
Species-Specificity in Thermopreference and CO2-Gated Heat-Seeking in Culex Mosquitoes



Abstract
:Simple Summary
Abstract
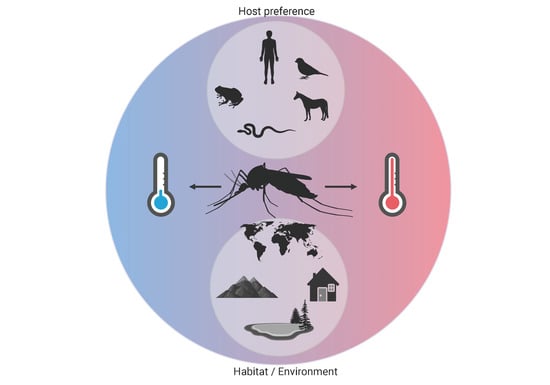
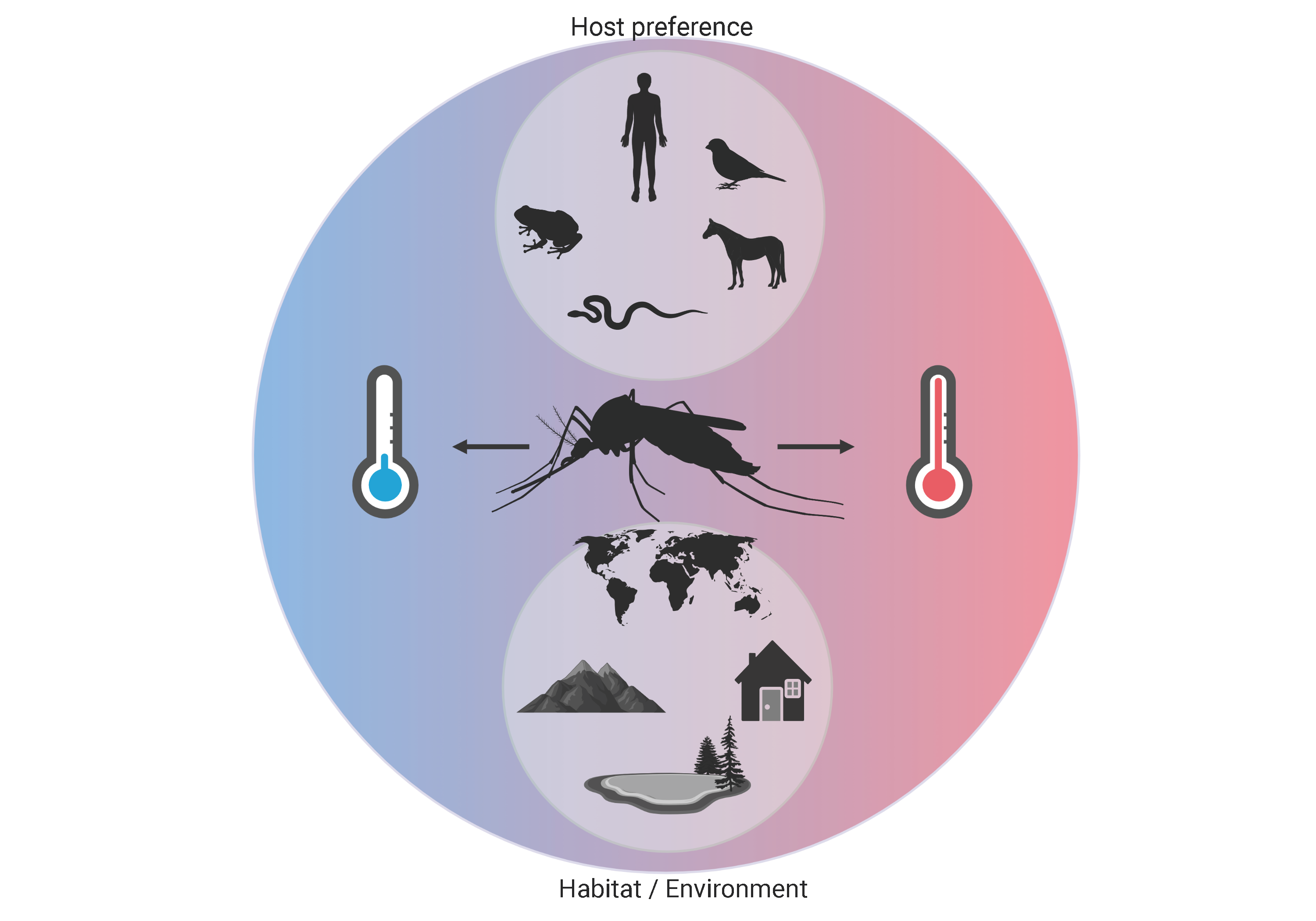
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
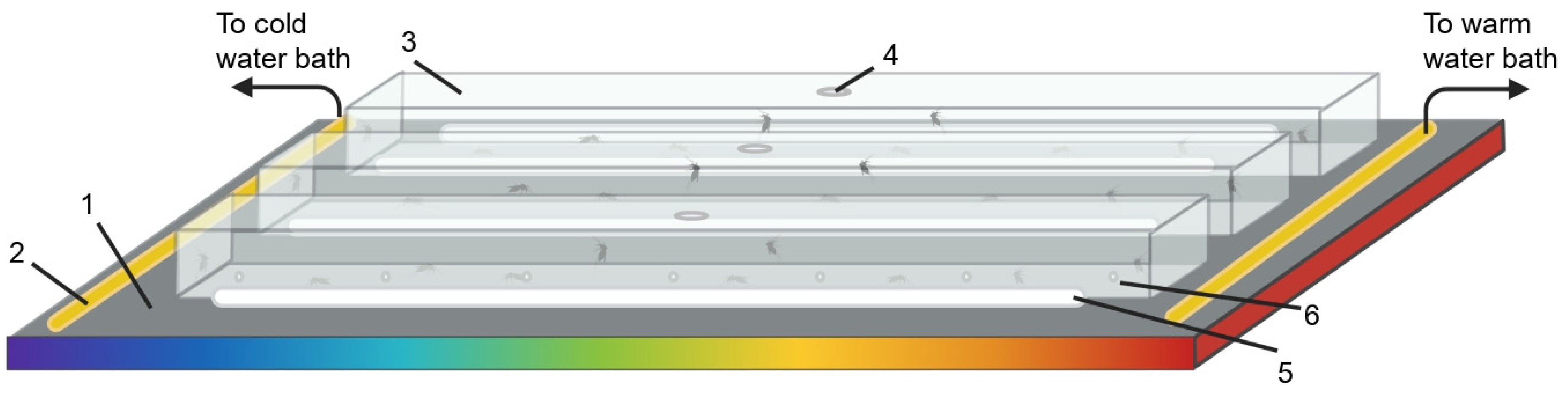
2.2. Thermopreference Assays
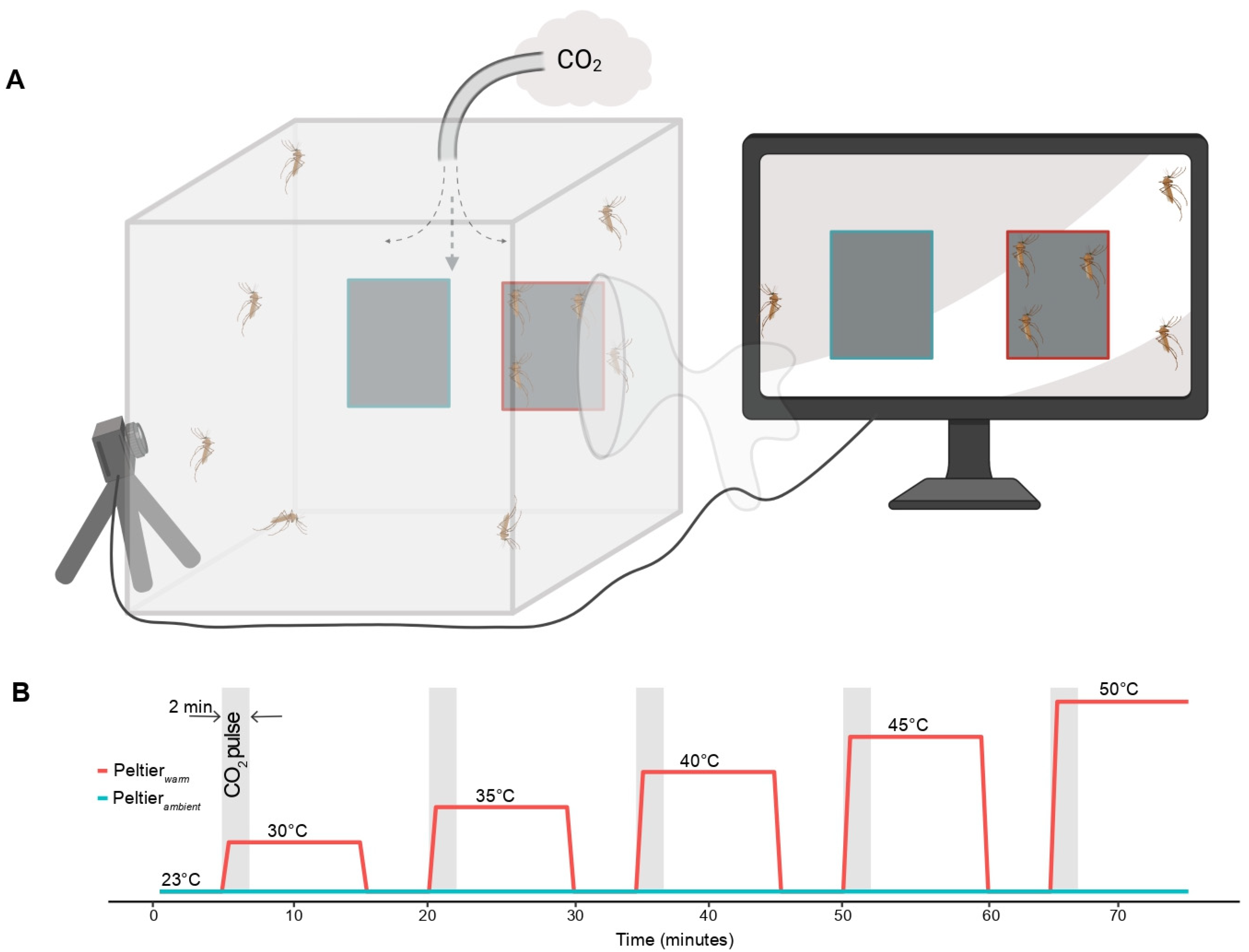
2.3. CO2-Gated Heat Seeking Assays
2.4. Statistical Analyses
2.4.1. Thermopreference Assays
2.4.2. CO2-Gated Heat Seeking Assays
3. Results
3.1. Thermopreference Assays
3.2. CO2-Gated Heat Seeking Assays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heinrich, B. Insect thermoregulation. Endeavour 1995, 19, 28–33. [Google Scholar] [CrossRef]
- Benoit, J.B.; Lazzari, C.R.; Denlinger, D.L.; Lahondère, C. Thermoprotective adaptations are critical for arthropods feeding on warm-blooded hosts. Curr. Opin. Insect Sci. 2019, 34, 7–11. [Google Scholar] [CrossRef]
- Dillon, M.E.; Wang, G.; Garrity, P.A.; Huey, R.B. Thermal preference in Drosophila. J. Therm. Biol. 2009, 34, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, A.L.; Miles, D.B. Natural selection on thermal preference, critical thermal maxima and locomotor performance. Proc. R. Soc. B Boil. Sci. 2017, 284, 20170536. [Google Scholar] [CrossRef] [Green Version]
- Lazzari, C.R. Temperature preference in Triatoma infestans (Hemiptera: Reduviidae). Bull. Entomol. Res. 1991, 81, 273–276. [Google Scholar] [CrossRef]
- Schilman, P.; Lazzari, C. Temperature preference in Rhodnius prolixus, effects and possible consequences. Acta Trop. 2004, 90, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Ciota, A.; Matacchiero, A.C.; Kilpatrick, A.M.; Kramer, L.D. The Effect of Temperature on Life History Traits of Culex Mosquitoes. J. Med. Entomol. 2014, 51, 55–62. [Google Scholar] [CrossRef]
- Eldridge, B.F. The Effect of Temperature and Photoperiod on Blood-Feeding and Ovarian Development in Mosquitoes of the Culex Pipiens Complex. Am. J. Trop. Med. Hyg. 1968, 17, 133–140. [Google Scholar] [CrossRef]
- Reinhold, J.M.; Lazzari, C.R.; Lahondère, C. Effects of the Environmental Temperature on Aedes aegypti and Aedes albopictus Mosquitoes: A Review. Insects 2018, 9, 158. [Google Scholar] [CrossRef] [Green Version]
- Rivas, G.; Teles-De-Freitas, R.; Pavan, M.G.; Lima, J.B.P.; Peixoto, A.A.; Bruno, R.V. Effects of Light and Temperature on Daily Activity and Clock Gene Expression in Two Mosquito Disease Vectors. J. Biol. Rhythm 2018, 33, 272–288. [Google Scholar] [CrossRef]
- Carnaghi, M.; Belmain, S.R.; Hopkins, R.J.; Hawkes, F.M. Multimodal synergisms in host stimuli drive landing response in malaria mosquitoes. Sci. Rep. 2021, 11, 7379. [Google Scholar] [CrossRef]
- Corfas, R.; Vosshall, L.B. The cation channel TRPA1 tunes mosquito thermotaxis to host temperatures. eLife 2015, 4, e11750. [Google Scholar] [CrossRef]
