
Phylogenetic Position of a New Trisetacus Mite Species (Nalepellidae) Destroying Seeds of North American Junipers and New Hypotheses on Basal Divergence of Eriophyoidea

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
3.1. Morphological Description of Trisetacus indelis n. sp.
3.2. GenBank Data and Cox1 Sequence Diversity
3.3. Molecular Phylogenetic Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bolton, S.J.; Chetverikov, P.E.; Klompen, H. Morphological support for a clade comprising two vermiform mite lineages: Eriophyoidea (Acariformes) and Nematalycidae (Acariformes). Syst. Appl. Acarol. 2017, 22, 1096–1131. [Google Scholar] [CrossRef]
- Bolton, S.J.; Bauchan, G.R.; Chetverikov, P.E.; Ochoa, R.; Klompen, H. A rudimentary sheath for the smallest of “biting” chelicerae: The mouthparts of Cunliffea (Nematalycidae) and a new hypothesis on the origin of the stylet sheath of Eriophyoidea (Acariformes). Int. J. Acarol. 2018, 44, 374–381. [Google Scholar] [CrossRef]
- Klimov, P.B.; OConnor, B.M.; Chetverikov, P.E.; Bolton, S.J.; Pepato, A.R.; Mortazavi, A.L.; Tolstikov, A.V.; Bauchan, G.R.; Ochoa, R. Comprehensive phylogeny of acariform mites (Acariformes) provides insights on the origin of the four-legged mites (Eriophyoidea), a long branch. Mol. Phylogenet. Evol. 2018, 119, 105–117. [Google Scholar] [CrossRef]
- Lindquist, E.E. External anatomy and notation of structures. In Eriophyoid Mites: Their Biology, Natural Enemies and Control. World Crop Pests 6; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 3–31. [Google Scholar]
- Nuzzaci, G.; Alberti, G. Internal anatomy and physiology. In Eriophyoid Mites: Their Biology, Natural Enemies and Control. World Crop Pests 6; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 101–150. [Google Scholar] [CrossRef]
- Craemer, C. A Systematic Appraisal of The Eriophyoidea (Acari: Prostigmata). Ph.D. Dissertation, University of Pretoria, Pretoria, South Africa, November 2010. Available online: https://repository.up.ac.za/handle/2263/28832?show=full (accessed on 10 February 2022).
- Li, H.S.; Xue, X.F.; Hong, X.Y. Homoplastic evolution and host association of Eriophyoidea (Acari, Prostigmata) conflict with the morphological-based taxonomic system. Mol. Phylogenet. Evol. 2014, 78, 185–198. [Google Scholar] [CrossRef] [PubMed]
- De Lillo, E.; Craemer, C.; Amrine, J.W.; Nuzzaci, G. Recommended procedures and techniques for morphological studies of Eriophyoidea (Acari: Prostigmata). In Eriophyoid Mites: Progress and Prognoses; Euckermann, E.A., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 283–307. [Google Scholar] [CrossRef]
- Fenton, B.; Jones, A.T.; Malloch, G.; Thomas, W.P. Molecular ecology of some Cecidophyopsis mites (Acari: Eriophyidae) on Ribes species and evidence for their natural cross colonisation of blackcurrant (R. nigrum). Ann. Appl. Biol. 1996, 128, 405–414. [Google Scholar] [CrossRef]
- Navia, D.; de Moraes, G.J.; Roderick, G.; Navajas, M. The invasive coconut mite Aceria guerreronis (Acari: Eriophyidae): Origin and invasion sources inferred from mitochondrial (16S) and nuclear (ITS) sequences. Bull. Entomol. Res. 2005, 95, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Skoracka, A.; Dabert, M. The cereal rust mite Abacarus hystrix (Acari: Eriophyoidea) is a complex of species: Evidence from mitochondrial and nuclear DNA sequences. Bull. Entomol. Res. 2010, 100, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.E.; de Mendonça, R.S.; Skoracka, A.; Silva, E.S.; Navia, D. Integrative taxonomy of Abacarus mites (Eriophyidae) associated with hybrid sugarcane plants, including description of a new species. Exp. Appl. Acarol. 2019, 78, 373–401. [Google Scholar] [CrossRef] [PubMed]
