
Nitrogen Acquisition Strategies Mediated by Insect Symbionts: A Review of Their Mechanisms, Methodologies, and Case Studies


{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Reaction Mechanisms of Nitrogen Acquisition Strategies Mediated by Symbionts
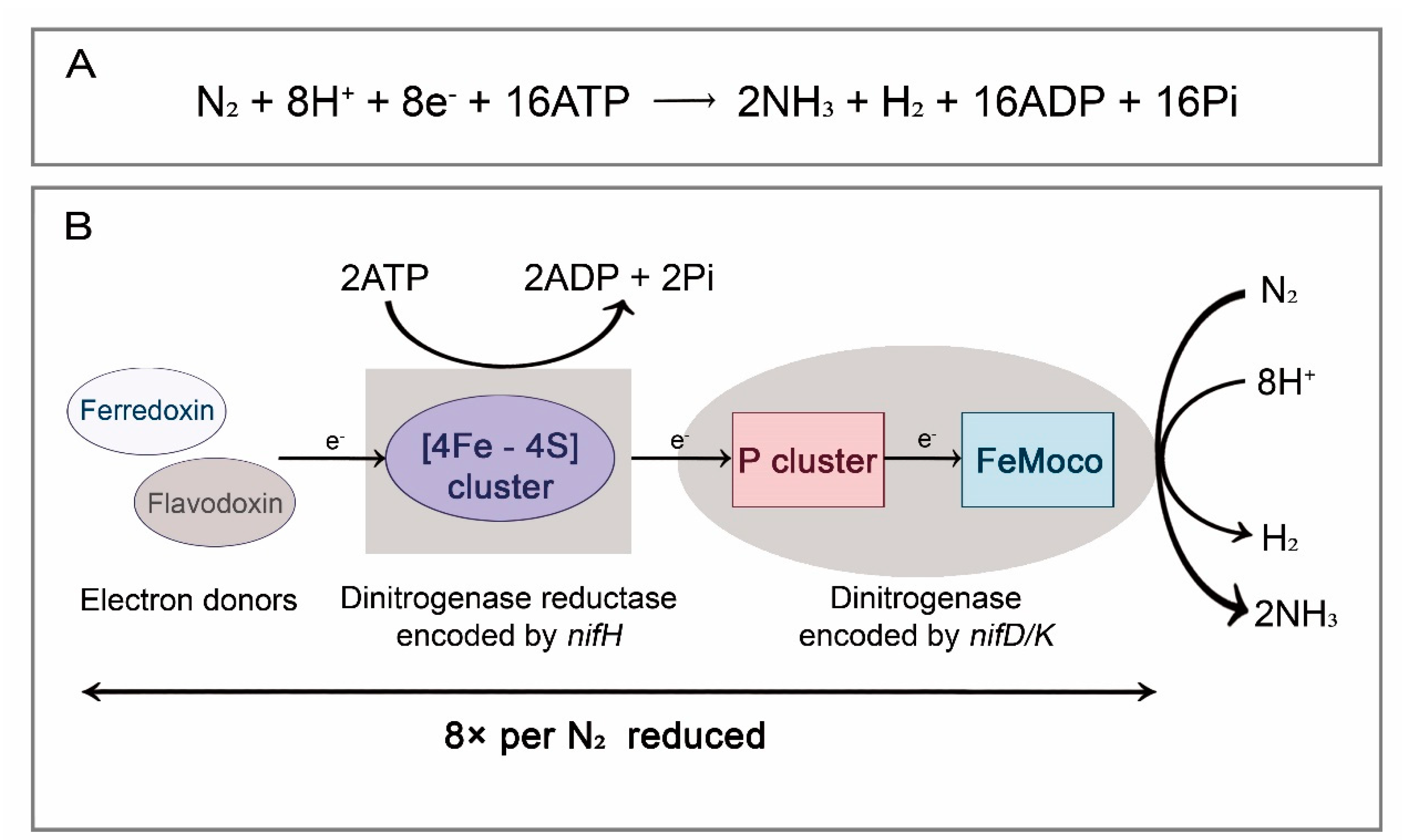
2.1. Biological Nitrogen Fixation
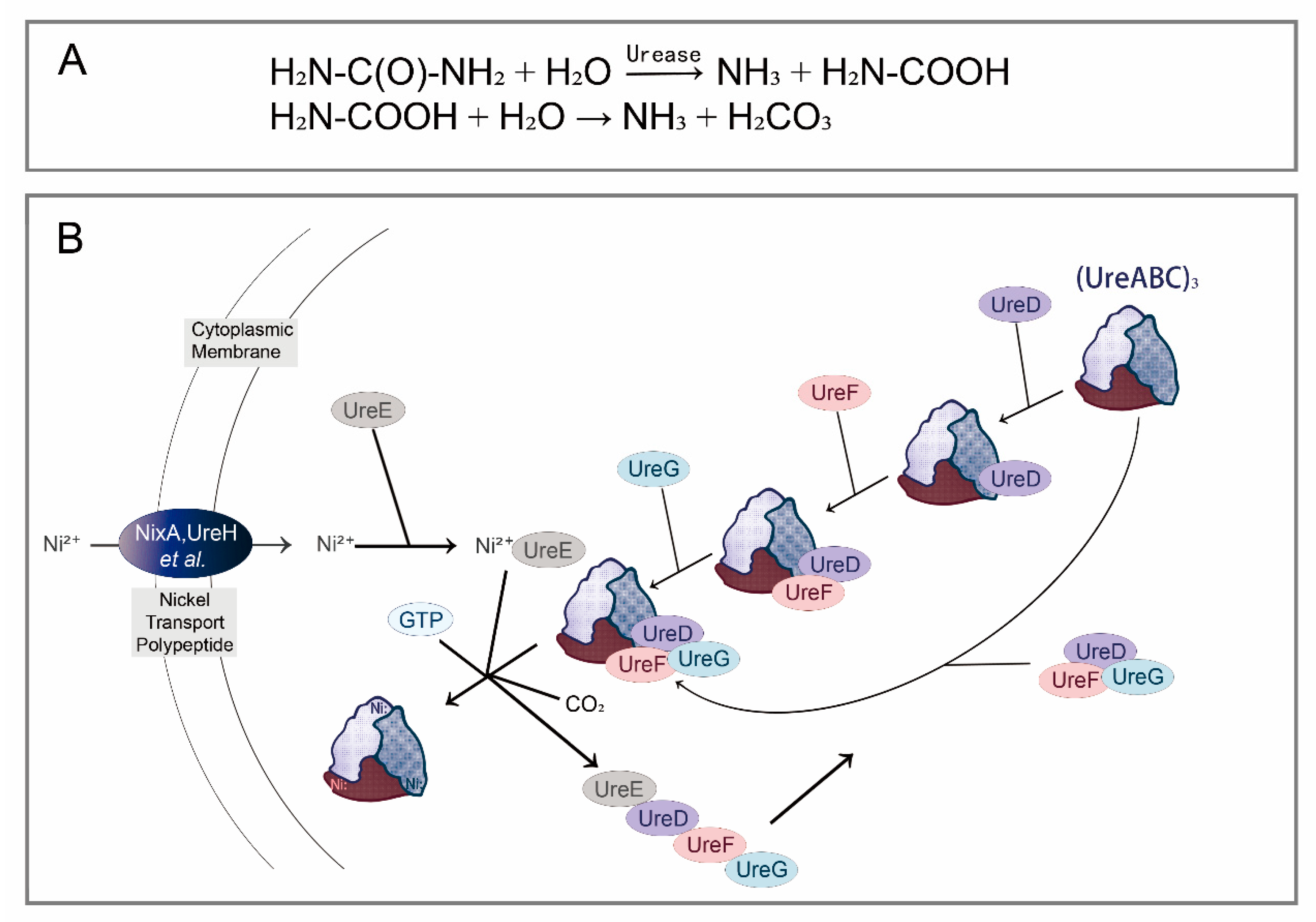
2.2. Nitrogenous Waste Recycling
3. Conventional Methods Related to BNF and NWR in Insects
3.1. Target Species Isolation and Functional Genes Identification
3.2. Enzyme Activity Determination
3.2.1. Determination of Nitrogenase Activity
3.2.2. Determination of Urease Activity
3.3. Isotopic Tracer Technique
3.4. High-Throughput Sequencing
4. The Various Applications of BNF and NWR Strategies in Insects
4.1. Application Cases of BNF Strategy in Insects
4.2. Application Cases of NWR Strategy in Insects
4.2.1. Blattaria
4.2.2. Coleoptera
4.2.3. Diptera
4.2.4. Hemiptera
4.2.5. Hymenoptera
5. Discussion
6. Future Perspectives
- Existing research about BNF and NWR strategies mediated by insect symbionts is incomplete and very limited. The priority level (which strategy is more suitable for given insects), occurrence rules (which category of insects are more dependent on BNF or NWR), and characteristics (what is the critical factor for strategies working) of these two strategies in insects have not yet been revealed;
- The extent of symbionts’ contribution to nitrogen provisioning in insects remains to be largely unknown. Specifically, how much nitrogen do insects fix through BNF or NWR? How much does this part of nitrogen nutrition account for the total insect demand? Addressing these questions will help us understand the significance of BNF and NWR during the coevolution between insects and symbionts;
- Could the functional symbionts involved in BNF and NWR be exploited in mass rearing insects for production of bioprotein or empowering sterile insect technique (SIT)? The purpose is similar with the feeding pattern of “urea + ureolytic bacteria” in ruminants breeding. This pattern will achieve significant cost savings by transferring atmospheric nitrogen or nitrogenous waste into EAA/protein.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hansen, A.K.; Pers, D.; Russell, J.A. Symbiotic Solutions to Nitrogen Limitation and Amino Acid Imbalance in Insect Diets. In Mechanisms Underlying Microbial Symbiosis; Oliver, K.M., Russell, J.A., Eds.; Academic Press Ltd.: London, UK; Elsevier Science Ltd.: London, UK, 2020; Volume 58, pp. 161–205. ISBN 9780081029879. [Google Scholar]
- Sudakaran, S.; Kost, C.; Kaltenpoth, M. Symbiont Acquisition and Replacement as a Source of Ecological Innovation. Trends Microbiol. 2017, 25, 375–390. [Google Scholar] [CrossRef]
- Hansen, A.K.; Moran, N.A. The Impact of Microbial Symbionts on Host Plant Utilization by Herbivorous Insects. Mol. Ecol. 2014, 23, 1473–1496. [Google Scholar] [CrossRef]