- Liu, M.Z.; Vosshall, L.B. General Visual and Contingent Thermal Cues Interact to Elicit Attraction in Female Aedes aegypti Mosquitoes. Curr. Biol. 2019, 29, 2250–2257. [Google Scholar] [CrossRef]
- Lazzari, C.R. The thermal sense of blood-sucking insects: Why physics matters. Curr. Opin. Insect Sci. 2019, 34, 112–116. [Google Scholar] [CrossRef]
- van Breugel, F.; Riffell, J.; Fairhall, A.; Dickinson, M.H. Mosquitoes Use Vision to Associate Odor Plumes with Thermal Targets. Curr. Biol. 2015, 25, 2123–2129. [Google Scholar] [CrossRef] [Green Version]
- Zermoglio, P.F.; Robuchon, E.; Leonardi, M.S.; Chandre, F.; Lazzari, C.R. What does heat tell a mosquito? Characterization of the orientation behaviour of Aedes aegypti towards heat sources. J. Insect Physiol. 2017, 100, 9–14. [Google Scholar] [CrossRef]
- Clarke, A.; Rothery, P. Scaling of body temperature in mammals and birds. Funct. Ecol. 2007, 22, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Prinzinger, R.; Preßmar, A.; Schleucher, E. Body temperature in birds. Comp. Biochem. Physiol. Part A Physiol. 1991, 99, 499–506. [Google Scholar] [CrossRef]
- Werner, A.; Gunga, H.-C. Monitoring of Core Body Temperature in Humans. Stress Chall. Immun. Space 2019, 477–498. [Google Scholar] [CrossRef]
- Verhulst, N.O.; Brendle, A.; Blanckenhorn, W.U.; Mathis, A. Thermal preferences of subtropical Aedes aegypti and temperate Ae. japonicus mosquitoes. J. Therm. Biol. 2020, 91, 102637. [Google Scholar] [CrossRef]
- Darcy, R.F.; Ward, R.A. Identification and Geographical Distribution of the Mosquitoes of North America, North of Mexico; University Press of Florida/State University System: Gainesville, FL, USA, 2005. [Google Scholar]
- Bartlett-Healy, K.; Crans, W.; Gaugler, R. Temporal and spatial synchrony of Culex territans (Diptera: Culicidae) with their amphibian hosts. J. Med. Entomol. 2008, 45, 1031–1038. [Google Scholar] [CrossRef]
- Godsey, M.S., Jr.; Burkhalter, K.; Delorey, M.; Savage, H.M. Seasonality and time of host-seeking activity of Culex tarsalis and floodwater Aedes in northern Colorado, 2006–2007. J. Am. Mosq. Control Assoc. 2010, 26, 148–159. [Google Scholar] [CrossRef]
- Campbell, G.L.; Marfin, A.A.; Lanciotti, R.S.; Gubler, D.J. West nile virus. Lancet Infect. Dis. 2002, 2, 519–529. [Google Scholar] [CrossRef]
- Gouck, H.K.; Godwin, D.R.; Schreck, C.E. Feeding patterns of six species of mosquitoes. Mosq. News 1971, 31, 99–101. [Google Scholar]
- Downe, A.E.R.; Archer, J.A. The Effects of Different Blood-Meal Sources on Digestion and Egg Production in Culex Tarsalis Coq. (Diptera: Culicidae). J. Med. Entomol. 1975, 12, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Kent, R.; Juliusson, L.; Weissmann, M.; Evans, S.; Komar, N.; Kading, R. Seasonal Blood-Feeding Behavior of Culex tarsalis (Diptera: Culicidae) in Weld County, Colorado, 2007. J. Med. Entomol. 2009, 46, 380–390. [Google Scholar] [CrossRef]
