
The Effect of Temperatures and Hosts on the Life Cycle of Spodoptera frugiperda (Lepidoptera: Noctuidae)


1
State Key Laboratory Breeding Base of Green Pesticide and Agricultural Bioengineering, Key Laboratory of Green Pesticide and Agricultural Bioengineering, Ministry of Education, Guizhou University, Guiyang 550025, China
2
Guizhou Provincial Key Laboratory for Agricultural Pest Management of the Mountainous Region, Scientific Observing and Experimental Station of Crop Pest in Guiyang, Ministry of Agriculture, Institute of Entomology, Guizhou University, Guiyang 550025, China
3
Crop Protection Center of Jinsha County, Bijie 551700, China
4
College of Tobacco Science, Guizhou University, Guiyang 550025, China
*
Author to whom correspondence should be addressed.
Insects 2022, 13(2), 211; https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020211
Submission received: 4 January 2022
/
Revised: 10 February 2022
/
Accepted: 16 February 2022
/
Published: 20 February 2022
Abstract
:Simple Summary
Fall armyworm (FAW), Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae) is an invasive worldwide agricultural pest that seriously threatens the safety of grain production. Temperatures and hosts play vital roles in the performance of S. frugiperda. In this study, we studied the effects of three different temperatures (20, 25, and 30 °C) and three different host plants (maize (Zea mays L. (Poaceae)), sorghum (Sorghum bicolor (L.) Moench), and coix seed (Coix lacryma-jobi L.)) on the whole life cycle of S. frugiperda. The results showed that both temperatures and host plants significantly influenced the mortality, developmental time, reproduction, and population parameters of FAW. In the treatment with host plants, the developmental period of S. frugiperda at each stage shortened significantly with the increasing of temperatures. At each temperature, the type of host plant did not affect the development time of S. frugiperda eggs. However, there was no significant difference when S. frugiperda 1st to 6th instar larvae were fed different host plants at 20 °C, while 2nd to 5th instar larvae developed rapidly on maize at 25 and 30 °C. Moreover, there were no significant differences in the longevity of FAW when fed sorghum and coix seed at 20 and 25 °C, but a significant difference in the longevity was observed at 30 °C. Feeding on maize achieved a higher intrinsic rate of increase (r), finite rate of increase (λ), and net reproductive rate (R0) of S. frugiperda at 25 and 30 °C than those FAW fed on sorghum and coix seed.
Abstract
The interactions between ambient temperatures and host plants are central to the population dynamics of invasive animal species. Despite significant research into the effects of temperatures, the performance of invasive species is also influenced by host plants. The effects of different temperatures (20, 25, and 30 °C) and host plants (maize, sorghum, and coix seed) were tested on the mortality, development, reproduction, and population parameters of the fall armyworm (FAW), Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), using an age-stage, two-sex life table. The results support the hypothesis that temperature and the species of the host plant significantly influences the performance of FAW. Feeding on maize at 30 °C resulted in a lower mortality rate, a shorter developmental time and longevity, a higher fecundity, intrinsic rate of natural increase (r), finite rate of increase (λ), and net reproductive rate (R0). However, at 20 °C, the host plant could eliminate temperature-mediated synergism in FAW performance, which did not reach statistical significance at 20 °C. Similar results induced by a relatively low temperature (20 °C) on different host plants were also found in the age-stage specific survival curves (sxj), fecundity (mx), maternity (lxmx), and reproductive value (vxj) curves of FAW. Consequently, we also need to pay more attention to FAW outbreaks on different host plants mediated by relatively low temperatures.
1. Introduction
Global climate change may substantially affect the diverse geographical distributions of organisms, alter biodiversity, and influence the interactions between species and ecosystems [1]. Invasive species can be more capable than native species at responding to climate change due to their high fecundity and abundance [2,3]. Temperature-driven changes are thought to alter ecosystem stability and sustain generational cycles for multivoltine insects as temperatures increase [4]. Therefore, increasing temperatures may expand the overall global distribution of invasive crop pest species, triggering outbreaks, and, in turn, increasing the damage to crops [4,5,6]. The establishment and spread of invasive pest species in new environments are commonly attributed to a lack of natural enemies. However, the lack of shared evolutionary host plants in invasive regions also plays an important role in facilitating pest invasions [7]. We tested the hypothesis that rising temperature leads to the enhancement of fitness of invasive species under the same host feeding condition for invasive species.