- Mattson, W.J. Herbivory in Relation to Plant Nitrogen-Content. Annu. Rev. Ecol. Syst. 1980, 11, 119–161. [Google Scholar] [CrossRef]
- Bar-Shmuel, N.; Behar, A.; Segoli, M. What Do We Know About Biological Nitrogen Fixation in Insects? Evidence and Implications for the Insect and the Ecosystem. Insect Sci. 2020, 27, 392–403. [Google Scholar] [CrossRef]
- Douglas, A.E. The Microbial Dimension in Insect Nutritional Ecology. Funct. Ecol. 2009, 23, 38–47. [Google Scholar] [CrossRef]
- Douglas, A.E. Multiorganismal Insects: Diversity and Function of Resident Microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Chiu, C.I.; Ou, J.H.; Chen, C.Y.; Li, H.F. Fungal Nutrition Allocation Enhances Mutualism with Fungus-Growing Termite. Fungal Ecol. 2019, 41, 92–100. [Google Scholar] [CrossRef]
- Nardi, J.B.; Mackie, R.I.; Dawson, J.O. Could Microbial Symbionts of Arthropod Guts Contribute Significantly to Nitrogen Fixation in Terrestrial Ecosystems? J. Insect Physiol. 2002, 48, 751–763. [Google Scholar] [CrossRef]
- Dixon, R.; Kahn, D. Genetic Regulation of Biological Nitrogen Fixation. Nat. Rev. Microbiol. 2004, 2, 621–631. [Google Scholar] [CrossRef]
- Kneip, C.; Lockhart, P.; Voss, C.; Maier, U.G. Nitrogen Fixation in Eukaryotes—New Models for Symbiosis. BMC Evol. Biol. 2007, 7, 55. [Google Scholar] [CrossRef] [Green Version]
- Mus, F.; Alleman, A.B.; Pence, N.; Seefeldt, L.C.; Peters, J.W. Exploring the Alternatives of Biological Nitrogen Fixation. Metallomics 2018, 10, 523–538. [Google Scholar] [CrossRef]
- Newton, W.E. Recent Advances in Understanding Nitrogenases and How They Work. In Biological Nitrogen Fixation; DeBruijn, F.J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; Volume 1, pp. 7–20. ISBN 9781118637043(H)/9781119053095(P). [Google Scholar]
- Anbar, A.D.; Knoll, A.H. Proterozoic Ocean Chemistry and Evolution: A Bioinorganic Bridge? Science 2002, 297, 1137–1142. [Google Scholar] [CrossRef] [Green Version]
- Macdonald, S.J.; Lin, G.G.; Russell, C.W.; Thomas, G.H.; Douglas, A.E. The Central Role of the Host Cell in Symbiotic Nitrogen Metabolism. Proc. R. Soc. B Biol. Sci. 2012, 279, 2965–2973. [Google Scholar] [CrossRef] [Green Version]
- Thong-On, A.; Suzuki, K.; Noda, S.; Inoue, J.; Kajiwara, S.; Ohkuma, M. Isolation and Characterization of Anaerobic Bacteria for Symbiotic Recycling of Uric Acid Nitrogen in the Gut of Various Termites. Microbes. Environ. 2012, 27, 186–192. [Google Scholar] [CrossRef] [Green Version]
- Fujii, T.; Banno, Y. Identification of a Novel Function of the Silkworm Integument in Nitrogen Metabolism: Uric Acid Is Synthesized within the Epidermal Cells in B. Mori. Insect Biochem. Molec. 2019, 105, 43–50. [Google Scholar] [CrossRef]
- Kashima, T.; Nakamura, T.; Tojo, S. Uric Acid Recycling in the Shield Bug, Parastrachia Japonensis (Hemiptera: Parastrachiidae), During Diapause. J. Insect Physiol. 2006, 52, 816–825. [Google Scholar] [CrossRef]
