
Herbivore-Induced Rice Volatiles Attract and Affect the Predation Ability of the Wolf Spiders, Pirata subpiraticus and Pardosa pseudoannulata

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Spiders
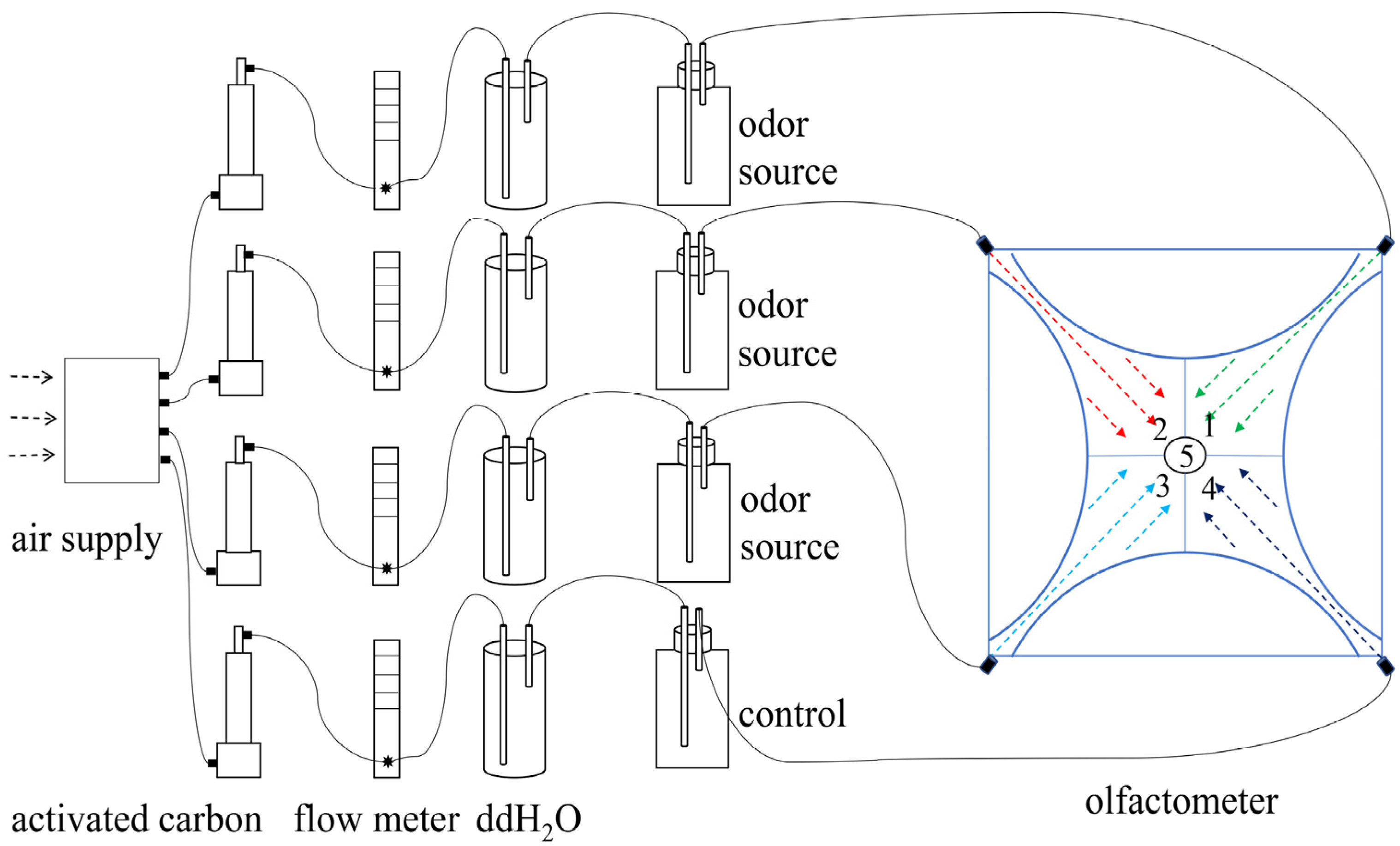
2.2. Volatiles
2.3. Experimental Treatments
2.3.1. Spider Selection Behavior Experiments in Response to HIRVs
2.3.2. Spider’s Predatory Ability Experiments
2.3.3. Field Trapping Experiments
2.4. Statistical Analysis
3. Results
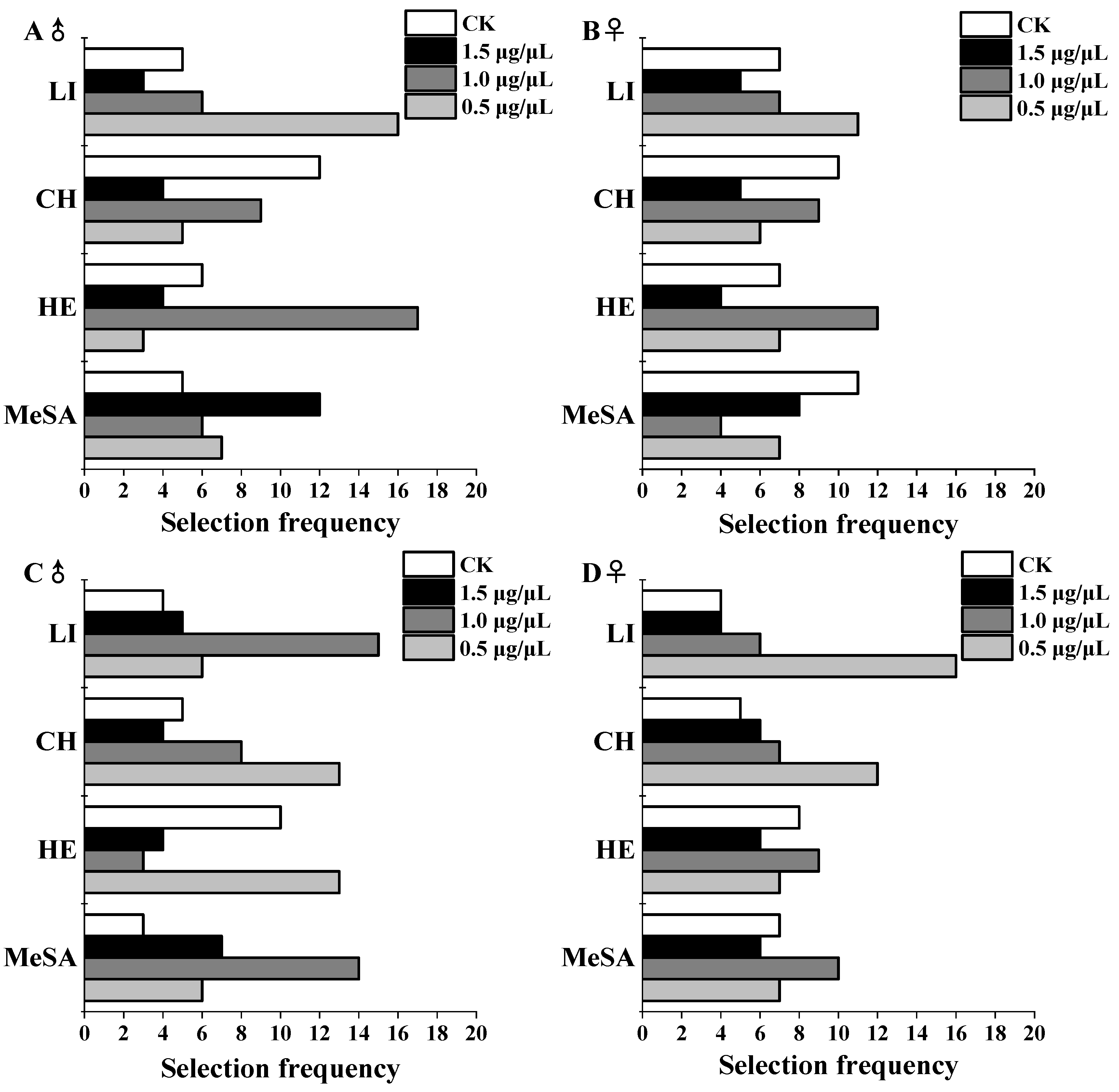
3.1. Spider Selection Behavior in Response to HIRVs
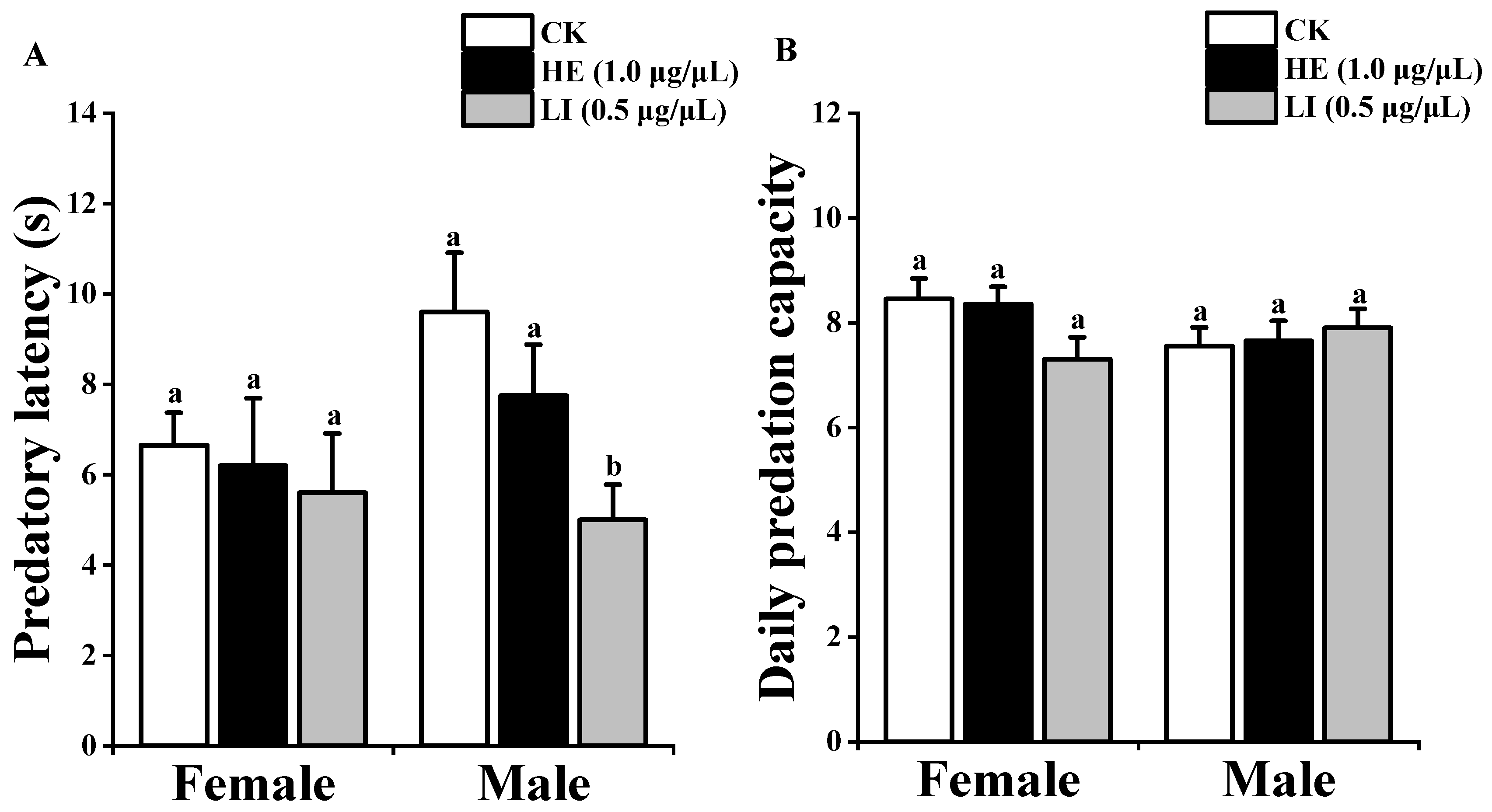
3.2. Spider’s Predatory Ability
3.2.1. Daily Predation Capacity and Predatory Latency of P. subpiraticus
3.2.2. Daily Predation Capacity and Predatory Latency of P. pseudoannulata
3.3. Field Trapping
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, L.L.; Wang, E.L.; Zhu, Y.; Tang, L. Contrasting effects of warming and autonomous breeding on single-rice productivity in China. Agric. Ecosyst. Environ. 2012, 149, 20–29. [Google Scholar] [CrossRef]
- Lou, Y.-G.; Zhang, G.-R.; Zhang, W.-Q.; Hu, Y.; Zhang, J. Biological control of rice insect pests in China. Biol. Control. 2013, 67, 8–20. [Google Scholar] [CrossRef]
- Qin, S.-l.; Lu, X.-y. Do large-scale farmers use more pesticides? Empirical evidence from rice farmers in five Chinese provinces. J. Integr. Agric. 2020, 19, 590–599. [Google Scholar] [CrossRef]
- Tu, L.H.; Boulange, J.; Phong, T.K.; Thuyet, D.Q.; Watanabe, H.; Takagi, K. Predicting rice pesticide fate and transport following foliage application by an updated PCPF-1 model. J. Environ. Manag. 2021, 277, 111356. [Google Scholar] [CrossRef] [PubMed]
- Tuanol, A.P.P.; Xu, Z.; Castillo, M.B.; Mamaril, C.P.; Manaois, R.V.; Romero, M.V.; Juliano, B.O. Content of Tocols, gamma-Oryzanol and Total Phenolics and Grain Quality of Brown Rice and Milled Rice Applied with Pesticides and Organic and Inorganic Nitrogen Fertilizer. Philipp. Agric. Sci. 2011, 94, 211–216. [Google Scholar]
- Dicke, M. Behavioural and community ecology of plants that cry for help. Plant Cell Environ. 2009, 32, 654–665. [Google Scholar] [CrossRef]
- War, A.R.; Sharma, H.C.; Paulraj, M.G.; War, M.Y.; Ignacimuthu, S. Herbivore induced plant volatiles: Their role in plant defense for pest management. Plant Signal. Behav. 2011, 6, 1973–1978. [Google Scholar] [CrossRef] [Green Version]