- Tempelis, C.H.; Reeves, W.C.; Bellamy, R.E.; Lofy, M.F. A Three-Year Study of the Feeding Habits of Culex Tarsalis in Kern County, California. Am. J. Trop. Med. Hyg. 1965, 14, 170–177. [Google Scholar] [CrossRef]
- Burkett-Cadena, N.; Eubanks, M.D.; Hassan, H.K.; Unnasch, T.R.; Guyer, C.; Graham, S.P.; Katholi, C.R. Blood Feeding Patterns of Potential Arbovirus Vectors of the Genus Culex Targeting Ectothermic Hosts. Am. J. Trop. Med. Hyg. 2008, 79, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Savage, H.M.; Aggarwal, D.; Apperson, C.S.; Katholi, C.R.; Gordon, E.; Hassan, H.K.; Anderson, M.; Charnetzky, D.; McMillen, L.; Unnasch, E.A.; et al. Host Choice and West Nile Virus Infection Rates in Blood-Fed Mosquitoes, Including Members of the Culex pipiens Complex, from Memphis and Shelby County, Tennessee, 2002–2003. Vector-Borne Zoonotic Dis. 2007, 7, 365–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malafronte, R.D.S.; Calvo, E.; A James, A.; Marinotti, O. The major salivary gland antigens of Culex quinquefasciatus are D7-related proteins. Insect Biochem. Mol. Biol. 2003, 33, 63–71. [Google Scholar] [CrossRef]
- Kent, R.J.; Crabtree, M.B.; Miller, B.R. Transmission of West Nile Virus by Culex quinquefasciatus Say Infected with Culex Flavivirus Izabal. PLoS Negl. Trop. Dis. 2010, 4, e671. [Google Scholar] [CrossRef] [PubMed]
- Molaei, G.; Andreadis, T.G.; Armstrong, P.M.; Bueno Jr, R.; Dennett, J.A.; Real, S.V.; Sargent, C.; Bala, A.; Randle, Y.; Guzman, H.; et al. Host feeding pattern of Culex quinquefasciatus (Diptera: Culicidae) and its role in transmission of West Nile virus in Harris County, Texas. Am. J. Trop. Med. 2007, 77, 73–81. [Google Scholar] [CrossRef] [Green Version]
- Nitatpattana, N.; Apiwathnasorn, C.; Barbazan, P.; Leemingsawat, S.; Yoksan, S.; Gonzalez, J.-P. First isolation of Japanese encephalitis from Culex quinquefasciatus in Thailand. Southeast Asian J. Trop. Med. Public Health 2005, 36, 875–878. [Google Scholar]
- Wang, Z.; Zhang, X.; Li, C.; Zhang, Y.; Xing, D.; Wu, Y.; Zhao, T. Vector Competence of Five Common Mosquito Species in the People’s Republic of China for Western Equine Encephalitis Virus. Vector-Borne Zoonotic Dis. 2012, 12, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Bartlett-Healy, K.; Crans, W.; Gaugler, R. Vertebrate Hosts and Phylogenetic Relationships of Amphibian Trypanosomes from a Potential Invertebrate Vector, Culex territans Walker (Diptera: Culicidae). J. Parasitol. 2009, 95, 381–387. [Google Scholar] [CrossRef]
