
Changes in Biomass and Diversity of Soil Macrofauna along a Climatic Gradient in European Boreal Forests

 , , ,
, , ,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites

2.2. Collection and Identification of Invertebrates
2.3. Weighing of Invertebrates and Correction for Weight Loss during Preservation
2.4. Feeding Strategies of Soil Macrofauna
2.5. Statistical Analysis
3. Results
3.1. Overall Biomass and Species Richness

{kind=link}
{kind=link}
{kind=link}
| Class | Order | Family | No. of Species | No. of Individuals | Biomass, mg d.w. |
|---|---|---|---|---|---|
| Clitellata | Haplotaxida | Lumbricidae | 4 | 177 | 2647.0 |
| Arachnida | Araneae | Hahniidae | 2 | 68 | 13.1 |
| Linyphiidae | 47 | 499 | 165.7 | ||
| Lycosidae | 4 | 59 | 77.2 | ||
| Theridiidae | 3 | 219 | 85.9 | ||
| Thomisidae | 2 | 25 | 41.4 | ||
| Other 11 families | 16 | 49 | 54.1 | ||
| Diplopoda | Julida | Julidae | 1 | 11 | 256.2 |
| Polydesmida | Polydesmidae | 1 | 5 | 13.0 | |
| Polyzoniida | Polyzonidae | 1 | 3 | 5.0 | |
| Chilopoda | Lithobiomorpha | Lithobiidae | 2 | 222 | 218.3 |
| Insecta | Blattoptera | Ectobiidae | 1 | 5 | 7.3 |
| Hemiptera | Lygaeidae | 4 | 41 | 36.0 | |
| Other 6 families | 6 | 28 | 52.1 | ||
| Hymenoptera | Formicidae | 10 | 714 | 313.2 | |
| Pamphiliidae | 2 | 4 | 109.5 | ||
| Coleoptera | Carabidae | 14 | 53 | 168.7 | |
| Cantharidae | 8 | 94 | 127.8 | ||
| Curculionidae | 11 | 49 | 283.0 | ||
| Elateridae | 11 | 605 | 2353.9 | ||
| Staphylinidae | 72 | 478 | 292.1 | ||
| Other 7 families | 13 | 26 | 632.0 | ||
| Diptera | Chironomidae | 3 | 22 | 1.0 | |
| Rhagionidae | 3 | 94 | 223.7 | ||
| Other 12 families | 21 | 62 | 353.6 | ||
| Lepidoptera | Hepialidae | 2 | 22 | 339.2 | |
| Oecophoridae | 1 | 1 | 2.6 |
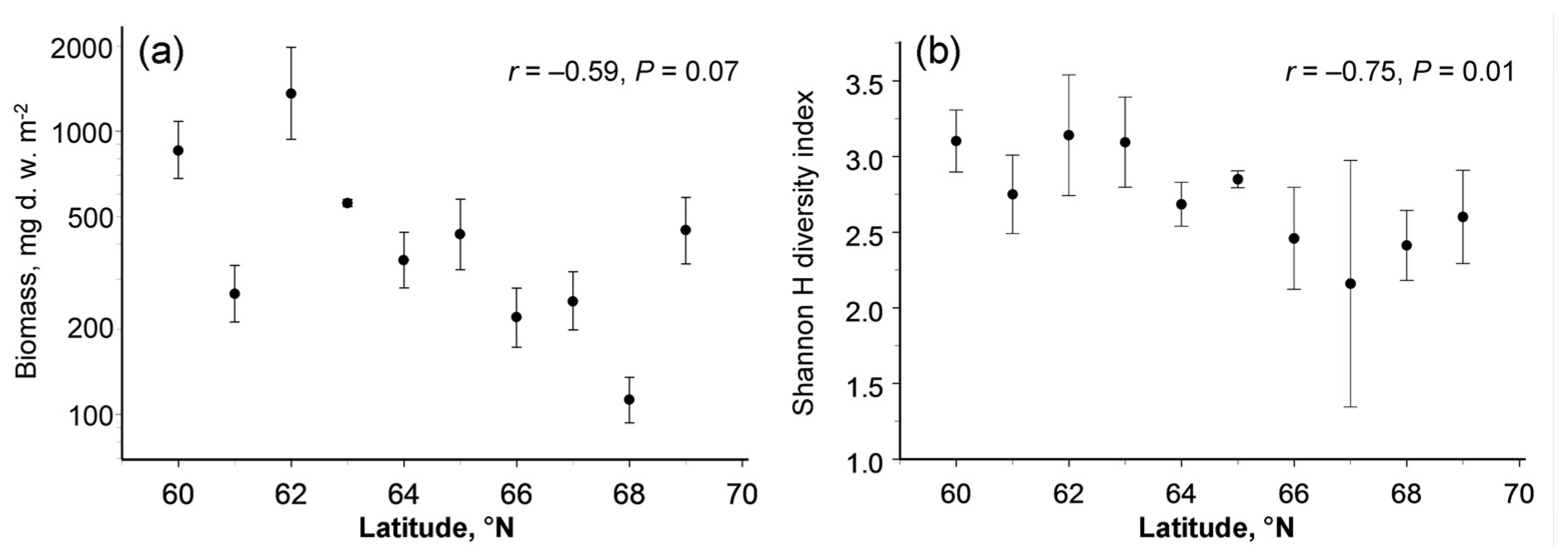
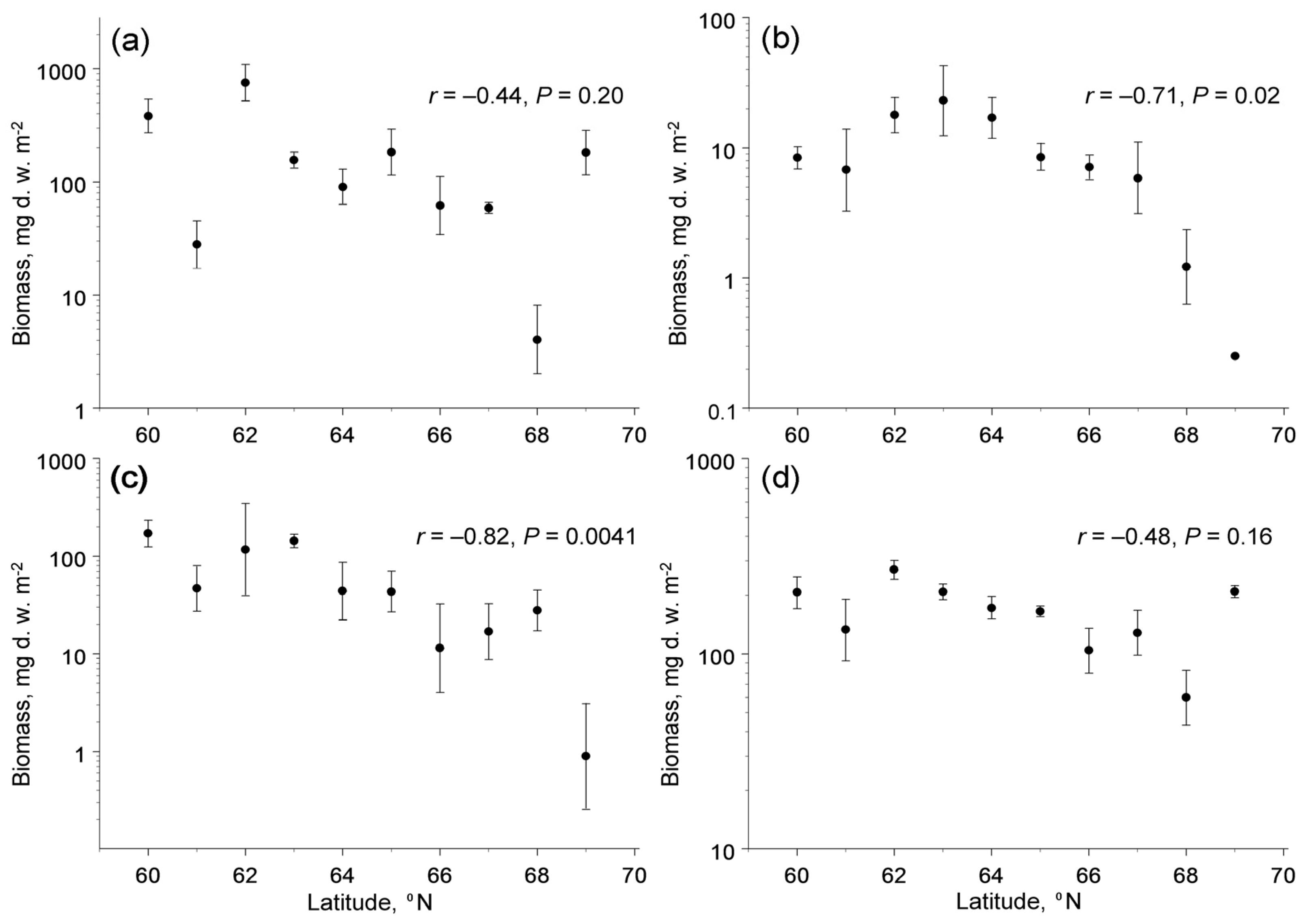
3.2. Latitudinal Patterns in Biomass
| Effect | Explanatory Variable | Biomass | Diversity | ||
|---|---|---|---|---|---|
| Test Statistics | p | Test Statistics | p | ||
| Fixed | Latitude | F1,8 = 4.31 | 0.07 | F1,8 = 10.58 | 0.0117 |
| Year | F1,24 = 2.65 | 0.12 | F1,24 = 3.45 | 0.08 | |
| Month | F1,24 = 8.83 | 0.0066 | F1,24 = 0.02 | 0.88 | |
| Latitude × Year | F1,24 = 2.80 | 0.11 | F1,24 = 3.65 | 0.07 | |
| Latitude × Month | F1,24 = 8.72 | 0.0069 | F1,24 = 0.09 | 0.77 | |
| Year × Month | F1,24 = 0.08 | 0.78 | F1,24 = 0.44 | 0.52 | |
| Latitude × Year × Month | F1,24 = 0.14 | 0.71 | F1,24 = 0.58 | 0.45 | |
| Random | Site | χ21 = 15.74 | <0.0001 | χ21 = 2.54 | 0.06 |


3.3. Latitudinal Pattern in Diversity
| Response Variable | Feeding Guild | Explanatory Variable | Slope | Standard Error | F | p | Partial R2 |
|---|---|---|---|---|---|---|---|