- Dicke, M.; Baldwin, I.T. The evolutionary context for herbivore-induced plant volatiles: Beyond the ‘cry for help’. Trends Plant Sci. 2010, 15, 167–175. [Google Scholar] [CrossRef]
- Mao, G.; Tian, J.; Li, T.; Fouad, H.; Mo, J. Behavioral responses of Anagrus nilaparvatae to common terpenoids, aromatic compounds, and fatty acid derivatives from rice plants. Entomol. Exp. Appl. 2018, 166, 483–490. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; ul Malook, S.; Shen, G.; Gao, L.; Zhang, C.; Li, J.; Zhang, J.; Wang, L.; Wu, J. Current understanding of maize and rice defense against insect herbivores. Plant Divers. 2018, 40, 189–195. [Google Scholar] [CrossRef]
- Zhao, N.; Zhuang, X.; Shrivastava, G.; Chen, F. Analysis of insect-induced volatiles from rice. Methods Mol. Biol. 2013, 956, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Zhou, Q.; Xia, Q.; Zhang, W.-Q.; Zhang, G.-R.; Gu, D.-X. Effect of insect pest-induced rice volatiles on the host selection behavior of brown planthopper. Chin. Sci. Bull. 2002, 849–853. [Google Scholar]
- Yan, F.; Wang, X.; Lu, J.; Pang, B.-P.; Lou, Y.-G. Comparison of rice volatiles induced by the larvae of Chilo suppressalis and Cnaphalocrocis medinalis. Chin. J. Appl. Entomol. 2010, 47, 96–101. [Google Scholar]
- Liu, X.-L.; Lou, Y.-G. Comparison of the defense responses in rice induced by brown planthopper Nilaparvata lugens (Stål) and white-backed planthopper Sogatella furcifera (Horváth). J. Plant Prot. 2018, 45, 971–978. [Google Scholar]
- Huang, T.-F.; Ma, Y.; Tang, B.-J.; Wang, B.-Y.; Zeng, R.-X.; Zhou, Q.; Zhang, G.-R. Sexual differences in the response of the parasitoid wasp Anagrus nilaparvatae to rice volatiles. Chin. J. Appl. Entomol. 2021, 58, 876–884. [Google Scholar]
- Li, T.; Wang, C.-P.; Jiang, N.-N.; Wei, J.-G.; Mo, J.-C. Attractiveness of rice plant volatiles to Anagrus nilaparvatae Pang et Wang. Chin. J. Appl. Entomol. 2018, 55, 360–367. [Google Scholar]
- Li, S.; Chen, W.-L.; Jin, D.-C.; Yang, H. The attraction of diverse rice volatiles to Haplogonatopus japonicus. J. Plant Prot. 2014, 41, 203–209. [Google Scholar]
- Jiang, N.-N.; Mao, G.-F.; Li, T.; Mo, J.-C. The olfactory behavior response of Cytorhinus lividipennis to single component of rice volatiles. Ecol. Environ. Sci. 2018, 27, 262–267. [Google Scholar]
- Lou, Y.-G.; Chen, J.-A. The role of rice volatiles in the predation behavior of A. lucorum. Entomol. Sin. 2001, 8, 240–250. [Google Scholar]