- Desser, S.S.; Hong, H.; Martin, D.S. The Life History, Ultrastructure, and Experimental Transmission of Hepatozoon catesbianae n. comb., an Apicomplexan Parasite of the Bullfrog, Rana catesbeiana and the Mosquito, Culex territans in Algonquin Park, Ontario. J. Parasitol. 1995, 81, 212. [Google Scholar] [CrossRef]
- LaPointe, D.A.; Goff, M.L.; Atkinson, C.T. Comparative susceptibility of introduced forest-dwelling mosquitoes in Hawai’i to avian malaria, Plasmodium relictum. J. Parasitol. 2005, 91, 843–849. [Google Scholar] [CrossRef]
- Votýpka, J.; Szabova, J.; Radrova, J.; Zídková, L.; Svobodova, M. Trypanosoma culicavium sp. nov., an avian trypanosome transmitted by Culex mosquitoes. Int. J. Syst. Evol. 2012, 62, 745–754. [Google Scholar] [CrossRef] [Green Version]
- Ball, G.H.; Oda, S.N. Sexual Stages in the Life History of the Hemogregarine Hepatozoon rarefaciens (Sambon and Seligmann, 1907). J. Protozool. 1971, 18, 697–700. [Google Scholar] [CrossRef]
- Ritchie, M.W.; Dawson, J.W.; MacMillan, H.A. A simple and dynamic thermal gradient device for measuring thermal performance in small ectotherms. Curr. Res. Insect Sci. 2020, 1, 100005. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: http://www.r-project.org/ (accessed on 5 December 2021).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package 2018, 1, 3. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2016; ISBN 9783319242774. [Google Scholar]
- Sun, W.; Shults, J.; Leonard, M. A Note on the Use of Unbiased Estimating Equations to Estimate Correlation in Analysis of Longitudinal Trials. Biom. J. 2009, 51, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Samy, A.; Elaagip, A.H.; Kenawy, M.; Ayres, C.F.J.; Peterson, A.T.; Soliman, D. Climate Change Influences on the Global Potential Distribution of the Mosquito Culex quinquefasciatus, Vector of West Nile Virus and Lymphatic Filariasis. PLoS ONE 2016, 11, e0163863. [Google Scholar] [CrossRef] [PubMed]
- Reisen, W.K. The Contrasting Bionomics of Culex Mosquitoes in Western North America. J. Am. Mosq. Control Assoc. 2012, 28, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Benoit, J.B.; Denlinger, D.L. Suppression of water loss during adult diapause in the northern house mosquito, Culex pipiens. J. Exp. Biol. 2007, 210, 217–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benoit, J.B.; Lopez-Martinez, G.; Phillips, Z.P.; Patrick, K.R.; Denlinger, D.L. Heat shock proteins contribute to mosquito dehydration tolerance. J. Insect Physiol. 2010, 56, 151–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinehart, J.P.; Robich, R.M.; Denlinger, D.L. Enhanced cold and desiccation tolerance in diapausing adults of Culex pipiens, and a role for Hsp70 in response to cold shock but not as a component of the diapause program. J. Med. Entomol. 2006, 43, 713–722. [Google Scholar] [CrossRef] [Green Version]