- Chetverikov, P.E.; Fedorov, D.S.; Letukhova, V.Y.; Romanovich, A.E. Description of Cecidophyes fibigiae n. sp., new combinations, records, and DNA barcodes of eriophyid mites (Eriophyoidea, Eriophyidae) from Karadag Nature Reserve (Crimea). Syst. Appl. Acarol. 2021, 26, 818–828. [Google Scholar] [CrossRef]
- Amrine, J.W., Jr.; Stasny, T.A.H.; Flechtmann, C.H.W. Revised Keys to The World Genera of The Eriophyoidea (Acari: Prostigmata); Indira Publishing House: Michigan, MI, USA, 2003; p. 244. [Google Scholar]
- Xue, X.F.; Dong, Y.; Deng, W.; Hong, X.Y.; Shao, R. The phylogenetic position of eriophyoid mites (superfamily Eriophyoidea) in Acariformes inferred from the sequences of mitochondrial genomes and nuclear small subunit (18S) rRNA gene. Mol. Phylogenet. Evol. 2017, 109, 271–282. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Cvrković, T.; Makunin, A.; Sukhareva, S.; Vidović, B.; Petanović, R. Basal divergence of Eriophyoidea (Acariformes, Eupodina) inferred from combined partial COI and 28S gene sequences and CLSM genital anatomy. Exp. Appl. Acarol. 2015, 67, 219–245. [Google Scholar] [CrossRef]
- Boczek, J.; Shevchenko, V.G.; Davis, R. Generic Key to World Fauna of Eriophyoid Mites (Acarida: Eriophyoidea); Warsaw Agricultural University Press: Warsaw, Poland, 1989; p. 192. [Google Scholar]
- Shevchenko, V.G.; Bagnyuk, I.G.; Sukhareva, S.I. A new family of Pentasetacidae (Acariformes, Tetrapodili) and its role in treatment of the origin and evolution of the group. Zool. Zh. 1991, 70, 47–53. [Google Scholar]
- Bagnjuk, I.G.; Sukhareva, S.I.; Shevchenko, V.G. Major trends in the evolution of four-legged mites as a specialized group (using families Pentasetacidae Shev., Nalepellidae Roiv. and Phytoptidae Murray (Acari: Tetrapodili) as examples). Acarina 1998, 6, 59–76. [Google Scholar]
- Li, H.S.; Hoffmann, A.A.; Guo, J.F.; Zuo, Y.; Xue, X.F.; Pang, H.; Hong, X.Y. Identification of two lineages of host-associated eriophyoid mites predisposed to different levels of host diversification. Mol. Phylogenet. Evol. 2016, 105, 235–240. [Google Scholar] [CrossRef]
- Castagnoli, M.; Lewandowski, M.; Łabanowski, G.S.; Simoni, S.; Soika, G.M. An insight into some relevant aspects concerning eriophyoid mites inhabiting forests, ornamental trees and shrubs. Exp. Appl. Acarol. 2010, 51, 169–189. [Google Scholar] [CrossRef]
- Lewandowski, M.; Kozak, M. Distribution of eriophyoid mites (Acari: Eriophyoidea) on coniferous trees. Exp. Appl. Acarol. 2008, 44, 89–99. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Cvrković, T.; Eimov, P.G.; Klimov, P.B.; Petanović, R.U.; Romanovich, A.E.; Schubert, M.A.; Sukhareva, S.I.; Zukof, S.N.; Amrine, J. Molecular phylogenetic analyses reveal a deep dichotomy in the conifer-inhabiting genus Trisetacus (Eriophyoidea: Nalepellidae), with the two lineages differing in their female genital morphology and host associations. Exp. Appl. Acarol. 2020, 81, 287–316. [Google Scholar] [CrossRef]