- Lei, C.L.; Rong, X.L. General Entomology, 1st ed.; China Agriculture Press Co., Ltd.: Beijing, China, 2003; Chapter 21; pp. 237–239. ISBN 9787109083806. [Google Scholar]
- Hu, Y.; Sanders, J.G.; Lukasik, P.; D’Amelio, C.L.; Millar, J.S.; Vann, D.R.; Lan, Y.M.; Newton, J.A.; Schotanus, M.; Kronauer, D.J.C.; et al. Herbivorous Turtle Ants Obtain Essential Nutrients from a Conserved Nitrogen-Recycling Gut Microbiome. Nat. Commun. 2018, 9, 964. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.K.; Aschenbach, J.R. Ureases in the Gastrointestinal Tracts of Ruminant and Monogastric Animals and Their Implication in Urea-N/Ammonia Metabolism: A Review. J. Adv. Res. 2018, 13, 39–50. [Google Scholar] [CrossRef]
- Yuen, M.H.; Fong, Y.H.; Nim, Y.S.; Lau, P.H.; Wong, K.-B. Structural Insights into How GTP-Dependent Conformational Changes in a Metallochaperone UreG Facilitate Urease Maturation. Proc. Natl. Acad. Sci. USA 2017, 114, 10890–10898. [Google Scholar] [CrossRef] [Green Version]
- Hailemariam, S.; Zhao, S.; He, Y.; Wang, J. Urea Transport and Hydrolysis in the Rumen: A Review. Anim. Nutr. 2021, 7, 989–996. [Google Scholar] [CrossRef]
- Jin, D.; Zhao, S.G.; Zheng, N.; Beckers, Y.; Wang, J.Q. Urea Metabolism and Regulation by Rumen Bacterial Urease in Ruminants —A Review. Ann. Anim. Sci. 2018, 18, 303–318. [Google Scholar] [CrossRef] [Green Version]
- Kertz, A.F. Review: Urea Feeding to Dairy Cattle: A Historical Perspective and Review. Prof. Anim. Sci. 2010, 26, 257–272. [Google Scholar] [CrossRef]
- Patra, A.K. Urea/Ammonia Metabolism in the Rumen and Toxicity in Ruminants. In Rumen Microbiology: From Evolution to Revolution; Puniya, A., Singh, R., Kamra, D., Eds.; Springer: New Delhi, India, 2015; pp. 329–341. ISBN 9788132224013. [Google Scholar]
- He, Y.; Zhang, X.; Li, M.; Zheng, N.; Zhao, S.; Wang, J. Coptisine: A Natural Plant Inhibitor of Ruminal Bacterial Urease Screened by Molecular Docking. Sci. Total Environ. 2021, 808, 151946. [Google Scholar] [CrossRef]
- Ben-Yosef, M.; Pasternak, Z.; Jurkevitch, E.; Yuval, B. Symbiotic Bacteria Enable Olive Flies (Bactrocera Oleae) to Exploit Intractable Sources of Nitrogen. J. Evol. Biol. 2014, 27, 2695–2705. [Google Scholar] [CrossRef]
- Isibika, A.; Vinneras, B.; Kibazohi, O.; Zurbrugg, C.; Lalander, C. Pre-Treatment of Banana Peel to Improve Composting by Black Soldier Fly (Hermetia Illucens (L.), Diptera: Stratiomyidae) Larvae. Waste Manag. 2019, 100, 151–160. [Google Scholar] [CrossRef]
- Ayayee, P.; Rosa, C.; Ferry, J.G.; Felton, G.; Saunders, M.; Hoover, K. Gut Microbes Contribute to Nitrogen Provisioning in a Wood-Feeding Cerambycid. Environ. Entomol. 2014, 43, 903–912. [Google Scholar] [CrossRef]
- Reid, N.M.; Addison, S.L.; Macdonald, L.J.; Lloyd-Jones, G. Biodiversity of Active and Inactive Bacteria in the Gut Flora of Wood-Feeding Huhu Beetle Larvae (Prionoplus Reticularis). Appl. Environ. Microb. 2011, 77, 7000–7006. [Google Scholar] [CrossRef] [Green Version]