- Zhang, Y.-H.; Li, T.; Mo, J.-C. The attractiveness of rice plant volatiles to Apanteles chilonis Munakata and Anagrus nilaparvatae Pang et Wang. Chin. J. Appl. Entomol. 2016, 53, 491–498. [Google Scholar]
- Ulhoa, L.A.; Barrigossi, J.A.F.; Borges, M.; Laumann, R.A.; Blassioli-Moraes, M.C. Differential induction of volatiles in rice plants by two stink bug species influence behaviour of conspecifics and their natural enemy Telenomus podisi. Entomol. Exp. Appl. 2020, 168, 76–90. [Google Scholar] [CrossRef]
- Sun, L.-Y.; Liu, J.; Li, Q.; Fu, D.; Zhu, J.-Y.; Guo, J.-J.; Xiao, R.; Jin, D.-C. Cloning and differential expression of three heat shock protein genes associated with thermal stress from the wolf spider Pardosa pseudoannulata (Araneae: Lycosidae). J. Asia-Pac. Entomol. 2021, 24, 158–166. [Google Scholar] [CrossRef]
- Wang, X.-Q.; Wang, G.-H.; Zhu, Z.-R.; Tang, Q.-Y.; Hu, Y.; Qiao, F.; Heong, K.L.; Cheng, J.-A. Spider (Araneae) predations on white-backed planthopper Sogatella furcifera in subtropical rice ecosystems, China. Pest Manag. Sci. 2017, 73, 1277–1286. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-Q.; Shi, G.-B. Discussion on Dominant Species and Causes of Spiders in Rice Fields in China. Sci. Agric. Sin. 2002, 11, 85–93. [Google Scholar]
- Wang, H.-Q.; Yan, H.-M. Study on the Ecology and Utilization of Spiders in Rice Fields in China. Sci. Agric. Sin. 1996, 29, 68–75. [Google Scholar]
- Wang, Z.; Zeng, B.-P.; Li, W.-J.; Wang, W.-B. Study on the Time Niche of the Paddy field Spiders and Objective Pests. Hunan Agric. Sci. 2002, 3, 28–29. [Google Scholar]
- Yan, H.-M. The Structure and Ecological Distribution of Spider Community in Rice Fields in Southwest China. J. Nat. Sci. Hunan Norm. Univ. 1991, 14, 80–85. [Google Scholar]
- Tan, Z.-J.; Yan, H.-M. Chemical Information between Spiders’Species and Within Species. Life Sci. Res. 2015, 19, 368–371. [Google Scholar]
- Wang, B.; Huang, T.; Han, M.; Wang, X.-L.; Chang, Y.-T.; Huang, X.-Y.; Li, Y.-Y.; Yan, H.-M. The Response of Pardosa pseudoannulata’s Chemoreceptor in Locating the Prey. Sichuan J. Zool. 2014, 33, 86–89. [Google Scholar]
- Cao, Y.; Liu, J.; Guo, J.; Wu, H.; Zhang, G. Identification and analysis of odorant-binding protein genes from the wolf spider Pardosa pseudoannulata (Araneae: Lycosidae) based on its transcriptome. Chemoecology 2018, 28, 123–130. [Google Scholar] [CrossRef]