- Anderson, A.W.; Harwood, R.F. Cold tolerance in adult female Culex tarsalis (Coquillett). Mosq. News 1966, 26. [Google Scholar]
- McMeniman, C.; Corfas, R.; Matthews, B.; Ritchie, S.A.; Vosshall, L.B. Multimodal Integration of Carbon Dioxide and Other Sensory Cues Drives Mosquito Attraction to Humans. Cell 2014, 156, 1060–1071. [Google Scholar] [CrossRef] [Green Version]
- Greppi, C.; Laursen, W.J.; Budelli, G.; Chang, E.C.; Daniels, A.M.; van Giesen, L.; Smidler, A.L.; Catteruccia, F.; Garrity, P.A. Mosquito heat seeking is driven by an ancestral cooling receptor. Science 2020, 367, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Reinhold, J.M.; Virginia Tech, Blacksburg, VA, USA; Roark, M.; University of Virginia at Wise, Wise, VA, USA; McLeod, D.; Unviersity of Virginia, Charlottesville, VA, USA; Lahondère, C.; Virginia Tech, Blacksburg, VA, USA. Carbon dioxide level detection in the green frog, Lithobates clamitans, and the bullfrog, Lithobates catesbeiannus. Unpublished work. 2021. [Google Scholar]
- Wheelwright, M.; Whittle, C.R.; Riabinina, O. Olfactory systems across mosquito species. Cell Tissue Res. 2021, 383, 75–90. [Google Scholar] [CrossRef]
- Wolff, G.H.; Riffell, J.A. Olfaction, experience and neural mechanisms underlying mosquito host preference. J. Exp. Biol. 2018, 221, 157131. [Google Scholar] [CrossRef] [Green Version]
- Miller, B.R.; Crabtree, M.B.; Savage, H.M. Phylogeny of fourteen Culex mosquito species, including the Culex pipiens complex, inferred from the internal transcribed spacers of ribosomal DNA. Insect Mol. Biol. 1996, 5, 93–107. [Google Scholar] [CrossRef]
- Harbach, R.E.; Culverwell, C.L.; Kitching, I.J. Phylogeny of the nominotypical subgenus of Culex (Diptera: Culicidae): Insights from analyses of anatomical data into interspecific relationships and species groups in an unresolved tree. Syst. Biodivers. 2017, 15, 296–306. [Google Scholar] [CrossRef]
- Weitzel, T.; Collado, A.; Jöst, A.; Pietsch, K.; Storch, V.; Becker, N. Genetic Differentiation of Populations within the Culex pipiens Complex and Phylogeny of Related Species. J. Am. Mosq. Control Assoc. 2009, 25, 6–17. [Google Scholar] [CrossRef]
- Reidenbach, K.R.; Cook, S.; Bertone, M.A.; Harbach, R.E.; Wiegmann, B.M.; Besansky, N.J. Phylogenetic analysis and temporal diversification of mosquitoes (Diptera: Culicidae) based on nuclear genes and morphology. BMC Evol. Biol. 2009, 9, 298. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Agramonte, N.; Linthicum, K.J.; Bloomquist, J.R. A Survey of Chemoreceptive Responses on Different Mosquito Appendages. J. Med. Entomol. 2020, 58, 475–479. [Google Scholar] [CrossRef]