- Rizzi, A.; Crotti, E.; Borruso, L.; Jucker, C.; Lupi, D.; Colombo, M.; Daffonchio, D. Characterization of the Bacterial Community Associated with Larvae and Adults of Anoplophora Chinensis Collected in Italy by Culture and Culture-Independent Methods. BioMed Res. Int. 2013, 2013, 420287. [Google Scholar] [CrossRef] [Green Version]
- Farnelid, H.; Harder, J.; Bentzon-Tilia, M.; Riemann, L. Isolation of Heterotrophic Diazotrophic Bacteria from Estuarine Surface Waters. Environ. Microbiol. 2014, 16, 3072–3082. [Google Scholar] [CrossRef]
- de Bruijn, F.J. Biological Nitrogen Fixation. In Principles of Plant-Microbe Interactions: Microbes for Sustainable Agriculture; Lugtenberg, B., Ed.; Springer: Cham Switzerland, 2015; pp. 215–224. ISBN 9783319085753. [Google Scholar]
- Dos Santos, P.C.; Fang, Z.; Mason, S.W.; Setubal, J.C.; Dixon, R. Distribution of Nitrogen Fixation and Nitrogenase-Like Sequences Amongst Microbial Genomes. BMC Genom. 2012, 13, 547. [Google Scholar] [CrossRef] [Green Version]
- Bellenger, J.P.; Darnajoux, R.; Zhang, X.; Kraepiel, A.M.L. Biological Nitrogen Fixation by Alternative Nitrogenases in Terrestrial Ecosystems: A Review. Biogeochemistry 2020, 149, 53–73. [Google Scholar] [CrossRef]
- Bishai, W.R.; Timmins, G.S. Potential for Breath Test Diagnosis of Urease Positive Pathogens in Lung Infections. J. Breath Res. 2019, 13, 032002. [Google Scholar] [CrossRef]
- Carter, E.L.; Flugga, N.; Boer, J.L.; Mulrooney, S.B.; Hausinger, R.P. Interplay of Metal Ions and Urease. Metallomics 2009, 1, 207–221. [Google Scholar] [CrossRef] [Green Version]
- Maroney, M.J.; Ciurli, S. Nickel as a Virulence Factor in the Class I Bacterial Carcinogen. Helicobacter. Pylori. Semin. Cancer Biol. 2021, 76, 143–155. [Google Scholar] [CrossRef]
- Soper, F.M.; Simon, C.; Jauss, V. Measuring Nitrogen Fixation by the Acetylene Reduction Assay (Ara): Is 3 the Magic Ratio? Biogeochemistry 2021, 152, 345–351. [Google Scholar] [CrossRef]
- Saiz, E.; Sgouridis, F.; Drijfhout, F.P.; Ullah, S. Biological Nitrogen Fixation in Peatlands: Comparison between Acetylene Reduction Assay and 15N2 Assimilation Methods. Soil Biol. Biochem. 2019, 131, 157–165. [Google Scholar] [CrossRef]
- Behar, A.; Yuval, B.; Jurkevitch, E. Enterobacteria-Mediated Nitrogen Fixation in Natural Populations of the Fruit Fly Ceratitis Capitata. Mol. Ecol. 2005, 14, 2637–2643. [Google Scholar] [CrossRef]
- Kuranouchi, T.; Nakamura, T.; Shimamura, S.; Kojima, H.; Goka, K.; Okabe, K.; Mochizuki, A. Nitrogen Fixation in the Stag Beetle, Dorcus (Macrodorcus) Rectus (Motschulsky) (Col., Lucanidae). J. Appl. Entomol. 2006, 130, 471–472. [Google Scholar] [CrossRef]
- Young, G.M.; Amid, D.; Miller, V.L. A Bifunctional Urease Enhances Survival of Pathogenic Yersinia Enterocolitica and Morganella Morganii at Low Ph. J. Bacteriol. 1996, 178, 6487–6495. [Google Scholar] [CrossRef] [Green Version]