- Vizueta, J.; Frias-Lopez, C.; Macias-Hernandez, N.; Arnedo, M.A.; Sanchez-Gracia, A.; Rozas, J. Evolution of Chemosensory Gene Families in Arthropods: Insight from the First Inclusive Comparative Transcriptome Analysis across Spider Appendages. Genome Biol. Evol. 2017, 9, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Yin, C.-M.; Peng, X.-J.; Yan, H.-M. Zoology of Hunan, Spiders; Hunan Science and Technology Press: Hunan, China, 2012. [Google Scholar]
- Wen, D.-D.; He, Y.-Y.; Lu, Z.-Y.; Yang, H.-M.; Wang, H.-Q. quantitative study of biomass flow in the rice-Sogatella furcifera-Pirata subpiraticus food chain using fluorescent substance tracing. Acta Entomol. Sin. 2003, 46, 178–183. [Google Scholar]
- Xiao, Y.-H.; He, Y.-Y.; Yang, H.-M.; Yan, H.-M. The Study on the Starvation Endurance of Larva Pirata subpiraticus. J. Nat. Sci. Hunan Norm. Univ. 2004, 27, 78–81. [Google Scholar]
- Wang, Z. Bionomics and behavior of the wolf spider Pardosa pseudoannulata (Araneae: Lycosidae). Acta Entomol. Sin. 2007, 9, 927–932. [Google Scholar]
- Vet, L.E.M.; Lenteren, J.C.V.; Heymans, M.V.; Meelis, E. An airflow olfactometer for measuring olfactory responses of hymenopterous parasitoids and other small insects. Physiol. Entomol. 1983, 8, 97–106. [Google Scholar] [CrossRef]
- Verheggen, F.J.; Fagel, Q.; Heuskin, S.; Lognay, G.; Francis, F.; Haubruge, E. Electrophysiological and behavioral responses of the multicolored Asian lady beetle, Harmonia axyridis pallas, to sesquiterpene semiochemicals. J. Chem. Ecol. 2007, 33, 2148–2155. [Google Scholar] [CrossRef] [PubMed]
- Skoczek, A.; Piesik, D.; Wenda-Piesik, A.; Buszewski, B.; Bocianowski, J.; Wawrzyniak, M. Volatile organic compounds released by maize following herbivory or insect extract application and communication between plants. J. Appl. Entomol. 2017, 141, 630–643. [Google Scholar] [CrossRef]
- Piesik, D.; Rochat, D.; Delaney, K.J.; Marion-Poll, F. Orientation of European corn borer first instar larvae to synthetic green leaf volatiles. J. Appl. Entomol. 2013, 137, 234–240. [Google Scholar] [CrossRef] [Green Version]
- Avila, G.A.; Withers, T.M.; Holwell, G.I. Olfactory cues used in host-habitat location and host location by the parasitoid Cotesia urabae. Entomol. Exp. Appl. 2016, 158, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Ghaninia, M.; Amooghli Tabari, M. Olfactory cues explain differential attraction of the striped rice stem borer to different varieties of rice plant. J. Appl. Entomol. 2016, 140, 376–385. [Google Scholar] [CrossRef]