- Boczek, J.; Shevchenko, V.G. Ancient associations: Eriophyoid mites on gymnosperms. In Eriophyoid Mites: Their Biology, Natural Enemies and Control. World Crop Pests 6; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 217–225. [Google Scholar] [CrossRef]
- Postner, M. Über eine in den Nadeln der Weißtanne (Abies alba Mill.) lebende Gallmilbe, Trisetacus abietis n. sp. (Eriophyidae, Acarina). Z. Für Angew. Entomol. 1968, 62, 106–117. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Bolton, S.J.; Burlakovskiy, M.S.; Craemer, C.; Efimov, P.G.; Klimov, P.; Neser, S.; Paponova, S.S.; Romanovich, A.; Sukhareva, S.I.; et al. Supplementary descriptions and DNA barcodes of two rarely encountered Trisetacus species (Eriophyoidea, Phytoptidae) associated with Tertiary relict conifers from the Mediterranean region. Syst. Appl. Acarol. 2019, 24, 1631–1652. [Google Scholar] [CrossRef]
- Bouneb, M.; de Lillo, E.; Roversi, P.F.; Simoni, S. Molecular detection assay of the bud mite Trisetacus juniperinus on Cupressus sempervirens in nurseries of central Italy. Exp. Appl. Acarol. 2014, 62, 161–170. [Google Scholar] [CrossRef]
- Shevchenko, V.G.; De-Millo, A.P. Life-cycle of Trisetacus kirghisorum (Acarina: Tetrapodili)—Pest of Juniper semiglobosa Rgl. Vestn. Leningr. Univ. 1968, 3, 60–67. [Google Scholar]
- De-Millo, A.P. O dimorfizme samtsov u chetyrekhnogikh kleshchei (Acarina, Eriophyidae). Vestn. Leningr. Univ. 1967, 3, 26–33. [Google Scholar]
- Smith, I.M. Review of species of Trisetacus (Acari: Eriophyoidea) from North America, with comments on all nominate taxa in the genus. Can. Entomol. 1984, 116, 1157–1211. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Amrine, J.W., Jr.; Manson, D.C.M. Preparation, mounting and descriptive study of eriophyoid mites. In Eriophyoid Mites: Their Biology, Natural Enemies and Control. World Crop Pests 6; Lindquist, E.E., Sabelis, M.W., Bruin, J., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1996; Volume 6, pp. 383–396. [Google Scholar] [CrossRef]
- Chetverikov, P.E. Comparative confocal microscopy of internal genitalia of phytoptine mites (Eriophyoidea, Phytoptidae): New generic diagnoses reflecting host-plant associations. Exp. Appl. Acarol. 2014, 62, 129–160. [Google Scholar] [CrossRef]
- World Flora Online. Available online: http://www.worldfloraonline.org (accessed on 1 December 2021).
- Chetverikov, P.E. Video projector: A digital replacement for camera lucida for drawing mites and other microscopic objects. Syst. Appl. Acarol. 2016, 21, 1278–1280. [Google Scholar] [CrossRef]
- Chetverikov, P.E.; Craemer, C.; Cvrković, T.; Klimov, P.B.; Petanović, R.U.; Romanovich, A.E.; Sukhareva, S.I.; Zukoff, S.N.; Bolton, S.; Amrine, J. Molecular phylogeny of the phytoparasitic mite family Phytoptidae (Acariformes: Eriophyoidea) identified the female genitalic anatomy as a major macroevolutionary factor and revealed multiple origins of gall induction. Exp. Appl. Acarol. 2021, 83, 31–68. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transformation. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2017, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Douaihy, B.; Chetverikov, P.; Machon, N.; Dagher-Kharrat, M. Eriophyoid Mite of the Genus Trisetacus Reported on Juniperus excelsa in Lebanon. Am. J. Plant Sci. 2013, 4, 395–399. [Google Scholar] [CrossRef] [Green Version]