The fall armyworm (FAW), Spodoptera frugiperda (J E Smith) (Lepidoptera: Noctuidae) is one of the most damaging crop pests in the Americas. Its larva can attack more than 353 host plants from 76 families, principally Poaceae (106 taxa) [8]. FAW has long-distance flight capability and is confirmed as an inhabitant in continental Africa (47 countries), Asia (18 countries), and mainland Australia as of February 2020 [9,10]. Due to the absence of diapause and a short generation time, FAW populations can occur all year round in non-native countries with suitable temperatures and host plants [11]. We should explore the fitness of FAW under abiotic (e.g., temperature) and biotic (e.g., host plant) stressors in invasion regions.
FAW is a highly polyphagous herbivore represented by two sympatric host plant strains: a rice strain that feeds on rice and various grasses and a corn strain that primarily feeds on maize, sorghum, cotton, and sugarcane [12,13]. Strain identification analyses based on COI and Tpi genes showed that a sample of FAW in Yunnan province in China collected in January 2019 belonged to a corn strain [14]. Based on analysis of the COI gene, a sample from Chongqing City belonged to the rice strain, while both the rice and corn strains were found in June 2019 when categorized by the Tpi gene [15]. This invasive FAW, possibly originating from the offspring of hybrid populations, may pose a risk to different host plants [16]. Understanding the utilization of hybrid populations of FAW to a variety of host plants is necessary to predict the FAW’s annual reproduction and migration [17]. FAW feeds on plants of the Poaceae family, including sorghum, maize, and coix seed [8]. Sorghum is an important dryland food, fodder, and energy crop in China, and the occurrence of FAW on sorghum was observed in June 2019 [18]. Coix seed is a high-quality cereal used both as a food and for medicine in China, where it accounts for more than 70% of the global market. In 2019, FAW was found to damage coix seed in Qichun county, Hubei province, and Xingyi City, Guizhou province in China [19].
The ability of invasive species to regulate their performance is crucial for adapting to new invading environments as the Earth’s climate rapidly changes [20]. Temperature is a central regulating abiotic variable controlling the development, fecundity, and range of invasive species [20]. The developmental rates of invasive insects increase in response to increasing temperatures [21]. Although FAW is not able to overwinter in northern China and North America due to a lack of diapause and suitable host plants, high temperatures trigger FAW outbreaks through an acceleration of their development and reduction in the duration of their life cycle [6,22,23,24]. Therefore, it is essential to evaluate the effects of a temperature gradient on the performance of FAW.
Life history studies of FAW have been undertaken on different host plants (cotton, millet, corn, sorghum, wheat, bermudagrass, soybean, peanut, oilseed rape, sunflower, tomato, pepper, and eggplant) [6,16,17,25,26,27,28,29]. Differences in the suitability of different maize cultivars and varieties of bermudagrass as a host plant for FAW have been recorded [6,25,26]. Climate change alters the responses of FAW to different maize landraces. FAW at high temperatures is more harmful to White Ranchero maize landraces than to Yellow maize landraces [6]. However, the combined role of host plants and temperatures on the entire life history of FAW is poorly understood.
In the current study, we assessed the life history characteristics of FAW on maize (Zea mays L. (Poaceae)), sorghum (Sorghum bicolor (L.) Moench), and coix seed (Coix lacryma-jobi var. ma-yuen (Rom. Caill.) Stapf (CL)) at three constant temperatures (20, 25, and 30 °C). An age-stage, two-sex life table was used to evaluate and compare FAW responses to different host plants and temperatures.