- Stanczyk, N.M.; Brookfield, J.F.Y.; Ignell, R.; Logan, J.G.; Field, L.M. Behavioral insensitivity to DEET in Aedes aegypti is a genetically determined trait residing in changes in sensillum function. Proc. Natl. Acad. Sci. USA 2010, 107, 8575–8580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ditzen, M.; Pellegrino, M.; Vosshall, L.B. Insect Odorant Receptors Are Molecular Targets of the Insect Repellent DEET. Science 2008, 319, 1838–1842. [Google Scholar] [CrossRef] [Green Version]
- Owen, W.B. The contact chemoreceptor appendages of the mosquito and their function in feeding behaviour. J. Insect Physiol. 1963, 9, 73–87. [Google Scholar] [CrossRef]
- Kwon, H.-W.; Lu, T.; Rutzler, M.; Zwiebel, L.J. Olfactory responses in a gustatory organ of the malaria vector mosquito Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2006, 103, 13526–13531. [Google Scholar] [CrossRef] [Green Version]
- Dennis, E.J.; Goldman, O.V.; Vosshall, L.B. Aedes aegypti Mosquitoes Use Their Legs to Sense DEET on Contact. Curr. Biol. 2019, 29, 1551–1556. [Google Scholar] [CrossRef] [PubMed]
- McIver, S.B. Comparative study of antennal sense organs of female culicine mosquitoes. Can. Entomol. 1970, 102, 1258–1267. [Google Scholar] [CrossRef]
- Davis, E.E.; Sokolove, P.G. Temperature responses of antennal receptors of the mosquito, Aedes aegypti. J. Comp. Physiol. A 1975, 96, 223–236. [Google Scholar] [CrossRef]
- Wang, G.; Qiu, Y.T.; Lu, T.; Kwon, H.-W.; Pitts, R.J.; Van Loon, J.J.A.; Takken, W.; Zwiebel, L.J. Anopheles gambiae TRPA1 is a heat-activated channel expressed in thermosensitive sensilla of female antennae. Eur. J. Neurosci. 2009, 30, 967–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIver, S.B. Fine structure of antennal sensilla coeloconica of culicine mosquitoes. Tissue Cell 1973, 5, 105–112. [Google Scholar] [CrossRef]
- McIver, S.; Charlton, C. Studies on the sense organs on the palps of selected culicine mosquitoes. Can. J. Zool. 1970, 48, 293–295. [Google Scholar] [CrossRef]
- Healy, K.; Crans, W.; Gaugler, R. Phonotaxis to Amphibian Vocalizations in Culex territans (Diptera: Culicidae). Ann. Entomol. Soc. Am. 2008, 101, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Erdelyan, C.N.G.; Mahood, T.H.; Bader, T.S.Y.; Whyard, S. Functional validation of the carbon dioxide receptor genes in Aedes aegypti mosquitoes using RNA interference. Insect Mol. Biol. 2012, 21, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wen, X.; Leal, W.S. CO2 per se activates carbon dioxide receptors. Insect Biochem. Mol. Biol. 2020, 117, 103284. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ye, Z.; Baker, A.; Sun, H.; Zwiebel, L.J. Gene editing reveals obligate and modulatory components of the CO2 receptor complex in the malaria vector mosquito, Anopheles coluzzii. Insect Biochem. Mol. Biol. 2020, 127, 103470. [Google Scholar] [CrossRef] [PubMed]