- Cvrković, T.; Chetverikov, P.; Vidović, B.; Petanović, R. Cryptic speciation within Phytoptus avellanae s.l. (Eriophyoidea: Phytoptidae) revealed by molecular data and observations on molting Tegonotus-like nymphs. Exp. Appl. Acarol. 2016, 68, 83–96. [Google Scholar] [CrossRef]
- Keifer, H.H. Eriophyid Studies XII. Bull. Calif. Dep. Agric. 1942, 31, 117–129. [Google Scholar]
- Guo, J.F.; Li, H.S.; Wang, B.; Xue, X.F.; Hong, X.Y. DNA barcoding reveals the protogyne and deutogyne of Tegolophus celtis sp. nov. (Acari: Eriophyidae). Exp. Appl. Acarol. 2015, 67, 393–410. [Google Scholar] [CrossRef]
- Shevchenko, V.G. Novii chetirekhnogij Kleshch Trisetacus kirghizorum sp. n. Acarina, Eriophyidae) vreditel semjan arci. Tr. Kirghyzskoy Lesn. Opytn. Stantzii 1962, 3, 299–305. [Google Scholar]
- Bagnjuk, I.G. The eriophyid mite, Trisetacus piceae (Roiv.) (Acarina, Eriophyoidea)—Pest of Norway spruce. Tr. Petergof. Biol. Inst. 1976, 24, 130–145. [Google Scholar]
- Skoracka, A.; Smith, L.; Oldfield, G.; Cristofaro, M.; Amrine, J.W. Host-plant specificity and specialization in eriophyoid mites and their importance for the use of eriophyoid mites as biocontrol agents of weeds. In Eriophyoid Mites: Progress and Prognoses; Euckermann, E.A., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 93–113. [Google Scholar] [CrossRef]
- Farjon, A.; Filer, D. An Atlas of The World’s Conifers: An Analysis of Their Distribution, Biogeography, Diversity and Conservation Status; Brill: Leiden, BO, USA, 2013; p. 512. [Google Scholar] [CrossRef]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Lewandowski, M.; Skoracka, A.; Szydło, W.; Kozak, M.; Druciarek, T.; Griffiths, D.A. Genetic and morphological diversity of Trisetacus species (Eriophyoidea: Phytoptidae) associated with coniferous trees in Poland: Phylogeny, barcoding, host and habitat specialization. Exp. Appl. Acarol. 2014, 63, 497–520. [Google Scholar] [CrossRef] [Green Version]
- Skoracka, A.; Magalhaes, S.; Rector, B.G.; Kuczyński, L. Cryptic speciation in the Acari: A function of species lifestyles or our ability to separate species? Exp. Appl. Acarol. 2015, 67, 165–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skoracka, A.; Lopes, L.F.; Alves, M.J.; Miller, A.; Lewandowski, M.; Szydło, W.; Majer, A.; Różańska, E.; Kuczyński, L. Genetics of lineage diversification and the evolution of host usage in the economically important wheat curl mite, Aceria tosichella Keifer, 1969. BMC Evol. Biol. 2018, 18, 122. [Google Scholar] [CrossRef] [PubMed]
- Chetverikov, P.E.; Klimov, P.; Letukhova, V.Y.; Ripka, G.; Zukoff, S. Two new phyllocoptine species and new records of eriophyoid mites (Eriophyidae, Phyllocoptinae) from Crimea, with evaluation of the phylogenetic position of the new taxa using GenBank data. Syst. Appl. Acarol. 2019, 24, 882–904. [Google Scholar] [CrossRef]
- Liu, Q.; Yuan, Y.M.; Lai, Y.; Wang, G.Q.; Xue, X.F. Unravelling the phylogeny, cryptic diversity and morphological evolution of Diptilomiopus mites (Acari: Eriophyoidea). Exp. Appl. Acarol. 2019, 79, 323–344. [Google Scholar] [CrossRef]
- Miller, R.F.; Chambers, J.C.; Evers, L.; Williams, C.J.; Snyder, K.A.; Roundy, B.A.; Pierson, F.B. The Ecology, History, Ecohydrology, and Management of Pinyon and Juniper Woodlands in the Great Basin and Northern Colorado Plateau of the Western United States; Gen. Tech. Rep. RMRS-GTR-403; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2019; p. 284.