- Nieckarz, M.; Kaczor, P.; Jaworska, K.; Raczkowska, A.; Brzostek, K. Urease Expression in Pathogenic Yersinia Enterocolitica Strains of Bio-Serotypes 2/O:9 and 1b/O:8 Is Differentially Regulated by the Ompr Regulator. Front. Microbiol. 2020, 11, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Y.; Yan, Z.K.; Wang, Z.K.; Batool, M.; El-Badri, A.M.; Bai, F.; Li, Z.; Wang, B.; Zhou, G.S.; Kuai, J. Subsoil Tillage Promotes Root and Shoot Growth of Rapeseed in Paddy Fields and Dryland in Yangtze River Basin Soils. Eur. J. Agron. 2021, 130, 126351. [Google Scholar] [CrossRef]
- Ceja-Navarro, J.A.; Nguyen, N.H.; Karaoz, U.; Gross, S.R.; Herman, D.J.; Andersen, G.L.; Bruns, T.D.; Pett-Ridge, J.; Blackwell, M.; Brodie, E.L. Compartmentalized Microbial Composition, Oxygen Gradients and Nitrogen Fixation in the Gut of Odontotaenius Disjunctus. ISME J. 2014, 8, 6–18. [Google Scholar] [CrossRef] [Green Version]
- Potrikus, C.J.; Breznak, J.A. Gut Bacteria Recycle Uric-Acid Nitrogen in Termites—A Strategy for Nutrient Conservation. Proc. Natl. Acad. Sci. USA 1981, 78, 4601–4605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso-Pernas, P.; Bartram, S.; Arias-Cordero, E.M.; Novoselov, A.L.; Halty-deLeon, L.; Shao, Y.Q.; Boland, W. In Vivo Isotopic Labeling of Symbiotic Bacteria Involved in Cellulose Degradation and Nitrogen Recycling within the Gut of the Forest Cockchafer (Melolontha Hippocastani). Front. Microbiol. 2018, 9, 124462. [Google Scholar] [CrossRef] [PubMed]
- Pinto-Tomas, A.A.; Anderson, M.A.; Suen, G.; Stevenson, D.M.; Chu, F.S.T.; Cleland, W.W.; Weimer, P.J.; Currie, C.R. Symbiotic Nitrogen Fixation in the Fungus Gardens of Leaf-Cutter Ants. Science 2009, 326, 1120–1123. [Google Scholar] [CrossRef]
- Douglas, A.E. Microbial Brokers of Insect-Plant Interactions Revisited. J. Chem. Ecol. 2013, 39, 952–961. [Google Scholar] [CrossRef] [Green Version]
- Herbold, C.W.; Pelikan, C.; Kuzyk, O.; Hausmann, B.; Angel, R.; Berry, D.; Loy, A. A Flexible and Economical Barcoding Approach for Highly Multiplexed Amplicon Sequencing of Diverse Target Genes. Front. Microbiol. 2015, 6, 731. [Google Scholar] [CrossRef] [Green Version]
- Niu, S.Y.; Yang, J.Y.; McDermaid, A.; Zhao, J.; Kang, Y.; Ma, Q. Bioinformatics Tools for Quantitative and Functional Metagenome and Metatranscriptome Data Analysis in Microbes. Brief. Bioinform. 2018, 19, 1415–1429. [Google Scholar] [CrossRef] [PubMed]
- Sabree, Z.L.; Kambhampati, S.; Moran, N.A. Nitrogen Recycling and Nutritional Provisioning by Blattabacterium, the Cockroach Endosymbiont. Proc. Natl. Acad. Sci. USA 2009, 106, 19521–19526. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.P.; Sanders, J.G.; Byrne, M.J.; Pierce, N.E. Gut Microbiota of Dung Beetles Correspond to Dietary Specializations of Adults and Larvae. Mol. Ecol. 2016, 25, 6092–6106. [Google Scholar] [CrossRef] [PubMed]
- de Leon, A.V.P.; Ormeno-Orrillo, E.; Ramirez-Puebla, S.T.; Rosenblueth, M.; Esposti, M.D.; Martinez-Romero, J.; Martinez-Romero, E. Candidatus Dactylopiibacterium Carminicum, a Nitrogen-Fixing Symbiont of Dactylopius Cochineal Insects (Hemiptera: Coccoidea: Dactylopiidae). Genome Biol. Evol. 2017, 9, 2237–2250. [Google Scholar] [CrossRef] [Green Version]