- Robertson, H.M.; Kent, L.B. Evolution of the Gene Lineage Encoding the Carbon Dioxide Receptor in Insects. J. Insect Sci. 2009, 9, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
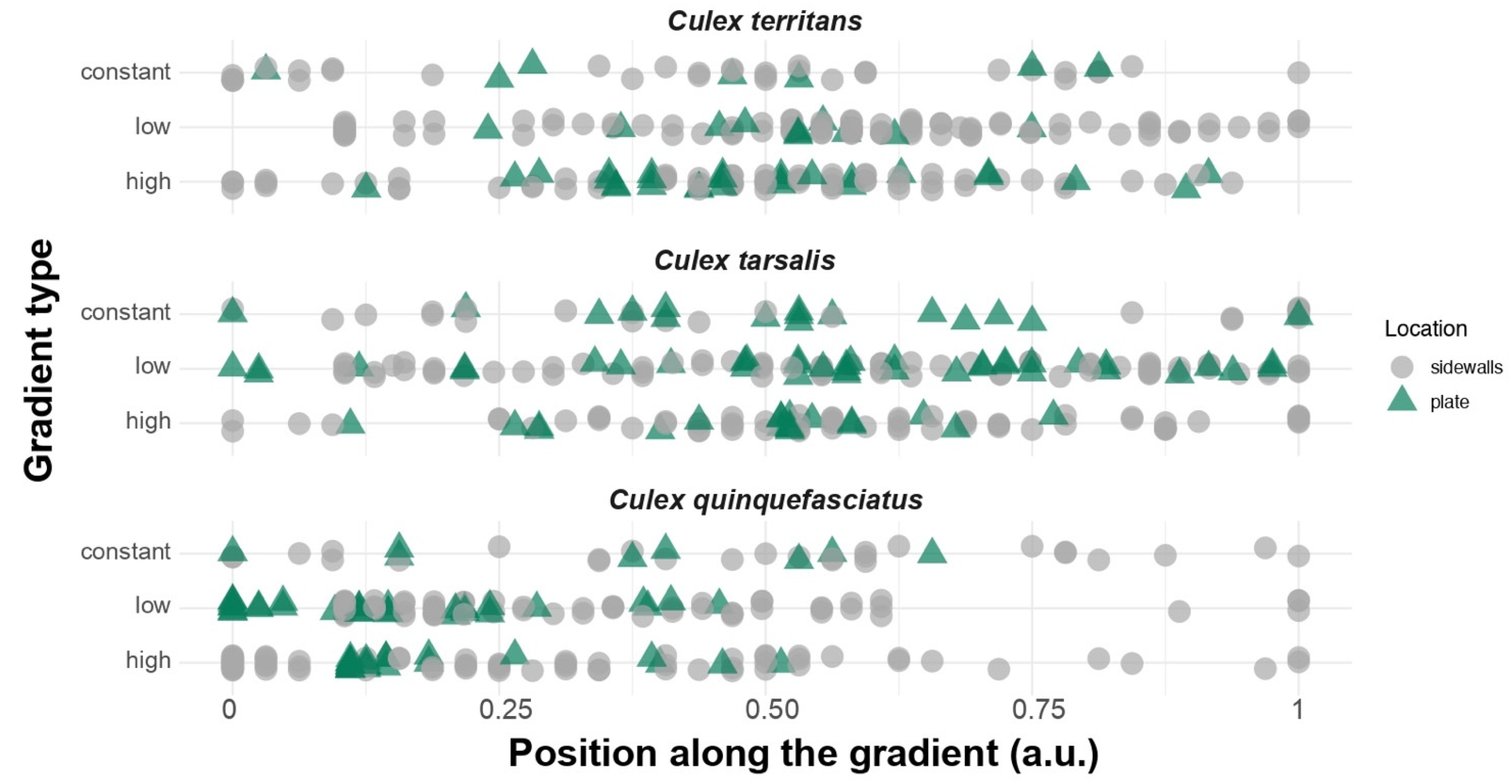{kind=link}
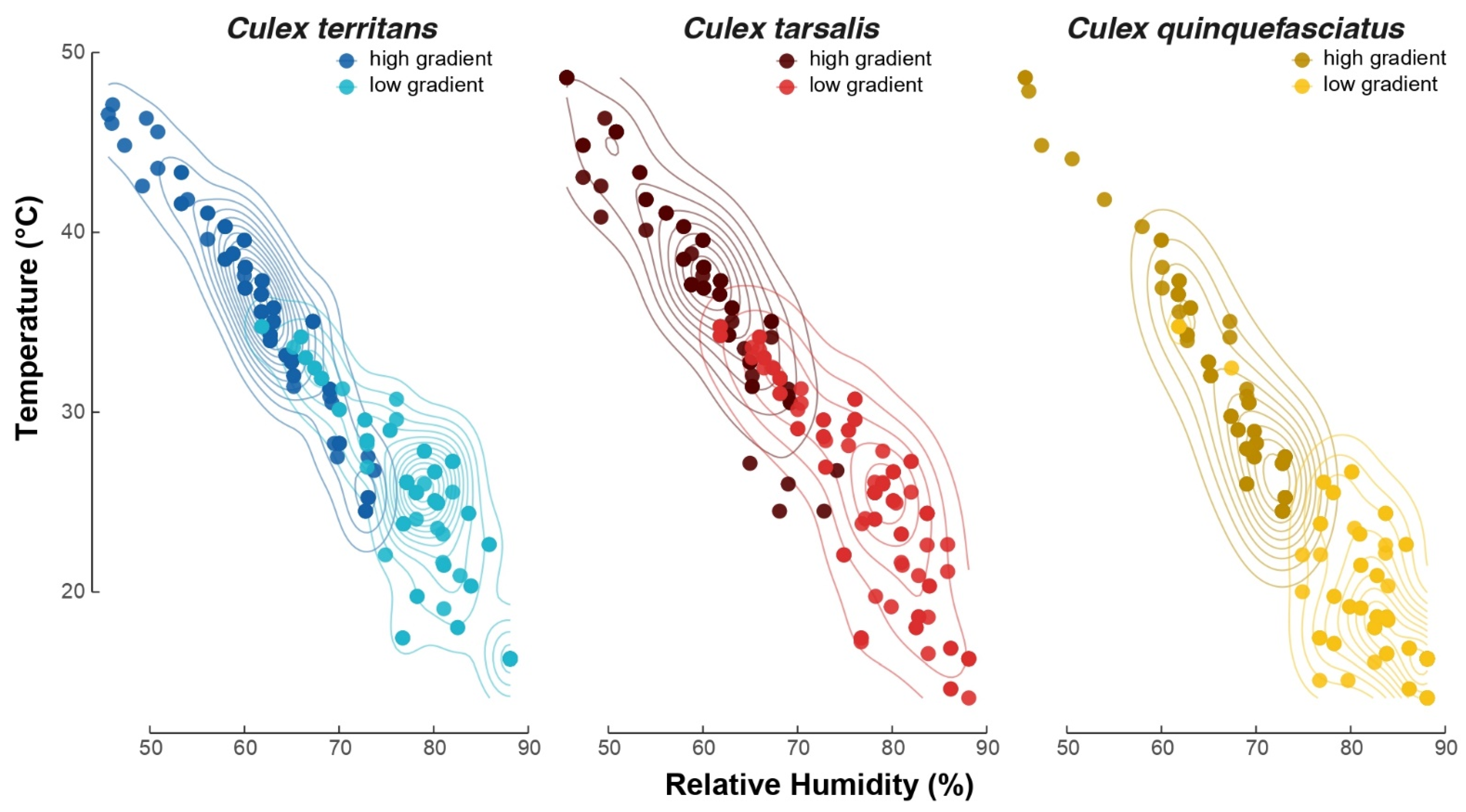{kind=link}
{kind=link}
| Species | Temperature of Peltierwarm | CO2 Conc. (ppm) | % Mosquito Landings | Proportion | SE | Lower CI | Upper CI | p |
|---|---|---|---|---|---|---|---|---|
| Cx. tarsalis | 30 °C | 2100 | 3.97 | 0.040 | 0.005 | 0.007 | 0.186 | 0.027 |
| 30,000 | 9.71 | 0.097 | 0.008 | 0.033 | 0.252 | 0.026 | ||
| 35 °C | 2100 | 3.7 | 0.037 | 0.005 | 0.007 | 0.184 | 0.027 | |
| 30,000 | 12.26 | 0.123 | 0.009 | 0.047 | 0.282 | 0.026 | ||
| 40 °C | 2100 | 3.8 | 0.038 | 0.005 | 0.007 | 0.184 | 0.027 | |
| 30,000 | 12.15 | 0.122 | 0.009 | 0.047 | 0.280 | 0.026 | ||
| 45 °C | 2100 | 3.49 | 0.035 | 0.005 | 0.006 | 0.182 | 0.027 | |
| 30,000 | 12.07 | 0.121 | 0.009 | 0.046 | 0.280 | 0.026 | ||
| 50 °C | 2100 | 3.47 | 0.035 | 0.005 | 0.006 | 0.183 | 0.028 | |
| 30,000 | 11.72 | 0.117 | 0.009 | 0.044 | 0.276 | 0.026 | ||
| Cx. quinquefasciatus | 30 °C | 2100 | 4.51 | 0.045 | 0.005 | 0.010 | 0.185 | 0.026 |
| 30,000 | 7.52 | 0.075 | 0.007 | 0.023 | 0.219 | 0.025 | ||
| 35 °C | 2100 | 5 | 0.050 | 0.006 | 0.012 | 0.191 | 0.026 | |
| 30,000 | 8.51 | 0.085 | 0.007 | 0.028 | 0.231 | 0.025 | ||
| 40 °C | 2100 | 5.16 | 0.052 | 0.006 | 0.012 | 0.192 | 0.025 | |
| 30,000 | 8.55 | 0.086 | 0.007 | 0.028 | 0.232 | 0.025 | ||
| 45 °C | 2100 | 5.2 | 0.052 | 0.006 | 0.012 | 0.193 | 0.025 | |
| 30,000 | 8.56 | 0.086 | 0.007 | 0.028 | 0.232 | 0.025 | ||
| 50 °C | 2100 | 5.13 | 0.051 | 0.006 | 0.012 | 0.192 | 0.025 | |
| 30,000 | 8.13 | 0.081 | 0.007 | 0.026 | 0.226 | 0.025 |
| Species | Duration (min) | CO2 Pulse | CO2 conc. (ppm) | % Mosquito Landings | Proportion | SE | Lower CI | Upper CI | p |
|---|---|---|---|---|---|---|---|---|---|
| Cx. tarsalis | 0–2 | Yes | 2100 | 0.06 | 0.001 | - | 0.000 | 1.000 | 0.999 |
| 30,000 | 10.72 | 0.107 | 0.009 | 0.034 | 0.290 | 0.029 | |||
| 2–4 | No | 2100 | 2.80 | 0.028 | 0.004 | 0.004 | 0.182 | 0.029 | |
| 30,000 | 22.16 | 0.222 | 0.014 | 0.090 | 0.450 | 0.042 | |||
| 4–6 | No | 2100 | 1.89 | 0.019 | 0.004 | 0.002 | 0.188 | 0.032 | |
| 30,000 | 17.86 | 0.179 | 0.014 | 0.062 | 0.415 | 0.039 | |||
| 6–8 | No | 2100 | 2.23 | 0.022 | 0.003 | 0.003 | 0.147 | 0.027 | |
| 30,000 | 13.21 | 0.132 | 0.012 | 0.038 | 0.372 | 0.036 | |||
| 8–10 | No | 2100 | 1.91 | 0.019 | 0.003 | 0.002 | 0.137 | 0.027 | |
| 30,000 | 9.61 | 0.096 | 0.011 | 0.020 | 0.355 | 0.037 | |||
| Cx. quinquefasciatus | 0–2 | Yes | 2100 | 3.72 | 0.037 | 0.005 | 0.008 | 0.165 | 0.025 |
| 30,000 | 6.82 | 0.068 | 0.007 | 0.018 | 0.228 | 0.027 | |||
| 2–4 | No | 2100 | 4.82 | 0.048 | 0.005 | 0.012 | 0.180 | 0.025 | |
| 30,000 | 13.37 | 0.134 | 0.010 | 0.048 | 0.322 | 0.030 | |||