- Hong, X.; Zhang, Z.Q. A cladistic analysis of the Eriophyoidea (Acari: Prostigmata): Tests of monophyly of families. Syst. Appl. Acarol. 1996, 1, 107–122. [Google Scholar] [CrossRef]
- Klimov, P.B.; (Michigan State University, Ann Arbor, MI, USA). Personal communication, 2021.
- Sukhareva, S.I. Family Phytoptidae Murray 1877 (Acari: Tetrapodili), its consisting, structure and suggested ways of evolution. Acarina 1994, 2, 47–72. [Google Scholar]
- Bagnjuk, I.G.; Sukhareva, S.I.; Shevchenko, V.G. Analysis of the genus Trisetacus Keifer as a key of the family Nalepellidae Roiv. (Acari, Tetrapodili). Acarina 1995, 3, 31–65. [Google Scholar]








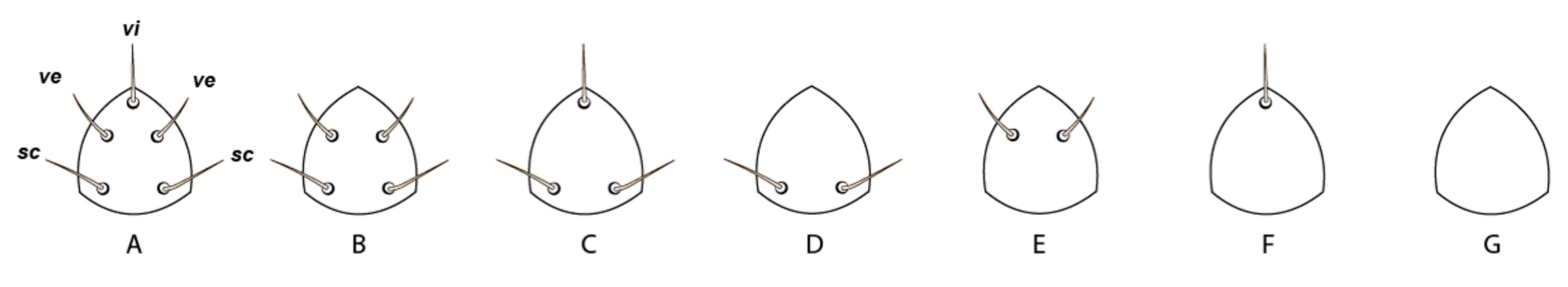{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
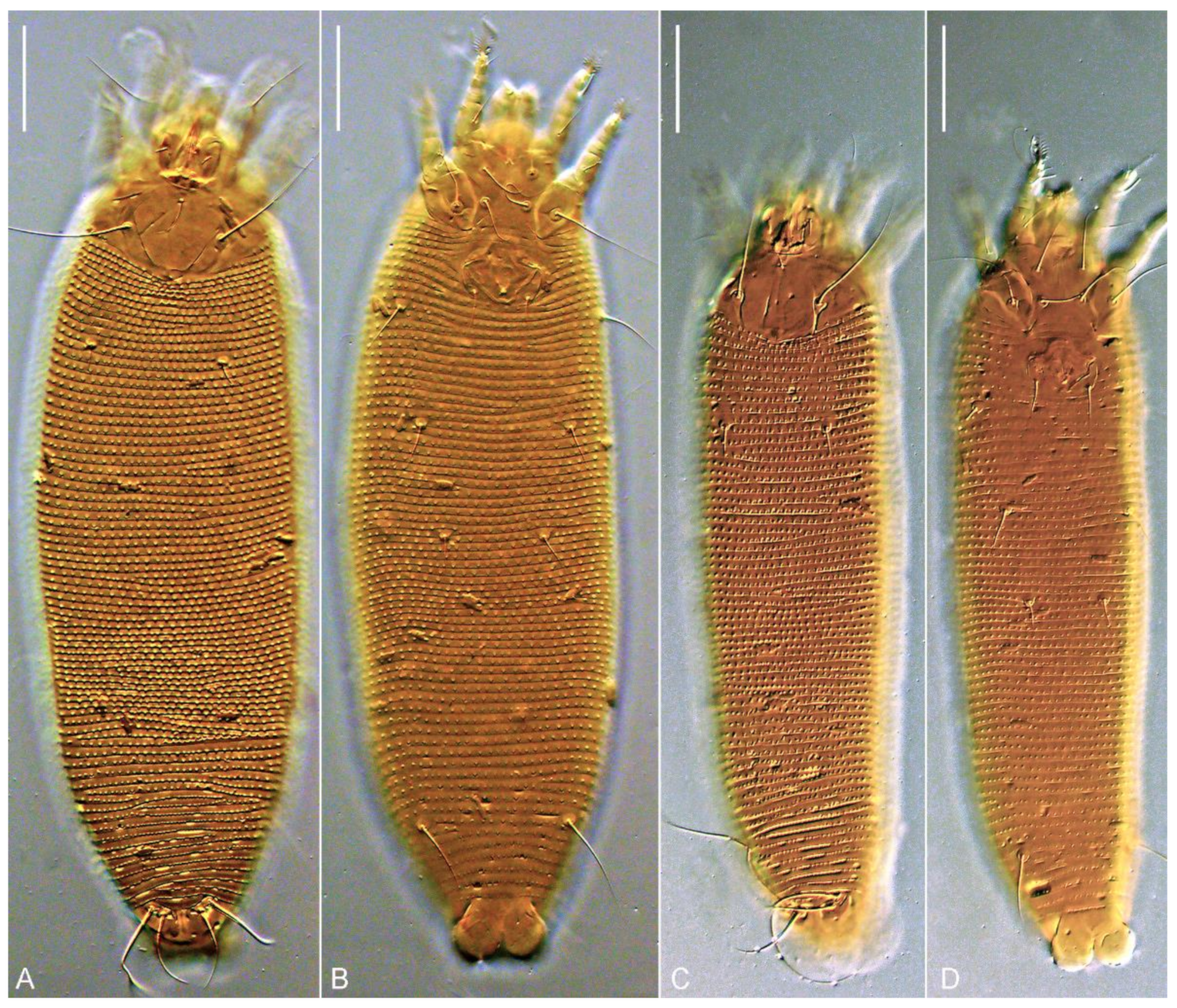{kind=link}
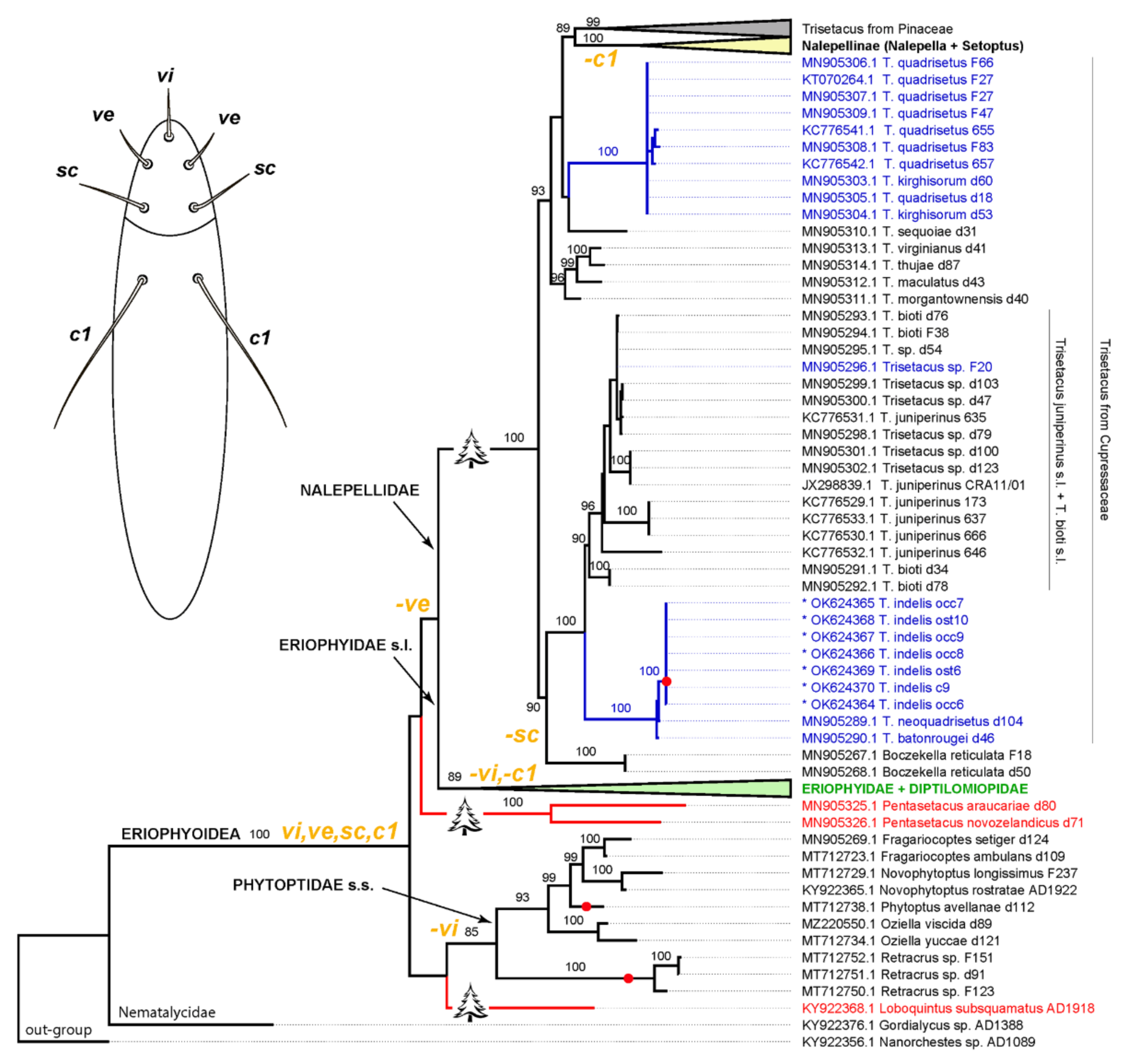{kind=link}
