
Organophosphate Insecticides Resistance in Field Populations of House Flies, Musca domestica L.: Levels of Resistance and Acetylcholinesterase Activity

 ,
, 
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Insects
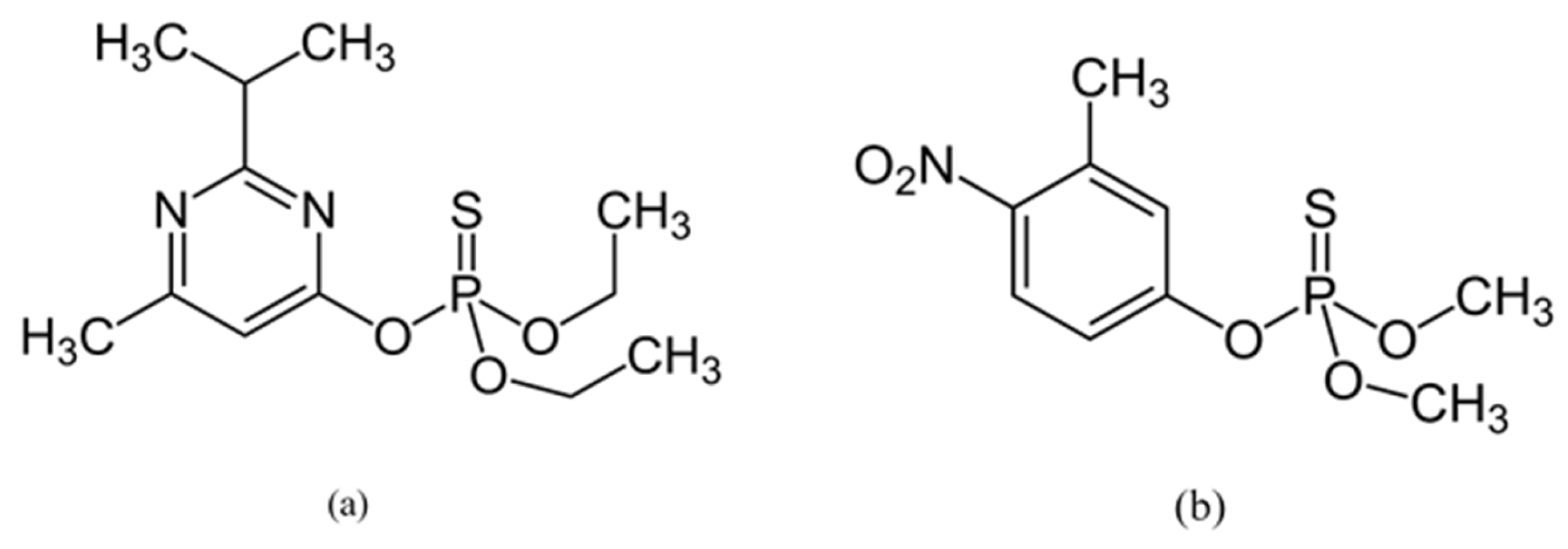
2.2. Insecticides
2.3. Bioassays
2.4. AChE Activity Assays
2.5. Statistical Analysis
3. Results
3.1. Susceptibility of House Flies to Diazinon and Fenitrothion
3.2. Activity of AChE and Inhibitory Effect of Diazinon and Fenitrothion
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Greenberg, B. Flies and Disease; Princeton University Press: Princeton, NJ, USA, 1971; p. 856. [Google Scholar]
- Kasprzak, W.; Majewska, A. Transmission of Giardia cysts. I. Role of flies and cockroaches. Parasitol. News 1981, 27, 555–563. [Google Scholar]
- Graczyk, T.K.; Knight, R.; Gilman, R.H.; Cranfield, M.R. The role of non-biting flies in the epidemiology of human infectious diseases. Microbes Infect. 2001, 3, 231–235. [Google Scholar] [CrossRef]
- Ugbogu, O.C.; Nwachukwu, N.C.; Obguagu, U.N. Isolation of Salmonella and Shigella species from house flies (Musca domestica L.) in Uturu, Nigeria. Afr. J. Biotechnol. 2006, 5, 1090–1091. [Google Scholar]
- Moon, R.D. Muscid flies (Muscidae). In Medical and Veterinary Entomology; Mullen, G.R., Durden, L.A., Eds.; Academic Press: New York, NY, USA, 2009; pp. 275–295. [Google Scholar]
- Pinto, M.; Prado, A. Resistance of Musca domestica L. populations to cyromazine (insect growth regulator) in Brazil. Mem. Inst. Oswaldo Cruz 2001, 96, 729–732. [Google Scholar] [CrossRef]
- Marçon, P.C.R.G.; Thomas, G.D.; Siegfried, B.D.; Campbell, J.B.; Skoda, S.R. Resistance status of house flies (Diptera: Muscidae) from southeastern Nebraska beef cattle feedlots to selected insecticides. J. Econ. Entomol. 2003, 96, 1016–1020. [Google Scholar] [CrossRef]
- Karunamoorthi, K.; Sabesan, S. Insecticide resistance in insect vectors of disease with special reference to mosquitoes: A potential threat to global public health. Health Scope 2013, 2, 4–18. [Google Scholar] [CrossRef] [Green Version]
- Plapp, F.W. The genetic basis of insecticide resistance in the house fly: Evidence that a single locus plays a major role in metabolic resistance to insecticides. Pestic. Biochem. Physiol. 1984, 22, 194–201. [Google Scholar] [CrossRef]
- Metcalf, R.L. Insect resistance to insecticides. Pestic. Sci. 1989, 26, 333–358. [Google Scholar] [CrossRef]
- Scott, J.G.; Alefantis, T.G.; Kaufman, P.E.; Rutz, D.A. Insecticide resistance in house flies from caged-layer poultry facilities. Pest Manag. Sci. 2000, 56, 147–153. [Google Scholar]