2. Materials and Methods
2.1. Insects and Plants
Spodoptera frugiperda individuals were originally collected from corn fields on the 22 April 2019 in Guiding County (107.24° N, 26.58° E), Guizhou Province, China. The stock colony was reared for three generations at the Institute of Entomology, Guizhou University on fresh young maize leaves within 15–20 cm in laboratory conditions, at 28 ± 2 °C, 70 ± 5% RH, and a photoperiod of 14:10 (L–D) h. Due to the highly cannibalistic behavior of older larvae, 1st to 3rd instar larvae were reared together in a breeding box (60 × 50 × 40 cm), and 4th to 6th instar larvae were reared individually in breeding boxes (100 mL; depth: 4 cm; bottom diameter: 5.5 cm; top diameter: 6.5 cm). The insects pupated in sterilized soil with 15% water. Cotton balls with 10% honey water were provided for the adult insects as supplemental nutrition. To collect the same age of S. frugiperda eggs, 5 pairs of adults were randomly selected from the colonies and placed inside a 1-L cup with Nylon 100 mesh cloth for ventilation and to encourage laying eggs. A cotton ball soaked with a 10% honey solution was placed in the cup to provide food for the adults described by Tian et al. (2020) [30].
The host plants in this study were maize (Jinongnuo No. 7, Beijing Jinnong Technology Co., Ltd., Beijing, China), sorghum (red tassel, Guizhou Hongyingzi Agricultural Technology Development Co., Ltd., Bijie, China) and coix seed (Jinsha Coix). These plants were seeded in a planter tray within potting mix (20 cm depth × 40 cm diameter) and were irrigated twice a week in the greenhouse. Plants with young and healthy leaves (15–20 cm plant height) were used in the experiment.
2.2. Life Table Analysis
To evaluate the effect of host plants and temperatures on the life history parameters of S. frugiperda, 900 eggs produced by S. frugiperda females were maintained in the nylon 100 mesh. Experiments were carried out at 20, 25, and 30 °C on maize, sorghum, and coix seed at 70% ± 5% RH, with a photoperiod of 14:10 (L–D) h within a climate chamber (Jiangnan Instrument Factory, Ningbo, China). One cluster of 100 eggs (less than 24 h old) was prepared on each host plant at each temperature and transferred into a petri dish with wet filter paper (diameter: 9 cm) at the bottom after spraying with sterile distilled water. The petri dish was then placed at a constant temperature, and leaves of a host plant were added. Petri dishes were closed at the top with transparent plastic wrap. An insect pin (No. 5) was used to puncture the plastic wrap 20 times for ventilation. The eggs were checked daily at 8:30 am and 8:30 pm until hatched, and the number of eggs hatched without success or shriveled was considered as death. When FAW eggs hatched (<6 h), they were individually transferred to a plastic box (6 × 5 × 4 cm) with fresh host plant leaves (maize, sorghum, or coix seed) at each temperature. We provided fresh leaves every day for each treatment. When the mature larvae started curling their bodies, indicating they had reached the pre-pupal period, we cleaned away the frass and covered the larvae with a thin layer of soil until eclosion. Newly emerged females and males were paired and placed in the 1-L cups. The plastic cups were furnished with cotton balls with 10% honey water, and the developmental time and mortality rates of immature stages of S. frugiperda were recorded twice every day, at 8:30 a.m. and 8:30 p.m. The duration of pre-oviposition, oviposition, fecundity, and longevity of S. frugiperda was observed.
The preliminary data for all individuals at different temperatures and on different host plants were organized and analyzed according to an age-stage, two-sex life table [31,32]. For the population parameters, mean values and standard errors of developmental time, mortality, pre-oviposition period, total pre-oviposition period, fecundity, and longevity of S. frugiperda were calculated using TWOSEX-MSChart (Chi 2021) [33], estimated with 100,000 bootstrap replicates.