- Adedipe, F.; Park, Y.-L. Visual and olfactory preference of Harmonia axyridis (Coleoptera: Coccinellidae) adults to various companion plants. J. Asia-Pac. Entomol. 2010, 13, 319–323. [Google Scholar] [CrossRef]
- Siddique, B.; Tariq, M.; Naeem, M.; Ali, M. Behavioral Responses of Coccinella septempunctata and Diaeretiella rapae under the Influence of Semiochemicals and Plant Extract in Four Arm Olfactometer. Pak. J. Zool. 2019, 51, 1403–1411. [Google Scholar] [CrossRef]
- Komatsuzaki, S.; Piyasaengthong, N.; Matsuyama, S.; Kainoh, Y. Effect of Leaf Maturity on Host Habitat Location by the Egg-Larval Parasitoid Ascogaster reticulata. J. Chem. Ecol. 2021, 47, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Nurkomar, I.; Buchori, D.; Taylor, D.; Kainoh, Y. Innate olfactory responses of female and male parasitoid Apanteles taragamae Viereck (Hymenoptera: Braconidae) toward host plant infested by the cucumber moth Diaphania indica Saunders (Lepidoptera: Crambidae). Biocontrol Sci. Technol. 2017, 27, 1373–1382. [Google Scholar] [CrossRef]
- Zhu, J.; Park, K.-C. Methyl salicylate, a soybean aphid-induced plant volatile attractive to the predator Coccinella septempunctata. J. Chem. Ecol. 2005, 31, 1733–1746. [Google Scholar] [CrossRef]
- De Boer, J.G.; Dicke, M. The role of methyl salicylate in prey searching behavior of the predatory mite Phytoseiulus persimilis. J. Chem. Ecol. 2004, 30, 255–271. [Google Scholar] [CrossRef]
- James, D.G. Methyl salicylate is a field attractant for the goldeneyed lacewing, Chrysopa oculata. Biocontrol Sci. Technol. 2007, 16, 107–110. [Google Scholar] [CrossRef]
- James, D.G. Field evaluation of herbivore-induced plant volatiles as attractants for beneficial insects: Methyl salicylate and the green lacewing, Chrysopa nigricornis. J. Chem. Ecol. 2003, 29, 1601–1609. [Google Scholar] [CrossRef]
- James, D.G.; Price, T.S. Field-Testing of Methyl Salicylate for Recruitment and Retention of Beneficial Insects in Grapes and Hops. J. Chem. Ecol. 2004, 30, 1613–1628. [Google Scholar] [CrossRef] [PubMed]
- Shimoda, T. A key volatile infochemical that elicits a strong olfactory response of the predatory mite Neoseiulus californicus, an important natural enemy of the two-spotted spider mite Tetranychus urticae. Exp. Appl. Acarol. 