Thelytokous Reproduction of Onion Thrips, Thrips tabaci Lindeman 1889, Infesting Welsh Onion and Genetic Variation among Their Subpopulations


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
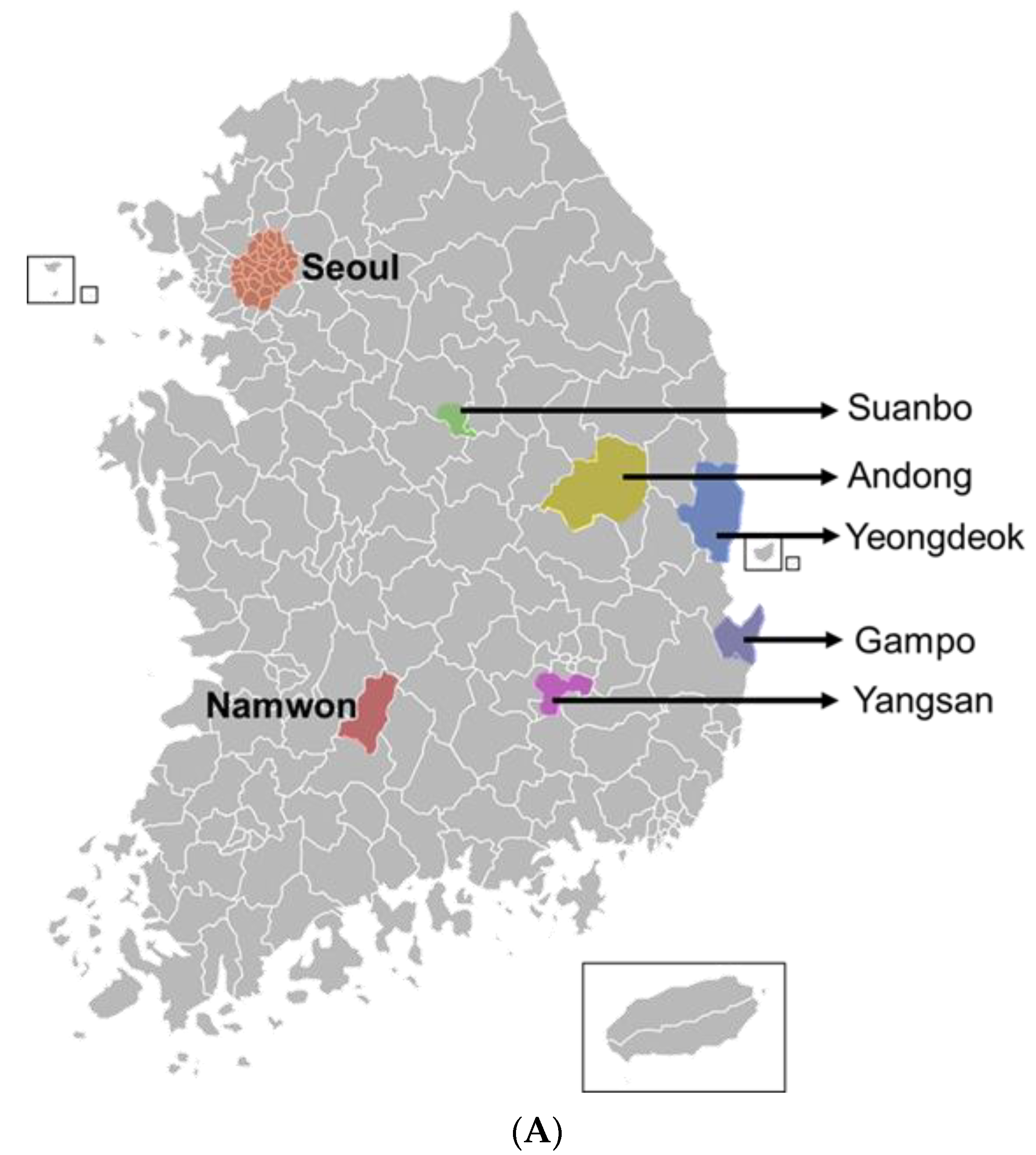
2.2. Field Monitoring of T. tabaci from Three Localities
2.3. Laboratory Test of Thelytokous Reproduction
2.4. Genomic DNA Extraction
2.5. PCR of Mitochondrial COI
2.6. Bioinformatics to Identify Thrips Species and Biotypes
2.7. Random Amplified Polymorphic DNA (RAPD) Analysis of Local Populations
2.8. Bioassay on Susceptibility Variation among Field Populations
2.9. Statistical Analysis
3. Results
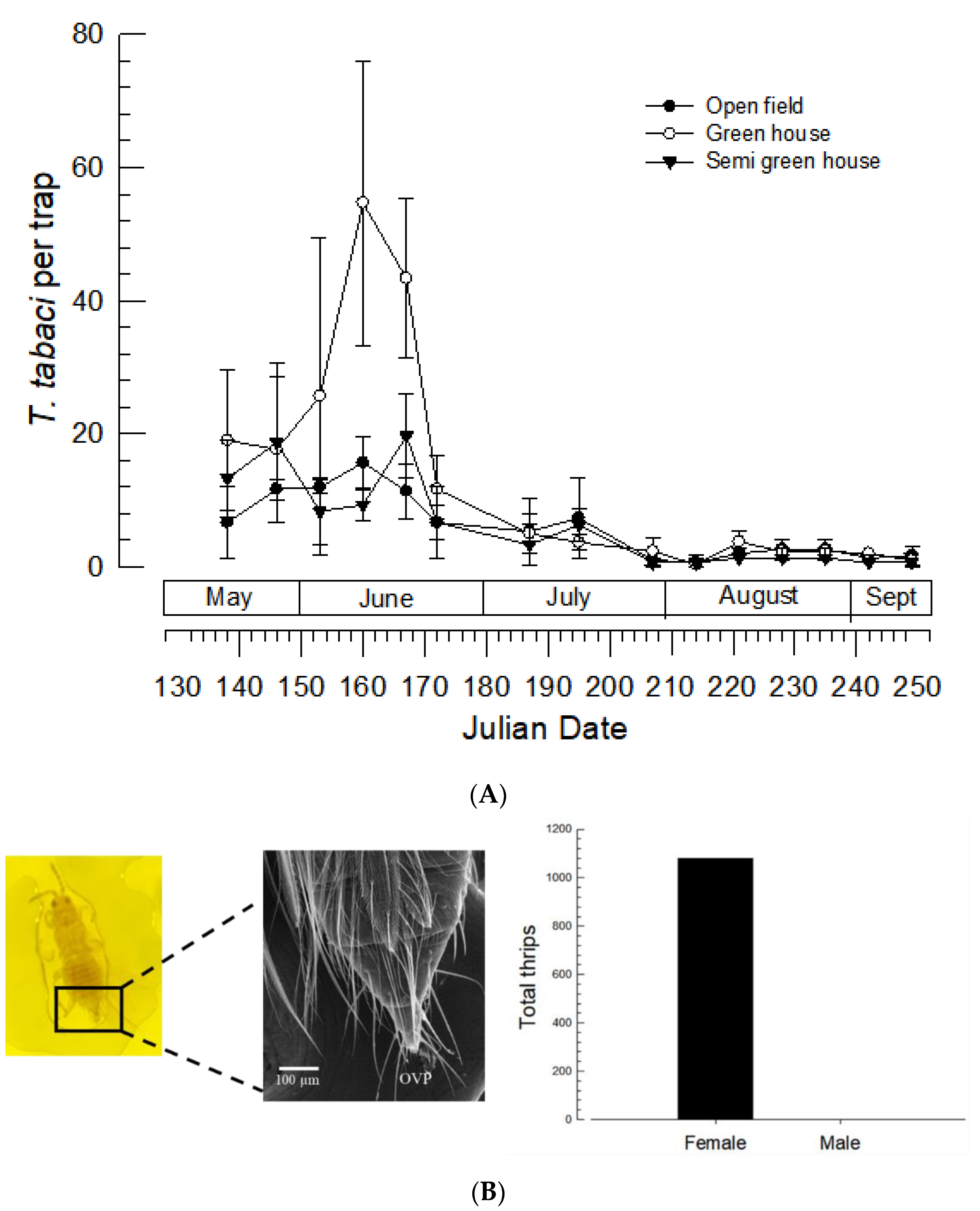
3.1. Seasonal Occurrence of T. tabaci in Welsh Onion Field
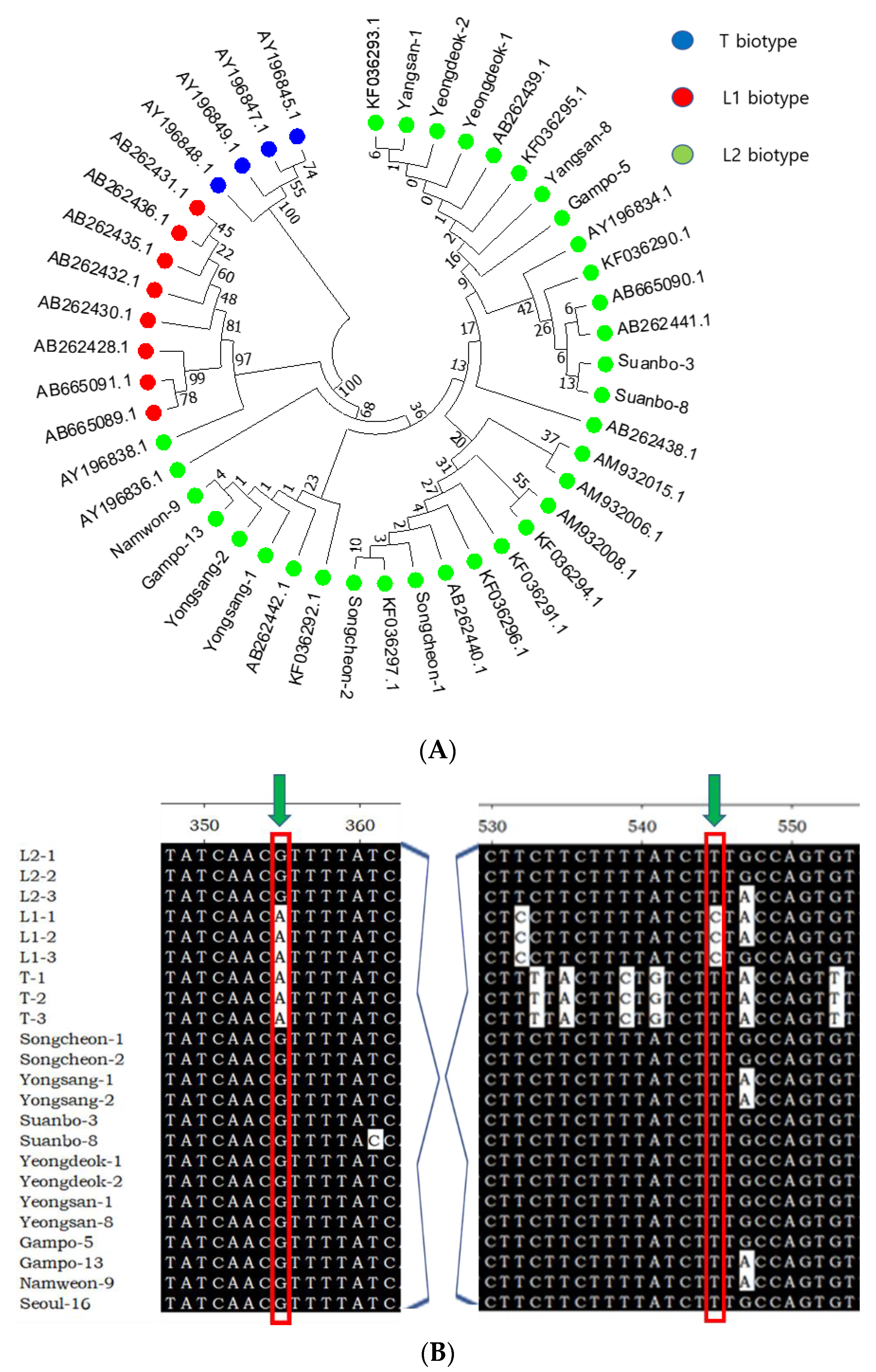
3.2. Thelytokous Reproduction and Its Genetic Characters
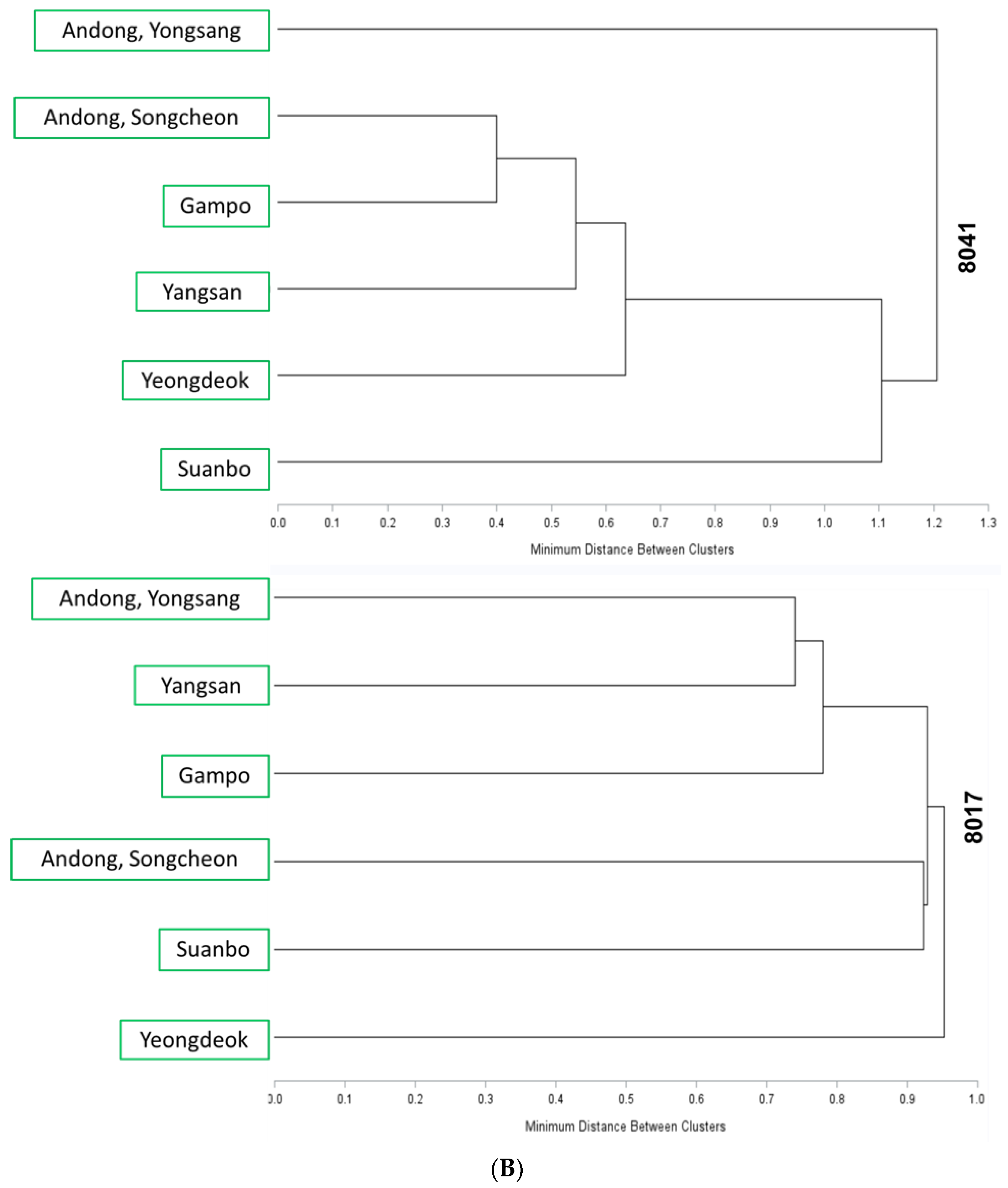
3.3. Genetic Diversity of Thelytokous Local Populations
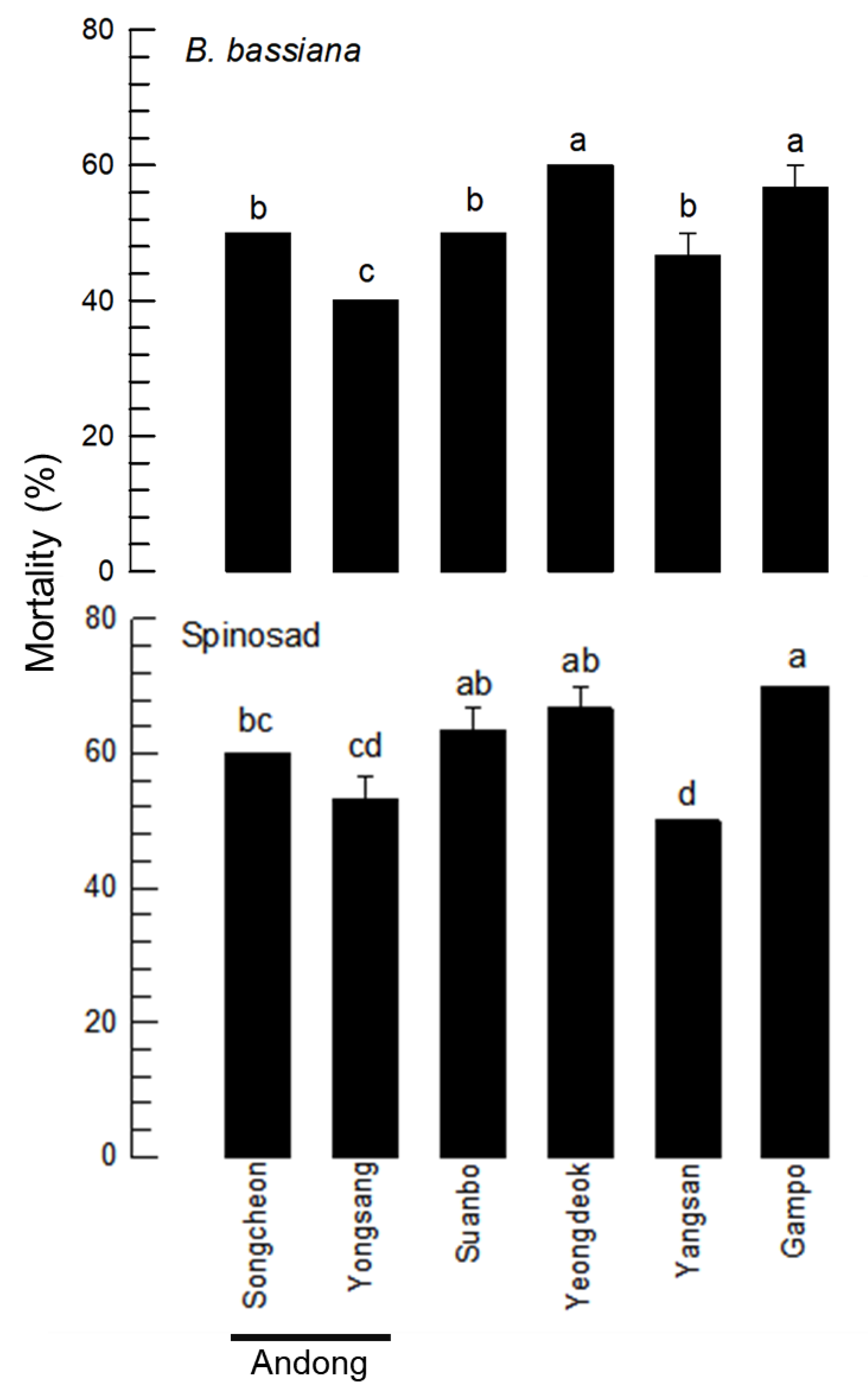
3.4. Variation in Insecticide Susceptibility of Local Populations—No Directional Population Subdivision