The age-stage survival rate curve (sxj) shows the probability that a newly oviposited egg will survive to age x and stage j. The age-specific survival rate (lx) was approximated as
where m is the number of pest stages. Age-specific fecundity (mx) was represented by the equation
The intrinsic rate of increase (r) was predicted using the equation
The net reproductive rate (R0) was the total offspring that an individual produced during its lifetime and was depicted as
The gross reproductive rate (GRR) was determined as
The finite rate of increase (λ) was calculated as
The mean generation time (T) was defined as the length of time a population requires to increase by R0-fold (i.e., ert = R0 or λT = R0) at a stable age distribution, and was estimated as
The age-stage life expectancy (exj) is the lifespan that an individual of age x and stage j is expected to live after age x. Because individuals of the same age may differ in stage of development, for individuals of the same age, the life expectancy for an individual of age x and stage y is calculated as
where n is the last age of individuals in the population and m is the number of stages. is the probability that an individual of age x and stage y will survive to age i and stage j, and is calculated by assuming = 1 [34]. Reproductive value (vxj) is described as the contribution of an individual of age x and age j to the future population and was determined as
For a proper application of the bootstrap technique, the randomization function was used in TWOSEX-MSChart. Moreover, according to Akca et al. (2015) [35], we used 100,000 resamplings to obtain stable estimates of standard errors. A paired bootstrap test was used to detect statistical differences in the mortality at different stages of S. frugiperda on different host plants or different temperatures [33].