- Ulyshen, M.D. Insect-Mediated Nitrogen Dynamics in Decomposing Wood. Ecol. Entomol. 2015, 40, 97–112. [Google Scholar] [CrossRef]
- Aylward, F.O.; Burnum, K.E.; Scott, J.J.; Suen, G.; Tringe, S.G.; Adams, S.M.; Barry, K.W.; Nicora, C.D.; Piehowski, P.D.; Purvine, S.O.; et al. Metagenomic and Metaproteomic Insights into Bacterial Communities in Leaf-Cutter Ant Fungus Gardens. ISME J. 2012, 6, 1688–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patino-Navarrete, R.; Piulachs, M.D.; Belles, X.; Moya, A.; Latorre, A.; Pereto, J. The Cockroach Blattella Germanica Obtains Nitrogen from Uric Acid through a Metabolic Pathway Shared with Its Bacterial Endosymbiont. Biol. Lett. 2014, 10, 20140407. [Google Scholar] [CrossRef] [Green Version]
- Weihrauch, D.; O’Donnell, M.J. Links between Osmoregulation and Nitrogen-Excretion in Insects and Crustaceans. Integr. Comp. Biol. 2015, 55, 816–829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa-Leonardo, A.M.; Laranjo, L.T.; Janei, V.; Haifig, I. The Fat Body of Termites: Functions and Stored Materials. J. Insect Physiol. 2013, 59, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Scharf, M.E. Challenges and Physiological Implications of Wood Feeding in Termites. Curr. Opin. Insect Sci. 2020, 41, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Kinjo, Y.; Bourguignon, T.; Tong, K.J.; Kuwahara, H.; Lim, S.J.; Yoon, K.B.; Shigenobu, S.; Park, Y.C.; Nalepa, C.A.; Hongoh, Y.; et al. Parallel and Gradual Genome Erosion in the Blattabacterium Endosymbionts of Mastotermes Darwiniensis and Cryptocercus Wood Roaches. Genome. Biol. Evol. 2018, 10, 1622–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabree, Z.L.; Huang, C.Y.; Arakawa, G.; Tokuda, G.; Lo, N.; Watanabe, H.; Moran, N.A. Genome Shrinkage and Loss of Nutrient-Providing Potential in the Obligate Symbiont of the Primitive Termite Mastotermes Darwiniensis. Appl. Environ. Microb. 2012, 78, 204–210. [Google Scholar] [CrossRef] [Green Version]
- Morales-Jimenez, J.; de Leon, A.V.P.; Garcia-Dominguez, A.; Martinez-Romero, E.; Zuniga, G.; Hernandez-Rodriguez, C. Nitrogen-Fixing and Uricolytic Bacteria Associated with the Gut of Dendroctonus Rhizophagus and Dendroctonus Valens (Curculionidae: Scolytinae). Microb. Ecol. 2013, 66, 200–210. [Google Scholar] [CrossRef]
- Lauzon, C.R.; Sjogren, R.E.; Prokopy, R.J. Enzymatic Capabilities of Bacteria Associated with Apple Maggot Flies: A Postulated Role in Attraction. J. Chem. Ecol. 2000, 26, 953–967. [Google Scholar] [CrossRef]
- Robacker, D.C.; Lauzon, C.R. Purine Metabolizing Capability of Enterobacter Agglomerans Affects Volatiles Production and Attractiveness to Mexican Fruit Fly. J. Chem. Ecol. 2002, 28, 1549–1563. [Google Scholar] [CrossRef] [PubMed]