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Parrella, M.P.; Lewis, T. Integrated pest management (IPM) in field crops. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: New York, NY, USA, 1997; pp. 595–614. [Google Scholar]
- Dutta, B.; Barman, A.K.; Srinivasan, R.; Avci, U.; Ullman, D.E.; Langston, D.B.; Gitaitis, R.D. Transmission of Pantoea ananatis and P. agglomerans, causal agents of center rot of onion (Allium cepa), by onion thrips (Thrips tabaci) through feces. Phytopathology 2014, 104, 812–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grode, A.; Chen, S.; Walker, E.D.; Szendrei, Z. Onion thrips (Thysanoptera: Thripidae) feeding promotes infection by Pantoea ananatis in onion. J. Econ. Entomol. 2017, 110, 2301–2307. [Google Scholar] [CrossRef]
- Gent, D.H.; du Toit, L.J.; Fichtner, S.F.; Mohan, S.K.; Pappu, H.R.; Schwartz, H.F. Iris yellow spot virus: An emerging threat to onion bulb and seed production. Plant Dis. 2006, 90, 1468–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotenberg, D.; Jacobson, A.L.; Schneweis, D.J.; Whitfield, A.E. Thrips transmission of tospoviruses. Curr. Opin. Virol. 2015, 15, 80–89. [Google Scholar] [CrossRef]