3. Results
3.1. Mortality Rate, Developmental Time, Adult Longevity, and Reproduction
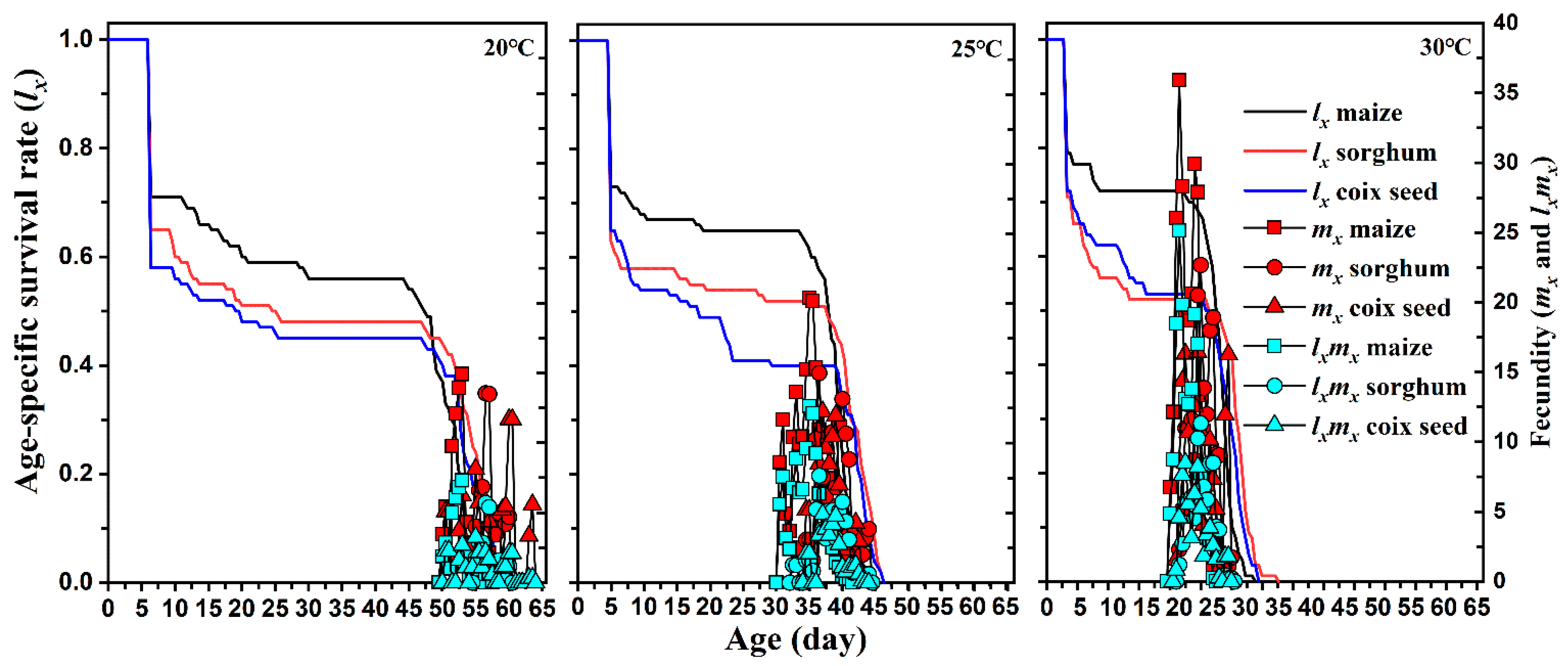
The mortality of S. frugiperda reared on different host plants at different temperatures is shown in Table 1. Host plants and temperatures significantly influenced the mortality rate of S. frugiperda pupae and pre-adults, while no significant differences were found in the mortality of larvae except between temperatures for the 3rd to 5th instar larvae fed sorghum. The mortality rates of S. frugiperda pre-adults were significantly influenced by temperatures when fed maize but not on sorghum and coix seed. Host plants induced significant differences in the mortality rate of S. frugiperda pre-adults between maize and the other two host plants at 30 °C, while no significant differences were found at 20 °C. A similar trend was also shown in the age-stage specific survival curves (sxj): the curves for pupa, female, and male fed on maize were higher than those FAW fed on sorghum and coix seed at 30 °C but not at 20 °C (Figure 1).
The developmental duration of S. frugiperda reared on different host plants under different temperatures is provided in Table 2. The age-specific survival rate (lx), fecundity (mx), and maternity (lxmx) curves of S. frugiperda reared on different host plants at different temperatures are shown in Figure 2. In all host plant conditions, the temperature significantly impacted the developmental time of S. frugiperda eggs, 1st to 6th