- Baumann, P. Biology of Bacteriocyte-Associated Endosymbionts of Plant Sap-Sucking Insects. Annu. Rev. Microbiol. 2005, 59, 155–189. [Google Scholar] [CrossRef]
- Douglas, A.E. How Multi- Partner Endosymbioses Function. Nat. Rev. Microbiol. 2016, 14, 731–743. [Google Scholar] [CrossRef] [PubMed]
- Ankrah, N.Y.D.; Chouaia, B.; Douglas, A.E. The Cost of Metabolic Interactions in Symbioses between Insects and Bacteria with Reduced Genomes. Mbio 2018, 9, e01433-18. [Google Scholar] [CrossRef] [Green Version]
- Hongoh, Y.; Sasaki, T.; Ishikawa, H. Cloning, Sequence Analysis and Expression in Escherichia Coli of the Gene Encoding a Uricase from the Yeast-Like Symbiont of the Brown Planthopper, Nilaparvata Lugens. Insect Biochem. Mol. 2000, 30, 173–182. [Google Scholar] [CrossRef]
- Xue, J.; Zhou, X.; Zhang, C.X.; Yu, L.L.; Fan, H.W.; Wang, Z.; Xu, H.J.; Xi, Y.; Zhu, Z.R.; Zhou, W.W.; et al. Genomes of the Rice Pest Brown Planthopper and Its Endosymbionts Reveal Complex Complementary Contributions for Host Adaptation. Genome Biol. 2014, 15, 521. [Google Scholar] [CrossRef] [Green Version]
- de Leon, A.V.P.; Sanchez-Flores, A.; Rosenblueth, M.; Martinez-Romero, E. Fungal Community Associated with Dactylopius (Hemiptera: Coccoidea: Dactylopiidae) and Its Role in Uric Acid Metabolism. Front. Microbiol. 2016, 7, 954. [Google Scholar] [CrossRef]
- Hansen, A.K.; Moran, N.A. Aphid Genome Expression Reveals Host-Symbiont Cooperation in the Production of Amino Acids. Proc. Natl. Acad. Sci. USA 2011, 108, 2849–2854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shetty, P.S. Gustatory Preferences of Ants (Camponotus-Compressus) for Urea and Sugars. Experientia 1982, 38, 259–260. [Google Scholar] [CrossRef]
- Feldhaar, H.; Straka, J.; Krischke, M.; Berthold, K.; Stoll, S.; Mueller, M.J.; Gross, R. Nutritional Upgrading for Omnivorous Carpenter Ants by the Endosymbiont Blochmannia. BMC Biol. 2007, 5, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zientz, E.; Beyaert, N.; Gross, R.; Feldhaar, H. Relevance of the Endosymbiosis of Blochmannia Floridanus and Carpenter Ants at Different Stages of the Life Cycle of the Host. Appl. Environ. Microb. 2006, 72, 6027–6033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bisch, G.; Neuvonen, M.M.; Pierce, N.E.; Russell, J.A.; Koga, R.; Sanders, J.G.; Lukasik, P.; Andersson, S.G.E. Genome Evolution of Bartonellaceae Symbionts of Ants at the Opposite Ends of the Trophic Scale. Genome. Biol. Evol. 2018, 10, 1687–1704. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.C.; Davidson, D.W. Nutritional and Functional Biology of Exudate-Feeding Ants. Entomol. Exp. Appl. 2006, 118, 1–10. [Google Scholar] [CrossRef]
- Scaraffia, P.Y.; Zhang, Q.F.; Thorson, K.; Wysocki, V.H.; Miesfeld, R.L. Differential Ammonia Metabolism in Aedes Aegypti Fat Body and Midgut Tissues. J. Insect Physiol. 2010, 56, 1040–1049. [Google Scholar] [CrossRef] [Green Version]