- Diaz-Montano, J.; Fuchs, M.; Nault, B.A.; Fail, J.; Shelton, A.M. Onion thrips (Thysanoptera: Thripidae): A global pest of increasing concern in onion. J. Econ. Entomol. 2011, 104, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Moretti, E.A.; Harding, R.S.; Scott, J.G.; Nault, B.A. Monitoring onion thrips (Thysanoptera: Thripidae) susceptibility to spinetoram in New York onion fields. J. Econ. Entomol. 2019, 112, 1493–1497. [Google Scholar] [CrossRef]
- Zawirska, I. Untersuchungen über zwei biologische Typen von Thrips tabaci Lind. (Thysanoptera, Thripidae) in der VR Pole. Arch. Phytopathol. Plant Prot. 1976, 12, 411–422. [Google Scholar] [CrossRef]
- Brunner, P.C.; Chatzivassiliou, E.K.; Katis, N.I.; Frey, J.E. Host-associated genetic differentiation in Thrips tabaci (Insecta; Thysanoptera), as determined from mtDNA sequence data. Heredity 2004, 93, 364–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobson, A.L.; Nault, B.A.; Vargo, E.L.; Kennedy, G.G. Restricted gene flow among lineages of Thrips tabaci supports genetic divergence among cryptic species groups. PLoS ONE 2016, 11, e0163882. [Google Scholar]