2010, 50, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Carroll, M.J.; Schmelz, E.A.; Meagher, R.L.; Teal, P.E. Attraction of Spodoptera frugiperda larvae to volatiles from herbivore-damaged maize seedlings. J. Chem. Ecol. 2006, 32, 1911–1924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, A.R.; Wanner, K.W.; Trowell, S.C.; Warr, C.G.; Jaquin-Joly, E.; Zagatti, P.; Robertson, H.; Newcomb, R.D. Molecular basis of female-specific odorant responses in Bombyx mori. Insect Biochem. Mol. Biol. 2009, 39, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Reisenman, C.E.; Riffell, J.A.; Bernays, E.A.; Hildebrand, J.G. Antagonistic effects of floral scent in an insect-plant interaction. Proc. R. Soc. B-Biol. Sci. 2010, 277, 2371–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruz-López, L.; Díaz-Díaz, B.; Rojas, J.C. Coffee volatiles induced after mechanical injury and beetle herbivory attract the coffee berry borer and two of its parasitoids. Arthropod-Plant Interact. 2016, 10, 151–159. [Google Scholar] [CrossRef]
- Mitra, S.; Karmakar, A.; Mukherjee, A.; Barik, A. The Role of Leaf Volatiles of Ludwigia octovalvis (Jacq.) Raven in the Attraction of Altica cyanea (Weber) (Coleoptera: Chrysomelidae). J. Chem. Ecol. 2017, 43, 679–692. [Google Scholar] [CrossRef]
- Oelbermann, K.; Scheu, S. Control of aphids on wheat by generalist predators: Effects of predator density and the presence of alternative prey. Entomol. Exp. Appl. 2009, 132, 225–231. [Google Scholar] [CrossRef]
- Zhang, N.; Xie, L.; Wu, X.; Liu, K.; Liu, C.; Yan, Y. Development, survival and reproduction of a potential biological control agent, Lasioseius japonicus Ehara (Acari: Blattisociidae), on eggs of Drosophila melanogaster (Diptera: Drosophilidae) and Sitotroga cerealella (Lepidoptera: Gelechiidae). Syst. Appl. Acarol. 2020, 25, 1461–1471. [Google Scholar] [CrossRef]
- Cheli, G.; Armendano, A.; Gonzalez, A. Feeding preferences of the spider Misumenops pallidus (Araneae: Thomisidae) on potential prey insects from alfalfa crops. Rev. Biol. Trop. 2006, 54, 505–513. [Google Scholar] [CrossRef] [Green Version]
- Hauge, M.S.; Nielsen, F.H.; Toft, S. Weak responses to dietary enrichment in a specialized aphid predator. Physiol. Entomol. 2011, 36, 360–367. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, X.-L.; Liu, B.-L.; Qin, X.-Y.; Guo, H.; Feng, R. Rice volatile components induced by rice planthopper. J. South. Agric. 2021, 52, 37–44. [Google Scholar]
- Sun, Z.-Y. The Effect of Two Plant Extracts on the Insect Control Potential of Pardosa pseudoannulata; Hunan Agricultural University: Hunan, China, 2010. [Google Scholar]
- Sun, Z.-Y.; Liang, P.-Z.; Hu, R.; Wang, Z. Effect of Plant Physiologically Active Substances on the Predation of Pardosa pseudoannulata. Hunan Agric. Sci. 2009, 39, 85–87. [Google Scholar] [CrossRef]