instar larvae, pre-pupae, pupae, and pre-adults (P < 0.05); when temperature increased, the developmental period of S. frugiperda at each stage shortened significantly. At each temperature, the type of host plant did not affect the development time of S. frugiperda eggs. There was no significant difference when S. frugiperda 1st to 6th instar larvae were fed different host plants at 20 °C, while 2nd to 5th instar larvae developed rapidly on maize at 25 and 30 °C (P < 0.05). However, at 20 and 30 °C, the developmental duration of S. frugiperda 2nd to 5th instar larvae fed maize was shorter than when fed sorghum and coix seed, while there was no significant difference between sorghum and coix seed. The duration of pre-pupae and pupae were significantly influenced by plant host type at all temperatures, except for the pre-pupae at 25 °C. Feeding on maize significantly shortened the duration of the immature stages of S. frugiperda at each temperature.
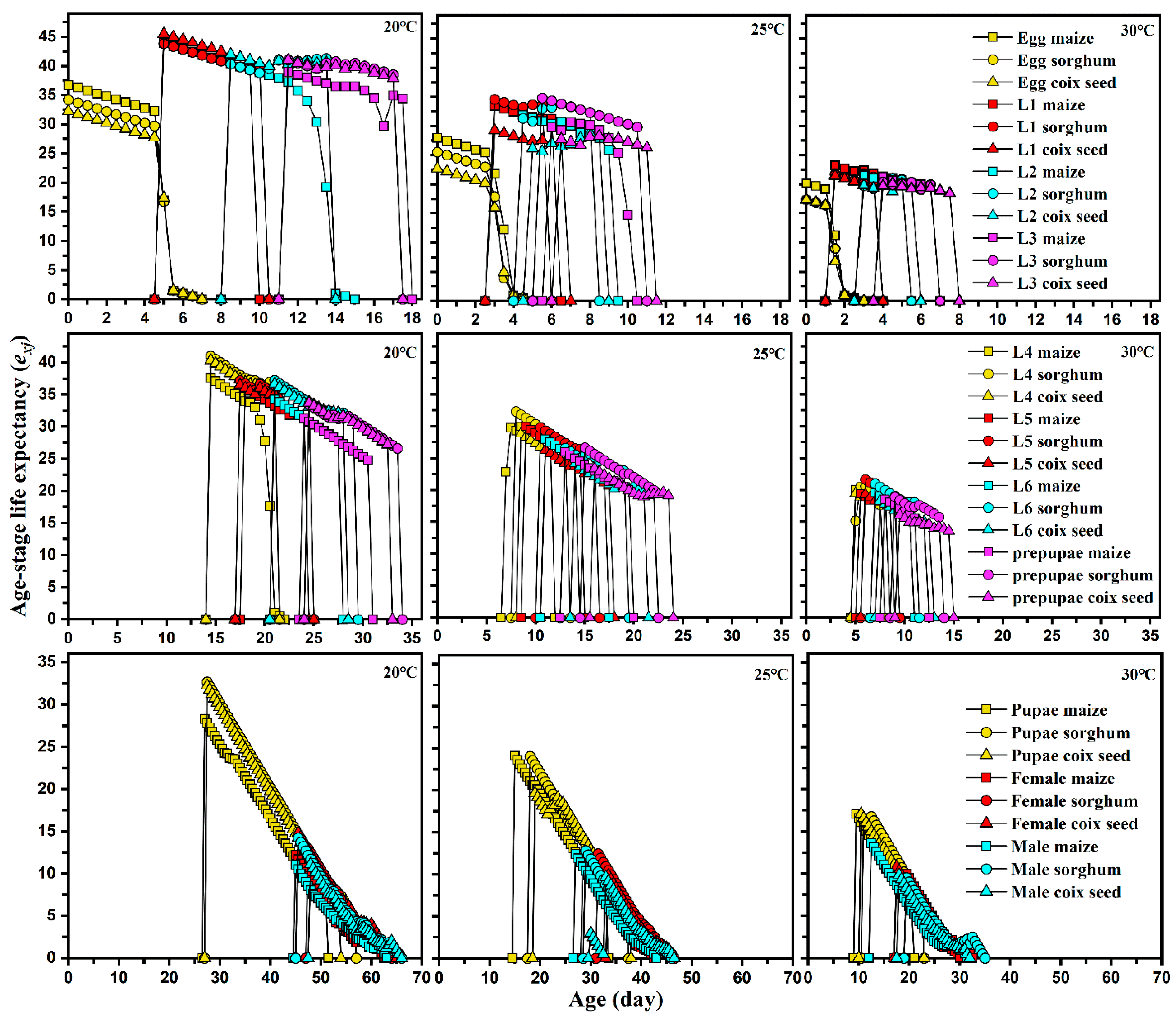
The adult longevity and reproduction of S. frugiperda reared on different host plants at different temperatures are shown in Table 3. Host plants and ambient temperatures significantly influenced the longevity, total pre-oviposition period, and fecundity of FAW. The longevity of FAW females and males was shortest when FAW were fed maize at 30 °C. There were no significant differences in the longevity of FAW when fed sorghum and coix seed at 20 and 25 °C, but a significant difference was observed at 30 °C. Under the same host, the life expectancy (exj) of S. frugiperda was shortened with increased temperature, showing a significant decreasing trend (Figure 3).