- Toda, S.; Murai, T. Phylogenetic analysis based on mitochondrial COI gene sequences in Thrips tabaci Lindeman (Thysanoptera: Thripidae) in relation to reproductive forms and geographic distribution. Appl. Entomol. Zool. 2007, 42, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Kang, T.J.; Cho, M.R.; Kim, H.H.; Jeon, H.Y.; Kim, D.S. Economic injury level of Thrips tabaci (Thysanoptera: Thripidae) on welsh onions (Allium fistulosum L. var) in the early transplanting stage. Korean J. Appl. Entomol. 2011, 50, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.; Choi, D.; Kim, Y. Report on outbreak of the onion thrips, Thrips tabaci, infesting welsh onion during winter season. Korean J. Appl. Entomol. 2021, 60, 247–254. [Google Scholar]
- Reiter, D.; Péter, F.; Sojnóczki, K.; Kristóf, D.K.; József, F. Laboratory rearing of Thrips tabaci Lindeman: A review. Bodenkult. J. Fur Landwirtsch. Forsch. 2015, 66, 33–40. [Google Scholar]
- Marullo, R.; Bonsignore, C.P.; Vono, G. Thrips: A review of sampling methods in relation to their habitats. Bull. Insectol. 2021, 74, 241–251. [Google Scholar]