- Kristensen, M.; Spencer, A.G.; Jespersen, J.B. The status and development of insecticide resistance in Danish populations of the house fly Musca domestica L. Pest Manag. Sci. 2001, 57, 82–89. [Google Scholar] [CrossRef]
- Alzahrani, S.M.; Ajlan, A.; Hajjar, M.J.; Albokari, M. Resistance of field strains of house fly Musca domestica L. to diazinon insecticide in Riyadh city, Saudi Arabia. J. Jazan Univ. Appl. Sci. Branch 2015, 4, 1–8. [Google Scholar]
- Hafez, A.M. First Evaluation of Field Evolved Resistance to Commonly Used Insecticides in House Fly Populations from Saudi Arabian Dairy Farms. Insects 2021, 12, 1120. [Google Scholar] [CrossRef]
- Lim, L.; Bolstad, H.M. Organophosphate insecticides: Neurodevelopmental effects. In Encyclopedia of Environmental Health; Nriagu, J., Ed.; Elsevier: London, UK, 2019; pp. 785–791. [Google Scholar]
- Nishizawa, Y.; Fujii, K.; Kadota, T.; Miyamoto, J.; Sakamoto, H. Studies on organophosphorus insecticides part VII. Chemical and biological properties of new low toxic organophosphorus insecticide, O,O-dimiethyl-O-(3-methyl-4-nitrophenyl) phosphorothioate. Agric. Biol. Chem. 1961, 25, 605–610. [Google Scholar] [CrossRef] [Green Version]
- Eto, M. Organophosphorus Pesticides: Organic and Biological Chemistry; CRC Press: Cleveland, OH, USA, 1974; p. 387. [Google Scholar]
- Oppenoorth, F.J.; Welling, W. Biochemistry and physiology of resistance. In Insecticide Biochemistry and Physiology; Wilkinson, C.F., Ed.; Plenum Press: New York, NY, USA, 1979; pp. 507–551. [Google Scholar]
- Escartín, E.; Porte, C. Bioaccumulation, metabolism, and bio-chemical effects of the organophosphorus pesticide fenitrothion in Procambarus Clarkii. Environ. Toxicol. Chem. 1996, 15, 915–920. [Google Scholar] [CrossRef]
- Barata, C.; Solayan, A.; Porte, C. Role of B-esterases in assessing toxicity of organophosphorus (chlorpyrifos, malathion) and carbamate (carbofuran) pesticides to Daphnia magna. Aquat. Toxicol. 2004, 66, 125–139. [Google Scholar] [CrossRef]
- Siegfried, B.D.; Scharf, M.E. Mechanisms of organophosphate resistance in insects. In Biochemical Sites of Insecticide Action and Resistance; Ishaaya, I., Ed.; Springer: Berlin, Heidelberg, 2001. [Google Scholar] [CrossRef]
- Roush, R.T.; Mckenzie, J.A. Ecological genetics of insecticide and acaricide resistance. Annu. Rev. Entomol. 1987, 32, 361–380. [Google Scholar] [CrossRef]
- Taylor, M.; Feyereisen, R. Molecular biology and evolution of resistance to toxicants. Mol. Biol. Evol. 1996, 13, 719–734. [Google Scholar] [CrossRef] [Green Version]
- The Saudi Food and Drug Authority (SFDA). Database for Pesticide Registration. Available online: https://www.sfda.gov.sa/en/informationlist/65748 (accessed on 17 October 2021).