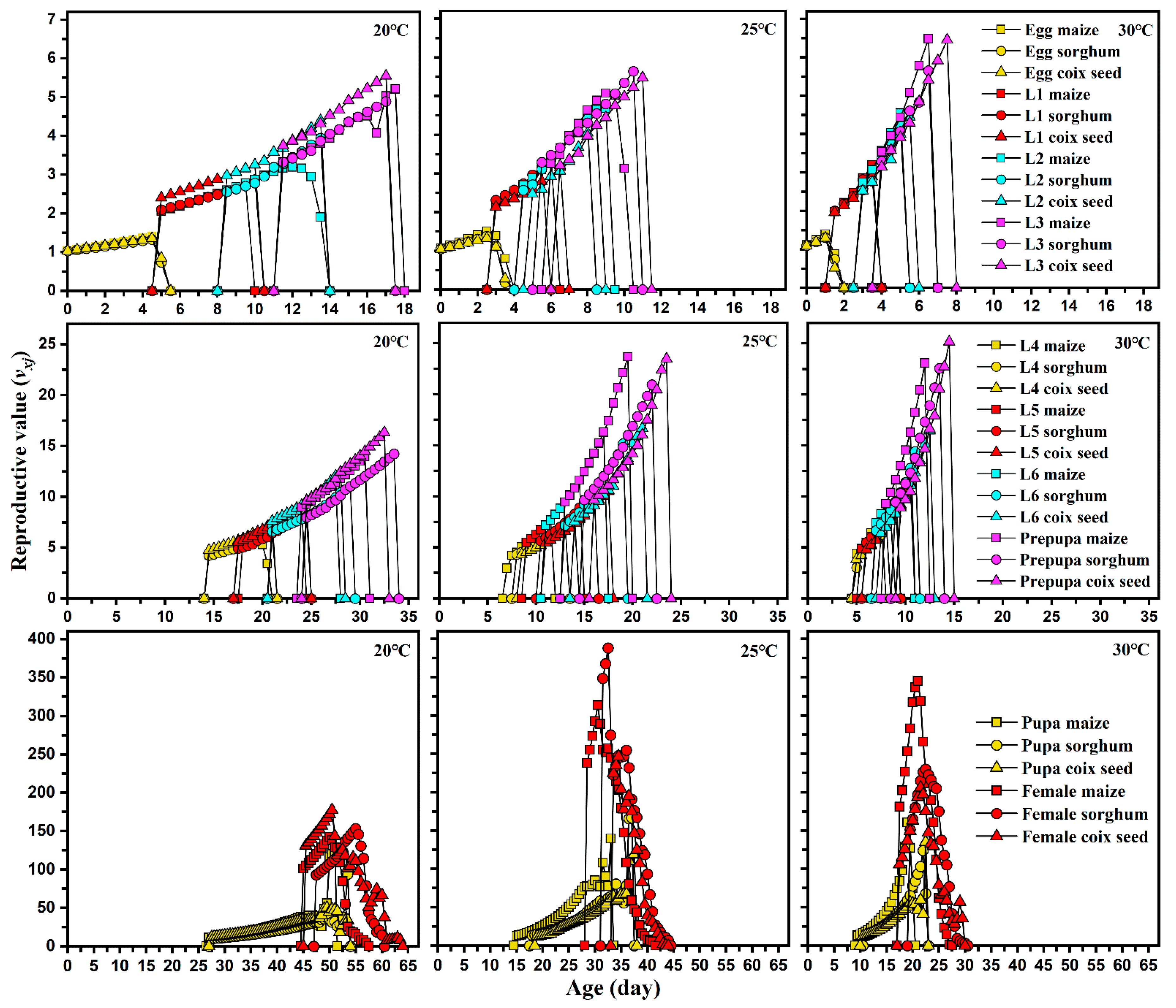
No significant difference was observed in APOP among the three host plants at any temperature. However, the total pre-oviposition period in maize was significantly shorter than that detected under the two other hosts. Female and male adults lived shorter in corn and produced more eggs than those reared on sorghum and coix seed at any temperature. The host plant did not affect the pre-oviposition period (APOP) and oviposition period of FAW at all temperatures. However, a significantly lower total pre-oviposition period (TPOP) was observed on maize host plants. Increasing temperature significantly reduced the APOP and TPOP of FAW, but a shortened oviposition period was observed when FAW was reared on each host plant at 20 °C. Host plants significantly influenced the fecundity of FAW at 25 and 30 °C, but relatively lower temperatures only reduced their fecundity when fed maize. The highest fecundity of FAW was 954.3 and 956.8 when fed maize at 25 and 30 °C, respectively. The highest peaks of mx that occurred on maize (14.9 offspring), sorghum (13.5 offspring), and coix seed (11.6 offspring) were similar at 20°C, but the highest value of mx on maize (35.91 offspring) at 30 °C was higher than that on sorghum and coix seed at 20 °C. Similar results were also found in the maternity (lxmx) curves of S. frugiperda. Moreover, the relatively lower temperature did not induce a significant difference in the highest reproductive values (vxj) of pupa and females among maize, sorghum, and coix seed at 20 °C, but feeding on maize at 30 °C had greater reproductive values of pupa and female than those moths fed on sorghum and coix seed (Figure 4).