- Weihrauch, D.; O’Donnell, M.J. Mechanisms of Nitrogen Excretion in Insects. Curr. Opin. Insect Sci. 2021, 47, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Winans, N.J.; Walter, A.; Chouaia, B.; Chaston, J.M.; Douglas, A.E.; Newell, P.D. A Genomic Investigation of Ecological Differentiation between Free-Living and Drosophila-Associated Bacteria. Mol. Ecol. 2017, 26, 4536–4550. [Google Scholar] [CrossRef]
- Baumann, A.; Skaljac, M.; Lehmann, R.; Vilcinskas, A.; Franta, Z. Urate Oxidase Produced by Lucilia Sericata Medical Maggots Is Localized in Malpighian Tubes and Facilitates Allantoin Production. Insect Biochem. Mol. 2017, 83, 44–53. [Google Scholar] [CrossRef]
- Tang, L.M.; Yang, D.H.; Wang, Y.H.; Yang, X.; Chen, K.; Luo, X.Y.; Xu, J.; Liu, Y.J.; Tang, Z.; Zhang, Q.Q.; et al. 5’-Nucleotidase Plays a Key Role in Uric Acid Metabolism of Bombyx Mori. Cells 2021, 10, 2243. [Google Scholar] [CrossRef]
- Hirayama, C.; Sugimura, M.; Saito, H.; Nakamura, M. Host Plant Urease in the Hemolymph of the Silkworm, Bombyx Mori. J. Insect Physiol. 2000, 46, 1415–1421. [Google Scholar] [CrossRef]
- Hongoh, Y.; Sharma, V.K.; Prakash, T.; Noda, S.; Toh, H.; Taylor, T.D.; Kudo, T.; Sakaki, Y.; Toyoda, A.; Hattori, M.; et al. Genome of an Endosymbiont Coupling N2 Fixation to Cellulolysis within Protist Cells in Termite Gut. Science 2008, 322, 1108–1109. [Google Scholar] [CrossRef]
- Jahnes, B.C.; Sabree, Z.L. Nutritional Symbiosis and Ecology of Host -Gut Microbe Systems in the Blattodea. Curr. Opin. Insect Sci. 2020, 39, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Soumare, A.; Diedhiou, A.G.; Thuita, M.; Hafidi, M.; Ouhdouch, Y.; Gopalakrishnan, S.; Kouisni, L. Exploiting Biological Nitrogen Fixation: A Route Towards a Sustainable Agriculture. Plants 2020, 9, 1011. [Google Scholar] [CrossRef] [PubMed]
- Alleman, A.B.; Mus, F.; Peters, J.W. Metabolic Model of the Nitrogen-Fixing Obligate Aerobe Azotobacter Vinelandii Predicts Its Adaptation to Oxygen Concentration and Metal Availability. Mbio 2021, 12, e0259321. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, P.C.; Dean, D.R. Co-Ordination and Fine-Tuning of Nitrogen Fixation in Azotobacter Vinelandii. Mol. Microbiol. 2011, 79, 1132–1135. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, X.; Guo, R.; Akami, M.; Niu, C. Nitrogen Acquisition Strategies Mediated by Insect Symbionts: A Review of Their Mechanisms, Methodologies, and Case Studies. Insects 2022, 13, 84. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010084
Ren X, Guo R, Akami M, Niu C. Nitrogen Acquisition Strategies Mediated by Insect Symbionts: A Review of Their Mechanisms, Methodologies, and Case Studies. Insects. 2022; 13(1):84. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010084
Chicago/Turabian StyleRen, Xueming, Ruxin Guo, Mazarin Akami, and Changying Niu. 2022. "Nitrogen Acquisition Strategies Mediated by Insect Symbionts: A Review of Their Mechanisms, Methodologies, and Case Studies" Insects 13, no. 1: 84. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010084