- Ellman, G.L.; Courtney, K.D.; Anders, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Abbott, W. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis; Cambridge University Press: New York, NY, USA, 1971; p. 333. [Google Scholar]
- Litchfield, J.; Wilcoxon, F. A simplified method of evaluating dose-effect experiments. J. Exp. Ther. 1949, 96, 99–110. [Google Scholar]
- Valles, S.M.; Koehler, P.G.; Brenner, R.J. Antagonism of fipronil toxicity by piperonyl butoxide and S, S, S-tributyl phosphorotrithioate in the German cockroach (Dictyoptera: Blattellidae). J. Econ. Entomol. 1997, 90, 1254–1258. [Google Scholar] [CrossRef]
- Akiner, M.M.; Caglar, S.S. The status and seasonal changes of organophosphate and pyrethroid resistance in Turkish populations of the house fly, Musca domestica L. (Diptera: Muscidae). J. Vector Ecol. 2006, 31, 58–64. [Google Scholar] [CrossRef]
- Acevedo, G.R.; Zapater, M.; Toloza, A.C. Insecticide resistance of house fly, Musca domestica (L.) from Argentina. Parasitol. Res. 2009, 105, 489–493. [Google Scholar] [CrossRef]
- Khan, H.A.A.; Akram, W.; Shad, S.A. Resistance to conventional insecticides in Pakistani populations of Musca domestica L. (Diptera: Muscidae): A potential ectoparasite of dairy animals. Ecotoxicology 2013, 22, 522–527. [Google Scholar] [CrossRef]
- Keiding, J. Resistance to fenitrothion in Danish house flies. Musca domestica. J. Hyg. Epidemiol. Microbiol. Immunol. 1976, 20, 171–185. [Google Scholar]
- Ugaki, M.; Shono, T.; Fukami, J.I. Metabolism of fenitrothion by organophosphorus-resistant and susceptible house flies, Musca domestica L. Pestic. Biochem. Physiol. 1985, 23, 33–40. [Google Scholar] [CrossRef]
- Pospischil, R.; Szomm, K.; Londershausen, M.; Schröder, I.; Turberg, A.; Fuchs, R. Multiple resistance in the larger house fly Musca domestica in Germany. Pestic Sci. 1996, 48, 333–341. [Google Scholar] [CrossRef]
- Akiner, M.M.; Çaglar, S.S. Monitoring of five different insecticide resistance status in Turkish house fly Musca domestica L. (Diptera: Muscidae) populations and the relationship between resistance and insecticide usage profile. Türkiye Parazitolojii Dergisi 2012, 36, 87–91. [Google Scholar] [CrossRef]
- Pai, H.; Lin, K.; Wang, S.; Hsu, E. Resistance to 10 common insecticides of house flies (Musca domestica) from garbage dumps in Taiwan. Int. J. Sci. Res. 2020, 9, 1–5. [Google Scholar]
- Tabashnik, B.E.; Cushing, N.L.; Johnson, M.W. Diamondback moth (Lepidoptera: Plutellidae) resistance to insecticides in Hawaii: Intra-island variation and cross-resistance. J Econ. Entomol 1987, 80, 1091–1099. [Google Scholar] [CrossRef]