- Li, R.; Li, N.; Liu, J.; Li, S.-C.; Hong, J.-P. The effect of low-dose of pesticide on predation of spider and its preliminary mechanisms. Acta Ecol. Sin. 2014, 34, 2629–2637. [Google Scholar]
- Li, R.; Li, S.-L.; Li, N.; Li, S.-C.; Hong, J.-P. The effects of low doses of insecticides on predation of green peach aphid Myzus persicae (Hemiptera: Aphididae) by a wolf spider Pardosa astrigera (Araneae: Lycosidae). J. Plant Prot. 2014, 41, 711–716. [Google Scholar]
- Wang, Z.; Song, D.-X.; Zhu, M.-S. Influences of Low-dose Pesticide on the Spatial Niche and Insect-controlling Efficiency of Paddy Field Spiders. J. Hebei Univ. (Nat. Sci. Ed.) 2006, 26, 278–282. [Google Scholar]
- Wang, Z.; Yan, H.-M.; Wang, H.-Q. Effects of low-dose pesticides on insect control ability of rice field spiders. Acta Ecol. Sin. 2002, 22, 346–351. [Google Scholar]
- Simpson, M.; Gurr, G.M.; Simmons, A.T.; Wratten, S.D.; James, D.G.; Leeson, G.; Nicol, H.I.; Orre-Gordon, G.U.S. Attract and reward: Combining chemical ecology and habitat manipulation to enhance biological control in field crops. J. Appl. Ecol. 2011, 48, 580–590. [Google Scholar] [CrossRef]
- Jaworski, C.C.; Xiao, D.; Xu, Q.; Ramirez-Romero, R.; Guo, X.; Wang, S.; Desneux, N. Varying the spatial arrangement of synthetic herbivore-induced plant volatiles and companion plants to improve conservation biological control. J. Appl. Ecol. 2019, 56, 1176–1188. [Google Scholar] [CrossRef]
- Liu, J.; Sun, L.-y.; Fu, D.; Zhu, J.-Y.; Xiao, R. Diversity, and dominant species, of rice field spiders in Guizhou province. Chin. J. Appl. Entomol. 2021, 58, 142–157. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spider | Sex | Treatment | χ2 | p | Significance |
|---|---|---|---|---|---|
| P. pseudoannulata | male | CK | 2.533 | 0.469 | NS |
| MeSA | 8.667 | 0.034 | * | ||
| HE | 9.200 | 0.027 | * | ||
| CH | 6.533 | 0.088 | NS | ||
| LI | 10.267 | 0.016 | * | ||
| female | CK | 2.267 | 0.519 | NS | |
| MeSA | 1.200 | 0.753 | NS | ||
| HE | 0.667 | 0.881 | NS | ||
| CH | 3.867 | 0.276 | NS | ||
| LI | 13.200 | 0.004 | ** | ||
| P. subpiraticus | male | CK | 5.467 | 0.141 | NS |
| MeSA | 3.867 | 0.276 | NS | ||
| HE | 16.667 | 0.001 | ** | ||
| CH | 5.467 | 0.141 | NS | ||
| LI | 13.467 | 0.004 | ** | ||
| female | CK | 7.600 | 0.055 | NS | |
| MeSA | 3.333 | 0.343 | NS | ||
| HE | 4.400 | 0.221 | NS | ||