- Kim, C.; Choi, D.; Kang, J.H.; Ahmed, S.; Kil, E.J.; Kwon, G.; Lee, G.S.; Kim, Y. Thrips infesting hot pepper cultured in greenhouses and variation in gene sequences encoded in TSWV. Korean J. Appl. Entomol. 2021, 60, 387–401. [Google Scholar]
- Christopher, S.; Francesco, F.; Andrew, B.; Bernie, C.; Hong, L.; Paul, F. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar]
- Kim, Y.; Al Baki, M.; Kwon, G.; Kim, D.; Park, K.C.; Ahn, J. Discrimination of different generations of Zeugodacus scutellata using age grading technique and their local genetic variation. J. Asia Pac. Entomol. 2019, 22, 908–915. [Google Scholar] [CrossRef]
- SAS Institute, Inc. SAS/STAT User’s Guide; SAS Institute: Cary, NC, USA, 1989. [Google Scholar]
- Marullo, R.; Mercati, F.; Vono, G. DNA Barcoding: A reliable method for the identification of thrips species (Thysanoptera, Thripidae) collected on sticky traps in onion fields. Insects 2020, 11, 489. [Google Scholar] [CrossRef] [PubMed]
- Chatzivassiliou, E.K. Thrips tabaci: An ambiguous vector of TSWV in perspective. In Thrips and Tospoviruses, 7th International Symposium on Thysanoptera; Marullo, R., Mound, L., Eds.; Australian National Insect Collection: Canberra, Australia, 2002; pp. 69–75. [Google Scholar]