- Wang, J.N.; Hou, J.; Wu, Y.Y.; Guo, S.; Liu, Q.M.; Li, T.Q.; Gong, Z.Y. Resistance of house fly, Musca domestica L.(Diptera: Muscidae), to five insecticides in Zhejiang Province, China: The situation in 2017. Can. J. Infect. Dis. Med. Microbiol. 2019, 2019, e4851914. [Google Scholar] [CrossRef] [Green Version]
- Alzahrani, S.M. Evaluation of triflumuron and pyriproxyfen as alternative candidates to control house fly, Musca domestica L. (Diptera: Muscidae), in Riyadh city, Saudi Arabia. PloS ONE 2021, 16, e0249496. [Google Scholar] [CrossRef]
- Fournier, D.; Mutero, A. Modification of acetylcholinesterase as a mechanism of resistance to insecticides. Comp. Biochem. Physiol. 1994, 108, 19–31. [Google Scholar] [CrossRef]
- Plapp, F.W.; Tripathi, R.K. Biochemical genetics of altered acetylcholinesterase resistance to insecticides in the house fly. Biochem. Genet. 1978, 16, 1–11. [Google Scholar] [CrossRef]
- Kristensen, M.; Huang, J.; Qiao, C.L.; Jespersen, J.B. Variation of Musca domestica L. acetylcholinesterase in Danish housefly populations. Pest Manag. Sci. 2006, 62, 738–745. [Google Scholar] [CrossRef]
- Tripathi, R.K.; O’Brien, R.D. Insensitive acetylcholinesterase as a factor in resistance of house flies to the organophosphate Rabon. Pestic. Biochem. Physiol. 1973, 3, 495–498. [Google Scholar] [CrossRef]
- Devonshire, A.L. Studies of the acetylcholinesterase from house flies (Musca domestica L.) resistant and susceptible to organophosphorus insecticides. Biochem. J. 1975, 149, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Devonshire, A.L.; Moores, G.D. Different forms of insensitive acetylcholinesterase in insecticide-resistant house flies (Musca domestica). Pestic. Biochem. Physiol. 1984, 21, 336–340. [Google Scholar] [CrossRef]
- Kim, W.T.; Boo, K.S. Insecticide sensitivity of acetylcholinesterase from a Korean Housefly (Musca domestica) strain to organophosphates. J. Asia-Pacific Entomol. 2004, 7, 187–193. [Google Scholar] [CrossRef]
- Kozaki, T.; Shono, T.; Tomita, T.; Kono, Y. Fenitroxon insensitive acetylcholinesterases of the house fly, Musca domestica associated with point mutations. Insect Biochem. Molec. Biol. 2001, 31, 991–997. [Google Scholar] [CrossRef]
- Yuan, J.Z.; Sun, X.Q.; Tang, Z.H.; Tao, L.M. Study on biochemical properties of acetylcholinesterase from propoxur-resistant and susceptible strains of the house fly Musca domestica. Chin. J. Pestic. Sci. 2009, 11, 181–185. [Google Scholar]
- Tao, L.M.; Shi, M.A.; Yuang, J.Z.; Zhuang, P.J.; Zhang, C.X.; Tang, Z.H. Resistance pattern and point mutations of insensitive acetylcholinesterase in a carbamate-resistant strain of house fly (Musca domestica). Pestic. Biochem. Physiol. 2006, 86, 1–6. [Google Scholar]


{kind=link}
{kind=link}