| Diversity | All combined | Latitude | −0.082 | 0.025 | 10.58 | 0.0117 | 0.570 |
| Biomass | All combined | Mean annual temperature | 0.105 | 0.040 | 6.14 | 0.0382 | 0.434 |
| Fine root biomass | −0.00082 | 0.00034 | 5.71 | 0.0482 | 0.254 | ||
| Fungivores | Latitude | −0.126 | 0.026 | 8.26 | 0.0207 | 0.508 | |
| Potassium in soil | −0.00148 | 0.00032 | 20.87 | 0.0026 | 0.368 | ||
| Herbivores | Latitude | −0.1668 | 0.025 | 15.84 | 0.0041 | 0.664 | |
| Potassium in soil | −0.00137 | 0.00031 | 19.65 | 0.0030 | 0.247 |
4. Discussion
4.1. Latitudinal Changes in Diversity and Biomass of Soil Macrofauna
4.2. Latitudinal Changes in Biotic Interactions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barrios, E. Soil biota, ecosystem services and land productivity. Ecol. Econ. 2007, 64, 269–285. [Google Scholar] [CrossRef]
- Gobat, J.; Aragno, M.; Matthey, W. The Living Soil: Fundamentals of Soil Science and Soil Biol; Science Publishers: Enfield, CT, USA, 2004. [Google Scholar]
- Brussaard, L. Ecosystem services provided by the soil biota. In Soil Ecology and Ecosystem Services; Wall, D.H., Ed.; Oxford University Press: Oxford, MS, USA, 2012; pp. 45–58. [Google Scholar]
- Adhikari, K.; Hartemink, A.E. Linking soils to ecosystem services—A global review. Geoderma 2016, 262, 101–111. [Google Scholar] [CrossRef]
- Copley, J. Ecology goes underground. Nature 2000, 406, 452–454. [Google Scholar] [CrossRef] [PubMed]
- Blois, J.L.; Zarnetske, P.L.; Fitzpatrick, M.C.; Finnegan, S. Climate change and the past, present, and future of biotic interactions. Science 2013, 341, 499–504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coyle, D.R.; Nagendra, U.J.; Taylor, M.K.; Campbell, J.H.; Cunard, C.E.; Joslin, A.H.; Callaham, M.A., Jr. Soil fauna responses to natural disturbances, invasive species, and global climate change: Current state of the science and a call to action. Soil Biol. Biochem. 2017, 110, 116–133. [Google Scholar] [CrossRef]
- Blankinship, J.C.; Niklaus, P.A.; Hungate, B.A. A meta-analysis of responses of soil biota to global change. Oecologia 2011, 165, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.T.; Kharouba, H.M.; Currie, D.J. The macroecological contribution to global change solutions. Science 2007, 316, 1581–1584. [Google Scholar] [CrossRef] [Green Version]
- De Frenne, P.; Graae, B.J.; Rodríguez-Sánchez, F.; Kolb, A.; Chabrerie, O.; Verheyen, K. Latitudinal gradients as natural laboratories to infer species’ responses to temperature. J. Ecol. 2013, 101, 784–795. [Google Scholar] [CrossRef]
- Zvereva, E.L.; Kozlov, M.V. Latitudinal gradient in the intensity of biotic interactions in terrestrial ecosystems: Sources of variation and differences from the diversity gradient revealed by meta-analysis. Ecol. Lett. 2021, 24, 2506–2520. [Google Scholar] [CrossRef]