- Lattorff, H.M.; Moritz, R.F.; Crewe, R.M.; Solignac, M. Control of reproductive dominance by the thelytoky gene in honeybees. Biol. Lett. 2007, 3, 292–295. [Google Scholar] [CrossRef] [Green Version]
- Robert, F.; Lebreton, F.; Bougnoux, M.E.; Paugam, A.; Wassermann, D.; Schlotterer, M. Use of random amplified polymorphic DNA as a typing method for Candida albicans in epidemiological surveillance of a burn unit. J. Clin. Microbiol. 1995, 33, 2366–2371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhang, Z.; Zhang, J.; Huang, J.; Wang, L.; Li, Y.; Hafeez, M.; Lu, Y. Population genetic diversity and structure of Thrips tabaci (Thysanoptera: Thripidae) on Allium hosts in China, inferred from mitochondrial COI gene sequences. J. Econ. Entomol. 2020, 113, 1426–1435. [Google Scholar] [CrossRef] [PubMed]
- Masel, J. Genetic drift. Curr. Biol. 2011, 21, 837–838. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, A.L.; Booth, W.; Vargo, E.L.; Kennedy, G.G. Thrips tabaci population genetic structure and polyploidy in relation to competency as a vector of tomato spotted wilt virus. PLoS ONE 2013, 8, e54484. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mating Parents | Replication | Alive Adults | Total Progeny | Progeny/Parent | |
|---|---|---|---|---|---|
| Female | Male | ||||
| 1 | 30 | 13 | 37 | 5.4 ± 0.9 | 0 |
| 4 | 10 | 26 | 96 | 3.6 ± 0.3 | 0 |
| 10 | 5 | 25 | 75 | 3.1 ± 0.2 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, F.; Roy, M.C.; Kim, Y. Thelytokous Reproduction of Onion Thrips, Thrips tabaci Lindeman 1889, Infesting Welsh Onion and Genetic Variation among Their Subpopulations. Insects 2022, 13, 78. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010078
Khan F, Roy MC, Kim Y. Thelytokous Reproduction of Onion Thrips, Thrips tabaci Lindeman 1889, Infesting Welsh Onion and Genetic Variation among Their Subpopulations. Insects. 2022; 13(1):78. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010078
Chicago/Turabian StyleKhan, Falguni, Miltan Chandra Roy, and Yonggyun Kim. 2022. "Thelytokous Reproduction of Onion Thrips, Thrips tabaci Lindeman 1889, Infesting Welsh Onion and Genetic Variation among Their Subpopulations" Insects 13, no. 1: 78. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010078