| Site Name | Activity | Coordinates | |
|---|---|---|---|
| Latitude | Longitude | ||
| Sa’ada (SAS) | Slaughterhouse | 24.70812 N | 46.85459 E |
| Mowanneseiah (MOS) | Slaughterhouse | 24.82754 N | 46.79580 E |
| North (NOS) | Slaughterhouse | 24.75572 N | 46.66544 E |
| West (WES) | Slaughterhouse | 24.56944 N | 46.50776 E |
| Mansoureiah (MAS) | Slaughterhouse | 24.58842 N | 46.73466 E |
| Azizeiah (AZM) | Wholesale market | 24.59261 N | 46.74299 E |
| Badea’ah (BAM) | Wholesale market | 24.58099 N | 46.61288 E |
| Rabwah (RAM) | Wholesale market | 24.69418 N | 46.77861 E |
| Population | N | LD50 (ng/♀) | 95% Confidence Limits | Slope ± SE | χ2 | p | RR | |
|---|---|---|---|---|---|---|---|---|
| Lower | Upper | |||||||
| LAB | 360 | 3.27 a | 2.83 | 3.75 | 2.45 ± 0.24 | 5.02 | 0.17 | - |
| RAM | 360 | 204.28 b | 163.02 | 249.74 | 1.66 ± 0.17 | 8.79 | 0.03 | 62.47 |
| NOS | 360 | 348.22 c | 275.56 | 419.74 | 2.08 ± 0.24 | 7.63 | 0.05 | 106.48 |
| BAM | 360 | 392.02 c | 323.02 | 496.93 | 1.95 ± 0.25 | 7.20 | 0.06 | 119.88 |
| AZM | 360 | 405.86 c | 333.63 | 489.53 | 1.89 ± 0.25 | 5.48 | 0.14 | 124.11 |
| WES | 360 | 452.48 c | 381.06 | 529.63 | 2.06 ± 0.23 | 5.90 | 0.12 | 138.37 |
| MAS | 360 | 892.26 d | 835.11 | 949.11 | 5.53 ± 0.53 | 3.57 | 0.31 | 272.86 |
| SAS | 360 | 960.38 de | 896.48 | 1027.58 | 4.97 ± 0.51 | 3.22 | 0.31 | 293.69 |
| MOS | 360 | 1013.01 e | 958.29 | 1070.32 | 6.50 ± 0.61 | 2.93 | 0.40 | 309.78 |
| Population | N | LD50 (ng/♀) | 95% Confidence Limits | Slope ± SE | χ2 | p | RR | |
|---|---|---|---|---|---|---|---|---|
| Lower | Upper | |||||||
| LAB | 360 | 4.99 a | 4.02 | 6.09 | 1.71 ± 0.16 | 0.59 | 0.90 | - |
| RAM | 360 | 264.88 b | 220.71 | 314.25 | 2.01 ± 0.18 | 6.68 | 0.08 | 53.08 |
| AZM | 360 | 391.30 c | 322.56 | 481.40 | 1.68 ± 0.18 | 8.67 | 0.03 | 78.41 |
| BAM | 360 | 481.24 c | 416.43 | 549.59 | 2.52 ± 0.27 | 9.48 | 0.02 | 96.44 |
| MOS | 360 | 971.15 d | 900.00 | 1047.99 | 4.37 ± 0.48 | 1.00 | 0.80 | 194.61 |
| MAS | 360 | 1071.92 de | 996.52 | 1161.06 | 4.49 ± 0.51 | 5.98 | 0.11 | 214.81 |
| NOS | 360 | 1101.06 de | 1037.78 | 1173.12 | 5.85 ± 0.60 | 4.44 | 0.22 | 220.65 |
| SAS | 360 | 1143.56 ef | 1074.29 | 1226.54 | 5.51 ± 0.59 | 0.72 | 0.87 | 229.17 |
| WES | 360 | 1303.60 f | 1212.77 | 1431.61 | 5.18 ± 0.63 | 0.91 | 0.82 | 261.24 |
| Population | Specific Activity (µmol/min/mg Protein) | SD | p-Value |
|---|---|---|---|
| LAB | 0.161 | 0.001 | |
| RAM | 0.173 | 0.0008 | 0.0004 |
| AZM | 0.194 | 0.0004 | <0.00001 |
| BAM | 0.188 | 0.0007 | 0.00001 |
| MOS | 0.180 | 0.0001 | 0.00004 |
| MAS | 0.193 | 0.0006 | <0.00001 |
| NOS | 0.191 | 0.002 | 0.00002 |
| SAS | 0.191 | 0.0009 | 0.00001 |
| WES | 0.192 | 0.005 | 0.0003 |