- Fierer, N.; Strickland, M.S.; Liptzin, D.; Bradford, M.A.; Cleveland, C.C. Global patterns in belowground communities. Ecol. Lett. 2009, 12, 1238–1249. [Google Scholar] [CrossRef]
- Peng, Y.; Guo, D.; Yang, Y. Global patterns of root dynamics under nitrogen enrichment. Glob. Ecol. Biogeogr. 2017, 26, 102–114. [Google Scholar] [CrossRef]
- Phillips, H.R.P.; Guella, C.A.; Bartz, M.L.C.; Briones, M.J.I.; Brown, G.; Crowther, T.W.; Eisenhauer, N. Global distribution of earthworm diversity. Science 2019, 366, 480–485. [Google Scholar] [CrossRef] [Green Version]
- Johnston, A.S.A.; Sibly, R.M. Multiple environmental controls explain global patterns in soil animal communities. Oecologia 2020, 192, 1047–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Petersen, H.; Luxton, M. A comparative analysis of soil fauna populations and their role in decomposition processes. Oikos 1982, 39, 288–388. [Google Scholar] [CrossRef]
- Gongalsky, K.B. Soil macrofauna: Study problems and perspectives. Soil Biol. Biochem. 2021, 159, 108281. [Google Scholar] [CrossRef]
- Bottinelli, N.; Jouquet, P.; Capowiez, Y.; Podwojewski, P.; Grimaldi, M.; Peng, X. Why is the influence of soil macrofauna on soil structure only considered by soil ecologists? Soil Tillage Res. 2015, 146, 118–124. [Google Scholar] [CrossRef]
- Potapov, A.M.; Tiunov, A.V.; Scheu, S. Uncovering trophic positions and food resources of soil animals using bulk natural stable isotope composition. Biol. Rev. 2019, 94, 37–59. [Google Scholar] [CrossRef]
- Voight, W.; Perner, J.; Davis, A.J.; Eggers, T.; Schumacher, J.; Bährmann, R.; Fabian, B.; Heinrich, W.; Köhler, G.; Lichter, D.; et al. Trophic levels are differentially sensitive to climate. Ecology 2003, 84, 2444–2453. [Google Scholar] [CrossRef] [Green Version]
- Holland, M.M.; Smith, J.A.; Everett, J.D.; Verges, A.; Suthers, I.M. Latitudinal patterns in trophic structure of temperate reef-associated fishes and predicted consequences of climate change. Fish Fish. 2020, 21, 1092–1108. [Google Scholar] [CrossRef]
- Cameron, E.K.; Martins, I.S.; Lavelle, P.; Mathieu, J.; Tedersoo, L.; Gottschall, F.; Eisenhauter, N. Global gaps in soil biodiversity data. Nature Ecol. Evol. 2018, 2, 1042–1043. [Google Scholar] [CrossRef]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.T.; Bezemer, M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Change Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- McNaughton, S.J.; Oesterheld, M.; Frank, D.A.; Williams, K.J. Ecosystem-level patterns of primary productivity and herbivory in terrestrial habitats. Nature 1989, 341, 142–144. [Google Scholar] [CrossRef]
