res. 1987, 55, 1–143. [Google Scholar]
- Jupp, P. Mosquitoes of Southern Africa; Ekogilde Publishers: Johannesburg, South Africa, 1996; p. 156. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Fall, F.K.; Laroche, M.; Bossin, H.; Musso, D.; Parola, P. Performance of MALDI-TOF Mass Spectrometry to Determine the Sex of Mosquitoes and Identify Specific Colonies from French Polynesia. Am. J. Trop. Med. Hyg. 2021, 104, tpmd200031. [Google Scholar] [CrossRef] [PubMed]
- Tandina, F.; Niaré, S.; Laroche, M.; Koné, A.K.; Diarra, A.Z.; Ongoiba, A.; Berenger, J.M.; Doumbo, O.K.; Raoult, D.; Parola, P. Using MALDI-TOF MS to Identify Mosquitoes Collected in Mali and Their Blood Meals. Parasitology 2018, 145, 1170–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yssouf, A.; Almeras, L.; Raoult, D.; Parola, P. Emerging Tools for Identification of Arthropod Vectors. Future Microbiol. 2016, 11, 549–566. [Google Scholar] [CrossRef] [PubMed]
- Diarra, A.Z.; Almeras, L.; Laroche, M.; Berenger, J.-M.; Kone, A.K.; Bocoum, Z.; Dabo, A.; Doumbo, O.; Raoult, D.; Parola, P. Molecular and MALDI-TOF Identification of Ticks and Tick-Associated Bacteria in Mali. PLoS Negl. Trop. Dis. 2017, 11, e0005762. [Google Scholar] [CrossRef] [PubMed]
- Sadeddine, R.; Diarra, A.Z.; Laroche, M.; Mediannikov, O.; Righi, S.; Benakhla, A.; Dahmana, H.; Raoult, D.; Parola, P. Molecular Identification of Protozoal and Bacterial Organisms in Domestic Animals and Their Infesting Ticks from North-Eastern Algeria. Ticks Tick-Borne Dis. 2020, 11, 101330. [Google Scholar] [CrossRef] [PubMed]
- Hamlili, F.Z.; Thiam, F.; Laroche, M.; Diarra, A.Z.; Doucouré, S.; Gaye, P.M.; Fall, C.B.; Faye, B.; Sokhna, C.; Sow, D.; et al. MALDI-TOF Mass Spectrometry for the Identification of Freshwater Snails from Senegal, Including Intermediate Hosts of Schistosomes. PLoS Negl. Trop. Dis. 2021, 15, e0009725. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Baldini, F.; Rougé, J.; Kreppel, K.; Mkandawile, G.; Mapua, S.A.; Sikulu-Lord, M.; Ferguson, H.M.; Govella, N.; Okumu, F.O. First Report of Natural Wolbachia Infection in the Malaria Mosquito Anopheles arabiensis in Tanzania. Parasites Vectors 2018, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Niang, E.H.A.; Bassene, H.; Makoundou, P.; Fenollar, F.; Weill, M.; Mediannikov, O. First Report of Natural Wolbachia Infection in Wild Anopheles funestus Population in Senegal. Malar. J. 2018, 17, 408. [Google Scholar] [CrossRef] [Green Version]
- Kittayapong, P.; Baisley, K.J.; Baimai, V.; O’Neill, S.L. Distribution and Diversity of Wolbachia Infections in Southeast Asian Mosquitoes (Diptera: Culicidae). J. Med. Entomol. 2000, 37, 340–345. [Google Scholar] [CrossRef]
- Wiwatanaratanabutr, I. Geographic Distribution of Wolbachial Infections in Mosquitoes from Thailand. J. Invertebr. Pathol. 2013, 114, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Chrostek, E.; Gerth, M. Is Anopheles gambiae a Natural Host of Wolbachia? mBio 2019, 10, e00784-19. [Google Scholar] [CrossRef] [Green Version]
- Ross, P.A.; Callahan, A.G.; Yang, Q.; Jasper, M.; Arif, M.A.K.; Afizah, A.N.; Nazni, W.A.; Hoffmann, A.A. An Elusive Endosymbiont: Does Wolbachia Occur Naturally in Aedes aegypti? Ecol. Evol. 2020, 10, 1581–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, A.; Yu, W.; Jiang, J.; Sanchez, C.; Karna, A.K.; Martinez, K.J.; Hanley, K.A.; Buenemann, M.; Hansen, I.A.; Xue, R. Wolbachia pipientis Occurs in Aedes aegypti Populations in New Mexico and Florida, USA. Ecol. Evol. 2019, 9, 6148–6156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osei-Poku, J.; Han, C.; Mbogo, C.M.; Jiggins, F.M. Identification of Wolbachia Strains in Mosquito Disease Vectors. PLoS ONE 2012, 7, e49922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiwatanaratanabutr, I.; Zhang, C. Wolbachia Infections in Mosquitoes and Their Predators Inhabiting Rice Field Communities in Thailand and China. Acta Trop. 2016, 159, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Carvajal, T.M.; Hashimoto, K.; Harnandika, R.K.; Amalin, D.M.; Watanabe, K. Detection of Wolbachia in Field-Collected Aedes aegypti Mosquitoes in Metropolitan Manila, Philippines. Parasites Vectors 2019, 12, 361. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Microorganisms | Targeted Sequence | Primers F, R (5′-3′) and Probes p (6FAM-TAMRA) | |
|---|---|---|---|
| qPCR primers | |||
| Anaplasmataceae | 23S | TtAna_F | TGACAGCGTACCTTTTGCAT |
| TtAna_R | GTAACAGGTTCGGTCCTCCA | ||
| TtAna_P | 6FAM-GGATTAGACCCGAAACCAAG | ||
| Bartonella | ITS | Barto_ITS2_F | GGGGCCGTAGCTCAGCTG |
| Barto_ITS2_R | TGAATATATCTTCTCTTCACAATTTC | ||
| Barto_ITS2_P | 6FAM-CGATCCCGTCCGGCTCCACCA | ||
| Borrelia | 16S | Bor_16S_3F | AGCCTTTAAAGCTTCGCTTGTAG |
| Bor_16S_3R | GCCTCCCGTAGGAGTCTGG | ||
| Bor_16S_3P | 6FAM-CCGGCCTGAGAGGGTGAACGG | ||
| ITS4 | Bor_ITS4_F | GGCTTCGGGTCTACCACATCTA | |
| Bor_ITS4_R | CCGGGAGGGGAGTGAAATAG | ||
| Bor_ITS4_P | 6FAM-TGCAAAAGGCACGCCATCACC | ||
| Coxiella burnetii | IS1111A | CB_IS1111_0706F | CAAGAAACGTATCGCTGTGGC |
| CB_IS1111_0706R | CACAGAGCCACCGTATGAATC | ||
| CB_IS1111_0706P | 6FAM-CCGAGTTCGAAACAATGAGGGCTG | ||
| Hyp. Protein IS30A | CB_IS30A_3F | CGCTGACCTACAGAAATATGTCC | |
| CB_IS30A_3R | GGGGTAAGTAAATAATACCTTCTGG | ||
| CB_IS30A_3P | 6FAM-CATGAAGCGATTTATCAATACGTGTATGC | ||
| Rickettsia spp. | gltA | RKND03_F | GTGAATGAAAGATTACACTATTTAT |
| RKND03_R | GTATCTTAGCAATCATTCTAATAGC | ||
| RKND03 P | 6FAM-CTATTATGCTTGCGGCTGTCGGTTC | ||
| Standard PCR primers | |||
| Invertebrate identification (Folmer) | COI | LCO1490 | GGTCAACAAATCATAAAGATATTGG |
| HCO2198 | TAAACTTCAGGGTGACCAAAAAATCA | ||
| Anaplasmataceae | 23S | Ana23S-212f | ATAAGCTGCGGGGAATTGTC |
| Ana23S-753r | TGCAAAAGGTACGCTGTCAC | ||
| Wolbachia | 16S | W-SpecF | CATACCTATTCGAAGGGATAG |
| W-SpecR | AGCTTCGAGTGAA ACCAATTC | ||
| Species | Tested | Wolbachia Positive (%) |
|---|---|---|
| Anopheles gambiae s.l. | 171 | 0 (0) |
| Anopheles paludis | 211 | 1 (0.4%) |
| Anopheles nili | 2 | 0 (0) |
| Anopheles moucheti | 20 | 0 (0) |
| Culex (Lutzia) tigripes | 18 | 1 (5.55) |
| Culex duttoni | 43 | 0 (0) |
| Culex perfuscus | 116 | 4 (3.44) |
| Culex quinquefasciatus | 730 | 7 (0.95) |
| Culex univittatus | 11 | 0 (0) |
| Culex moucheti | 15 | 7 (46.67) |
| Culex wigglesworthi | 1 | 1 (100) |
| Culex sp. | 52 | 2 (3.84) |
| Aedes albopictus | 155 | 8 (5.16) |
| Aedes aegypti | 43 | 0 (0) |
| Aedes africanus | 4 | 0 (0) |
| Aedes simpsoni | 6 | 0 (0) |
| Aedes contigus | 32 | 1 (3.12) |
| Aedes sp. | 6 | 0 (0) |
| Coquillettidia spp. | 7 | 1 (14.28) |
| Mansonia africana | 42 | 0 (0) |
| Mansonia uniformis | 55 | 0 (0) |
| Total | 1740 | 33 (1.89) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bamou, R.; Diarra, A.Z.; Mayi, M.P.A.; Djiappi-Tchamen, B.; Antonio-Nkondjio, C.; Parola, P. Wolbachia Detection in Field-Collected Mosquitoes from Cameroon. Insects 2021, 12, 1133. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121133
Bamou R, Diarra AZ, Mayi MPA, Djiappi-Tchamen B, Antonio-Nkondjio C, Parola P. Wolbachia Detection in Field-Collected Mosquitoes from Cameroon. Insects. 2021; 12(12):1133. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121133
Chicago/Turabian StyleBamou, Roland, Adama Zan Diarra, Marie Paul Audrey Mayi, Borel Djiappi-Tchamen, Christophe Antonio-Nkondjio, and Philippe Parola. 2021. "Wolbachia Detection in Field-Collected Mosquitoes from Cameroon" Insects 12, no. 12: 1133. https://0-doi-org.brum.beds.ac.uk/10.3390/insects12121133