| CH | 2.267 | 0.519 | NS | ||
| LI | 2.533 | 0.469 | NS |
| Spider Name | Sex | Treatment | 0.5 μg/μL | 1.0 μg/μL | 1.5 μg/μL | ck |
|---|---|---|---|---|---|---|
| P. pseudoannulata | male | CH | 2.151 ± 0.457 a | 1.323 ± 0.407 ab | 0.661 ± 0.313 b | 0.826 ± 0.343 ab |
| MeSA | 0.990 ± 0.368 b | 2.313 ± 0.459 a | 1.157 ± 0.381 b | 0.494 ± 0.275 b | ||
| HE | 2.110 ± 0.450 a | 0.534 ± 0.273 b | 0.668 ± 0.312 b | 1.652 ± 0.434 a b | ||
| LI | 1.171 ± 0.387 b | 2.295 ± 0.456 a | 0.822 ± 0.335 b | 0.661 ± 0.313 b | ||
| female | CH | 1.986 ± 0.452 a | 1.161 ± 0.391 a | 0.993 ± 0.369 a | 0.827 ± 0.343 a | |
| MeSA | 1.157 ± 0.390 a | 1.651 ± 0.434 a | 0.992 ± 0.368 a | 1.156 ± 0.389 a | ||
| HE | 1.161 ± 0.391 a | 1.491 ± 0.423 a | 0.990 ± 0.368 a | 1.326 ± 0.409 a | ||
| LI | 2.640 ± 0.456 a | 0.989 ± 0.367 b | 0.656 ± 0.311 b | 0.655 ± 0.310 b | ||
| P. subpiraticus | male | CH | 0.827 ± 0.343 a | 1.489 ± 0.424 a | 0.661 ± 0.313 a | 1.987 ± 0.452 a |
| MeSA | 1.156 ± 0.389 a | 1.154 ± 0.390 a | 1.821 ± 0.445 a | 0.828 ± 0.344 a | ||
| HE | 0.825 ± 0.342 b | 2.474 ± 0.460 a | 0.661 ± 0.313 b | 0.991 ± 0.368 b | ||
| LI | 2.647 ± 0.460 a | 0.991 ± 0.368 b | 0.496 ± 0.276 b | 0.826 ± 0.343 b | ||
| female | CH | 0.993 ± 0.368 a | 1.486 ± 0.421 a | 0.836 ± 0.343 a | 1.656 ± 0.434 a | |
| MeSA | 1.159 ± 0.390 a | 0.662 ± 0.313 a | 1.324 ± 0.407 a | 1.819 ± 0.443 a | ||
| HE | 1.167 ± 0.392 a | 1.986 ± 0.451 a | 0.662 ± 0.313 a | 1.157 ± 0.389 a | ||
| LI | 1.820 ± 0.444 a | 1.158 ± 0.389 a | 0.828 ± 0.344 a | 1.157 ± 0.386 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Sun, L.; Fu, D.; Zhu, J.; Liu, M.; Xiao, F.; Xiao, R. Herbivore-Induced Rice Volatiles Attract and Affect the Predation Ability of the Wolf Spiders, Pirata subpiraticus and Pardosa pseudoannulata. Insects 2022, 13, 90. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010090
Liu J, Sun L, Fu D, Zhu J, Liu M, Xiao F, Xiao R. Herbivore-Induced Rice Volatiles Attract and Affect the Predation Ability of the Wolf Spiders, Pirata subpiraticus and Pardosa pseudoannulata. Insects. 2022; 13(1):90. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010090
Chicago/Turabian StyleLiu, Jing, Liangyu Sun, Di Fu, Jiayun Zhu, Min Liu, Feng Xiao, and Rong Xiao. 2022. "Herbivore-Induced Rice Volatiles Attract and Affect the Predation Ability of the Wolf Spiders, Pirata subpiraticus and Pardosa pseudoannulata" Insects 13, no. 1: 90. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010090