- Huston, M.A.; Wolverton, S. The global distribution of net primary production: Resolving the paradox. Ecol. Monogr. 2009, 79, 343–377. [Google Scholar] [CrossRef]
- Zvereva, E.L.; Zverev, V.; Usoltsev, V.A.; Kozlov, M.V. Latitudinal pattern in community-wide herbivory does not match the pattern in herbivory averaged across common plant species. J. Ecol. 2020, 108, 2511–2520. [Google Scholar] [CrossRef]
- Finér, L.; Zverev, V.; Palviainen, M.; Romanis, T.; Kozlov, M.V. Variation in fine root biomass along a 1000 km long latitudinal climatic gradient in mixed boreal forests. Forest Ecol. Manage 2019, 432, 649–655. [Google Scholar] [CrossRef]
- Xu, X.F.; Thornton, P.E.; Post, W.M. A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Glob. Ecol. Biogeogr. 2013, 22, 737–749. [Google Scholar] [CrossRef]
- FAO. World Reference Base for Soil Resources. In International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Gilyarov, M.S. Methods in Soil Zoology; Nauka: Moscow, Russia, 1975. (In Russian) [Google Scholar]
- deWaard, J.R.; Ivanova, N.V.; Hajibabaei, M.; Hebert, P.D.N. Assembling DNA barcodes: Analytical protocols. In Methods in Molecular Biology: Environmental Genetics; Cristofre, M., Ed.; Humana Press Inc.: Totowa, NJ, USA, 2008; pp. 275–293. [Google Scholar]
- Leuven, R.S.E.W.; Brock, T.C.M.; van Druten, H.A.M. Effects of preservation on dry- and ash-free dry weight biomass of some common aquatic macro-invertebrates. Hydrobiologia 1985, 127, 151–159. [Google Scholar] [CrossRef]
- Wetzel, M.A.; Leuchs, H.; Koop, J.H.E. Preservation effects on wet weight, dry weight, and ash-free dry weight biomass estimates of four common estuarine macro-invertebrates: No difference between ethanol and formalin. Helgol. Mar. Res. 2005, 59, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Finér, L.; Jurgensen, M.F.; Domisch, T.; Kilpeläinen, J.; Neuvonen, S.; Punttila, P.; Risch, A.C.; Ohashi, M.; Niemelä, P. The role of wood ants (Formica rufa group) in carbon and nutrient dynamics of a boreal Norway spruce forest ecosystem. Ecosystems 2013, 16, 196–208. [Google Scholar] [CrossRef]
- Dolin, V.G. On trophic links of click beetle larvae (wireworms). In Materials to the Investigation of Fauna and Ecology of Insects in Central Parts of Forest-Steppes of Ukraine; Kryshtal, A.F., Ed.; Kiev State University: Kiev, Russia, 1963; pp. 116–147. (In Russian) [Google Scholar]
- Kozlov, M.V.; Prosvirov, A.S.; Zvereva, E.L. Can larvae of forest click beetles (Coleoptera: Elateridae) feed on live plant roots? Insects 2020, 11, 850. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeont. Electron. 2001, 4, 1–9. [Google Scholar]