| Population | N | IC50 (µg/mL) | 95% Confidence Limits | Slope ± SE | χ2 | p | IR | |
|---|---|---|---|---|---|---|---|---|
| Lower | Upper | |||||||
| LAB | 90 | 0.77 a | 0.60 | 0.97 | 1.27 ± 0.19 | 0.92 | 0.82 | - |
| WES | 90 | 48.34 b | 31.38 | 63.42 | 1.53 ± 0.23 | 1.31 | 0.73 | 62.77 |
| RAM | 90 | 63.91 bc | 40.43 | 85.03 | 1.19 ± 0.20 | 3.42 | 0.33 | 83.00 |
| AZM | 90 | 64.91 bc | 48.02 | 80.44 | 1.66 ± 0.21 | 0.43 | 0.93 | 84.29 |
| BAM | 90 | 74.05 bcd | 53.85 | 92.91 | 1.44 ± 0.20 | 1.76 | 0.62 | 96.16 |
| NOS | 90 | 79.41 bcd | 60.23 | 97.69 | 1.55 ± 0.20 | 0.15 | 0.98 | 103.12 |
| SAS | 90 | 102.26 cd | 77.29 | 128.13 | 1.30 ± 0.19 | 4.64 | 0.20 | 132.80 |
| MAS | 90 | 108.09 d | 91.18 | 125.97 | 2.02 ± 0.21 | 4.04 | 0.26 | 140.37 |
| MOS | 90 | 172.59 e | 135.17 | 229.43 | 1.19 ± 0.19 | 0.03 | 1.00 | 224.14 |
| Population | N | IC50 (µg/mL) | 95% Confidence Limits | Slope ± SE | χ2 | p | IR | |
|---|---|---|---|---|---|---|---|---|
| Lower | Upper | |||||||
| LAB | 90 | 0.99 a | 0.78 | 1.20 | 1.57 ± 0.20 | 2.01 | 0.57 | - |
| RAM | 90 | 71.66 b | 56.49 | 85.96 | 1.90 ± 0.22 | 3.60 | 0.31 | 72.38 |
| AZM | 90 | 79.04 b | 58.26 | 98.73 | 1.43 ± 0.20 | 0.59 | 0.90 | 79.83 |
| BAM | 90 | 84.53 b | 60.85 | 107.25 | 1.29 ± 0.20 | 1.81 | 0.62 | 85.38 |
| SAS | 90 | 100.49 b | 78.87 | 122.72 | 1.50 ± 0.20 | 4.11 | 0.25 | 101.50 |
| MOS | 90 | 104.97 bc | 57.58 | 156.29 | 0.72 ± 0.18 | 0.03 | 1.00 | 106.03 |
| NOS | 90 | 173.28 cd | 127.13 | 253.73 | 0.94 ± 0.19 | 0.89 | 0.83 | 175.03 |
| WES | 90 | 214.28 d | 172.88 | 281.99 | 1.40 ± 0.20 | 1.24 | 0.74 | 216.44 |
| MAS | 90 | 307.76 d | 246.25 | 422.88 | 1.58 ± 0.21 | 0.19 | 0.98 | 310.86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abobakr, Y.; Al-Hussein, F.I.; Bayoumi, A.E.; Alzabib, A.A.; Al-Sarar, A.S. Organophosphate Insecticides Resistance in Field Populations of House Flies, Musca domestica L.: Levels of Resistance and Acetylcholinesterase Activity. Insects 2022, 13, 192. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020192
Abobakr Y, Al-Hussein FI, Bayoumi AE, Alzabib AA, Al-Sarar AS. Organophosphate Insecticides Resistance in Field Populations of House Flies, Musca domestica L.: Levels of Resistance and Acetylcholinesterase Activity. Insects. 2022; 13(2):192. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020192
Chicago/Turabian StyleAbobakr, Yasser, Faisal I. Al-Hussein, Alaa E. Bayoumi, Ali A. Alzabib, and Ali S. Al-Sarar. 2022. "Organophosphate Insecticides Resistance in Field Populations of House Flies, Musca domestica L.: Levels of Resistance and Acetylcholinesterase Activity" Insects 13, no. 2: 192. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020192