| GB Accession Number | Isolate | Length | Female Form | Host Species |
|---|---|---|---|---|
| OK624364 | occ6 | 696 bp | deutogyne | Juniperus occidentalis |
| OK624365 | occ7 | 1152 bp | protogyne | Juniperus occidentalis |
| OK624366 | occ8 | 1152 bp | deutogyne | Juniperus occidentalis |
| OK624367 | occ9 | 1152 bp | protogyne | Juniperus occidentalis |
| OK624368 | ost10 | 1152 bp | deutogyne | Juniperus osteosperma |
| OK624369 | ost6 | 1103 bp | protogyne | Juniperus osteosperma |
| OK624370 | c9 | 632 bp | protogyne | Juniperus californica |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chetverikov, P.E.; Rector, B.G.; Tonkel, K.; Dimitri, L.; Cheglakov, D.S.; Romanovich, A.E.; Amrine, J. Phylogenetic Position of a New Trisetacus Mite Species (Nalepellidae) Destroying Seeds of North American Junipers and New Hypotheses on Basal Divergence of Eriophyoidea. Insects 2022, 13, 201. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020201
Chetverikov PE, Rector BG, Tonkel K, Dimitri L, Cheglakov DS, Romanovich AE, Amrine J. Phylogenetic Position of a New Trisetacus Mite Species (Nalepellidae) Destroying Seeds of North American Junipers and New Hypotheses on Basal Divergence of Eriophyoidea. Insects. 2022; 13(2):201. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020201
Chicago/Turabian StyleChetverikov, Philipp E., Brian G. Rector, Kirk Tonkel, Lindsay Dimitri, Denis S. Cheglakov, Anna E. Romanovich, and James Amrine. 2022. "Phylogenetic Position of a New Trisetacus Mite Species (Nalepellidae) Destroying Seeds of North American Junipers and New Hypotheses on Basal Divergence of Eriophyoidea" Insects 13, no. 2: 201. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020201