- SAS Institute. SAS/Stat. User’s Guide, Version 9.2; SAS Institute: Cary, NC, USA, 2009.
- Kenward, M.G.; Roger, J.H. An improved approximation to the precision of fixed effects from restricted maximum likelihood. Comput. Stat. Data Anal. 2009, 53, 2583–2595. [Google Scholar] [CrossRef]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Schabenberger, O. SAS for Mixed Models, 2nd ed.; SAS Institute: Cary, NC, USA, 2006. [Google Scholar]
- Kozlov, M.V.; Stańska, M.; Hajdamowicz, I.; Zverev, V.; Zvereva, E.L. Abundance and diversity of arboreal spiders along latitudinal gradients in northern Europe. Ecography 2015, 38, 1026–1035. [Google Scholar] [CrossRef]
- Kinlock, N.L.; Prowant, L.; Herstoff, E.M.; Foley, C.M.; Akin-Fajiye, M.; Bender, N.; Umarani, M.; Ryu, H.Y.; Şen, B.; Gurevitch, J.; et al. Explaining global variation in the latitudinal diversity gradient: Meta-analysis confirms known patterns and uncovers new ones. Glob. Ecol. Biogeogr. 2018, 27, 125–141. [Google Scholar] [CrossRef]
- Wu, T.; Ayres, E.; Bardgett, R.D.; Wall, D.H.; Garey, J.R. Molecular study of worldwide distribution and diversity of soil animals. Proc. Nat. Acad. Sci. USA 2011, 108, 17720–17725. [Google Scholar] [CrossRef] [Green Version]
- Cameron, E.K.; Martins, I.S.; Lavelle, P.; Mathieu, J.; Tedersoo, L.; Bahram, M.; Eisenhauter, N. Global mismatches in aboveground and belowground biodiversity. Conserv. Biol. 2019, 33, 1187–1192. [Google Scholar] [CrossRef]
- Hillebrand, H. On the generality of the latitudinal diversity gradient. Am. Nat. 2004, 163, 192–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlov, M.V.; van Nieukerken, E.J.; Zverev, V.; Zvereva, E.L. Abundance and diversity of birch-feeding leafminers along latitudinal gradients in Northern Europe. Ecography 2013, 36, 1138–1149. [Google Scholar] [CrossRef]
- Kanervo, J.; Kozlov, M.V. Diversity and abundance of arboreal psocids (Insecta: Psocodea: ‘Psocoptera’) along latitudinal gradients in northern Europe. Eur. J. Entomol. 2014, 111, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.Y.; Chen, Y.H. Fine root biomass, production, turnover rates, and nutrient contents in boreal forest ecosystems in relation to species, climate, fertility, stand age: Literature review and meta-analyses. Crit. Rev. Plant Sci. 2010, 29, 204–221. [Google Scholar] [CrossRef]
- Jacob, D.; Petersen, J.; Eggert, B.; Alias, A.; Christensen, O.B.; Yiou, P. EURO-CORDEX: New high-resolution climate change projections for European impact research. Region. Environ. Chang. 2014, 14, 563–578. [Google Scholar] [CrossRef]
- Odum, E.P.; Barrett, G.W. Fundamentals of Ecology; Saunders: Philadelphia, PA, USA, 1971; Volume 3. [Google Scholar]