| 4–6 | No | 2100 | 3.76 | 0.038 | 0.005 | 0.007 | 0.176 | 0.026 | |
| 30,000 | 11.56 | 0.116 | 0.010 | 0.038 | 0.303 | 0.030 | |||
| 6–8 | No | 2100 | 3.50 | 0.035 | 0.005 | 0.006 | 0.186 | 0.028 | |
| 30,000 | 8.97 | 0.090 | 0.009 | 0.025 | 0.272 | 0.029 | |||
| 8–10 | No | 2100 | 3.36 | 0.034 | 0.005 | 0.005 | 0.181 | 0.028 | |
| 30,000 | 7.48 | 0.075 | 0.008 | 0.018 | 0.261 | 0.029 |
| Species | Peltierwarm at 23 °C | CO2 Conc. (ppm) | % Mosquito Landings | Probability | SE | Lower CI | Upper CI | p |
|---|---|---|---|---|---|---|---|---|
| Cx. tarsalis | before 30 °C | 2100 | 0.83 | 0.008 | 0.002 | 0.001 | 0.120 | 0.029 |
| 30,000 | 0.00 | 0.000 | 0.000 | 0.000 | 1.000 | 1.000 | ||
| before 35 °C | 2100 | 1.17 | 0.012 | 0.002 | 0.001 | 0.111 | 0.027 | |
| 30,000 | 0.50 | 0.005 | 0.001 | 0.000 | 0.154 | 0.034 | ||
| before 40 °C | 2100 | 1.17 | 0.012 | 0.002 | 0.001 | 0.111 | 0.027 | |
| 30,000 | 4.92 | 0.049 | 0.004 | 0.016 | 0.143 | 0.020 | ||
| before 45 °C | 2100 | 1.50 | 0.015 | 0.002 | 0.002 | 0.109 | 0.025 | |
| 30,000 | 11.67 | 0.117 | 0.006 | 0.057 | 0.225 | 0.020 | ||
| before 50 °C | 2100 | 2.25 | 0.023 | 0.003 | 0.004 | 0.113 | 0.023 | |
| 30,000 | 10.33 | 0.103 | 0.006 | 0.048 | 0.209 | 0.019 | ||
| Cx. quinquefasciatus | before 30 °C | 2100 | 0.92 | 0.009 | 0.002 | 0.001 | 0.117 | 0.028 |
| 30,000 | 0.58 | 0.006 | 0.002 | 0.000 | 0.141 | 0.032 | ||
| before 35 °C | 2100 | 0.83 | 0.008 | 0.002 | 0.001 | 0.120 | 0.029 | |
| 30,000 | 0.83 | 0.008 | 0.002 | 0.001 | 0.120 | 0.029 | ||
| before 40 °C | 2100 | 3.17 | 0.032 | 0.003 | 0.008 | 0.122 | 0.021 | |
| 30,000 | 5.58 | 0.056 | 0.005 | 0.019 | 0.151 | 0.020 | ||
| before 45 °C | 2100 | 4.42 | 0.044 | 0.004 | 0.013 | 0.137 | 0.020 | |
| 30,000 | 11.92 | 0.119 | 0.006 | 0.058 | 0.228 | 0.020 | ||
| before 50 °C | 2100 | 6.08 | 0.061 | 0.005 | 0.022 | 0.158 | 0.019 | |
| 30,000 | 8.83 | 0.088 | 0.006 | 0.038 | 0.191 | 0.019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reinhold, J.M.; Chandrasegaran, K.; Oker, H.; Crespo, J.E.; Vinauger, C.; Lahondère, C. Species-Specificity in Thermopreference and CO2-Gated Heat-Seeking in Culex Mosquitoes. Insects 2022, 13, 92. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010092
Reinhold JM, Chandrasegaran K, Oker H, Crespo JE, Vinauger C, Lahondère C. Species-Specificity in Thermopreference and CO2-Gated Heat-Seeking in Culex Mosquitoes. Insects. 2022; 13(1):92. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010092
Chicago/Turabian StyleReinhold, Joanna M., Karthikeyan Chandrasegaran, Helen Oker, José E. Crespo, Clément Vinauger, and Chloé Lahondère. 2022. "Species-Specificity in Thermopreference and CO2-Gated Heat-Seeking in Culex Mosquitoes" Insects 13, no. 1: 92. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010092