- Hairston, N.G., Jr.; Hairston, N.G., Sr. Cause-effect relationships in energy flow, trophic structure, and interspecific interactions. Am. Nat. 1993, 142, 379–411. [Google Scholar] [CrossRef]
- Andrew, N.R.; Hughes, L. Arthropod community structure along a latitudinal gradient: Implications for future impacts of climate change. Austral Ecol. 2005, 30, 281–297. [Google Scholar] [CrossRef]
- de Sassi, C.; Tylianakis, J.M. Climate change disproportionately increases herbivore over plant or parasitoid biomass. PLoS ONE 2012, 7, e40557. [Google Scholar] [CrossRef]
- Zvereva, E.L.; Zverev, V.; Kozlov, M.V. Predation and parasitism on herbivorous insects change in opposite directions in a latitudinal gradient crossing a boreal forest zone. J. Anim. Ecol. 2020, 89, 2946–2957. [Google Scholar] [CrossRef]
- Lemoine, N.P.; Burkepile, D.E.; Parker, J.D. Variable effects of temperature on insect herbivory. PeerJ 2014, 2, e376. [Google Scholar] [CrossRef] [Green Version]
- Yli-Halla, M. Soil temperature regimes in Finland. Agricult. Food Sci. Finland 1998, 7, 507–512. [Google Scholar] [CrossRef]
- Schemske, D.W.; Mittelbach, G.G.; Cornell, H.V.; Sobel, J.M.; Roy, K. Is there a latitudinal gradient in the importance of biotic interactions? Ann. Rev. Ecol. Evol. Syst. 2009, 40, 245–269. [Google Scholar] [CrossRef] [Green Version]
- Blois, J.L.; Williams, J.W.; Fitzpatrick, M.C.; Jackson, S.T.; Ferrier, S. Space can substitute for time in predicting climate-change effects on biodiversity. Proc. Nat. Acad. Sci. USA 2013, 110, 9374–9379. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kozlov, M.V.; Zverev, V.; Gusarov, V.I.; Korobushkin, D.I.; Krivosheina, N.P.; Mattila, J.; Mutanen, M.; Popova, A.; Prosvirov, A.S.; Punttila, P.; et al. Changes in Biomass and Diversity of Soil Macrofauna along a Climatic Gradient in European Boreal Forests. Insects 2022, 13, 94. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010094
Kozlov MV, Zverev V, Gusarov VI, Korobushkin DI, Krivosheina NP, Mattila J, Mutanen M, Popova A, Prosvirov AS, Punttila P, et al. Changes in Biomass and Diversity of Soil Macrofauna along a Climatic Gradient in European Boreal Forests. Insects. 2022; 13(1):94. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010094
Chicago/Turabian StyleKozlov, Mikhail V., Vitali Zverev, Vladimir I. Gusarov, Daniil I. Korobushkin, Nina P. Krivosheina, Jaakko Mattila, Marko Mutanen, Anna Popova, Alexander S. Prosvirov, Pekka Punttila, and et al. 2022. "Changes in Biomass and Diversity of Soil Macrofauna along a Climatic Gradient in European Boreal Forests" Insects 13, no. 1: 94. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13010094