3.2. Life Table Parameters
Host plants and temperatures significantly influenced the intrinsic rate of natural increase (r), finite rate of increase (λ), and average generation calendar period (T) of FAW, except at 20 °C for r and λ (Table 4). The values of r, λ, and net reproductive rate (R0) of FAW significantly increased with increasing temperature on each host plant apart from the R0 on coix seed. The highest r, λ, and R0 of FAW were found on maize at 30 °C. Feeding on maize achieved higher r, λ, and R0 at 25 and 30 °C than feeding on sorghum and coix seed. The T was significantly shorter when FAW were fed maize at all temperatures, while the gross reproductive rate (GRR) was significantly higher than sorghum and coix seed.
4. Discussion
Fall armyworm (FAW) is a highly multivoltine polyphagous herbivore that feeds on commercial crops. Higher temperatures reduce its development time, resulting in more generations during the crop growing season [6,26,36,37]. FAW feed on economically important crops, such as rice, sorghum, maize, sugarcane, wheat, millet, cotton, bermudagrass, and different vegetables [6,17,25,26,27,28,29]. Previous studies only considered the independent responses of temperature or host plant on the performance of FAW. There was little information available on the interactive effects of different temperatures and host plants on the entire life cycle of FAW [6,25]. Our study found that both temperature and host plant significantly influenced the survival, development, reproduction, and population growth of FAW. Host plants did not influence the developmental time of larvae, APOP, oviposition period, fecundity, or life table parameters of FAW at relatively low temperature (20 °C). However, significant differences were found at both 25 and 30 °C. Therefore, different responses by FAW were triggered by temperatures to different host plants.
The development rates of S. frugiperda increased with increasing temperatures. Optimal temperature can improve the survival rate, accelerate the development rate of immature stages, result in more generations, and enhance the probability of their establishment [2,36,37]. The survival rate of FAW larvae and pupae increases in response to increasing temperature [36,38]. However, different temperatures do not influence the percentage of emerged FAW adults [6]. Our study found a lower mortality of FAW pre-adults under higher temperatures when fed maize. It seems that temperature triggers different survival responses by invasive species depending on the host plant. FAW fed on tomato and potato have significantly lower survival rates at 25 °C compared to maize host plants [39]. In contrast, FAW larva–adult survival does not differ between maize and sorghum [17], cotton, millet, corn, and soybean [27], and between soybean, cotton, maize, wheat, oat leaves, and an artificial diet [40]. Moreover, no differences were found in larva–adult survival rates when fed on corn, sorghum, wheat, soybean, peanut, and cotton [28]. In our study, no significant difference was found in the mortality of FAW pre-adults across the three host plants at 20 °C. However, 30 °C caused a significantly lower mortality rate of FAW on maize than on sorghum and coix seed, which is consistent with du Plessi’s results [36]. The differential response of host plants to increasing temperature may cause different mortality rates of FAW. Therefore, it was critical to evaluate the performance of invasive species on different host plants at different temperatures.
The developmental time of the immature stages of FAW is significantly influenced by temperature and the type of host plant [6,25,26,36,38,40]. At higher temperatures, the development period of FAW is significantly reduced [26,36,38]. In the present study, we found that increasing temperatures significantly shortened the developmental time of FAW eggs, larvae, pre-pupae, pupae, and pre-adults on each host plant, which significantly accelerated generations. However, temperature caused differences in FAW developmental responses to host plants. There were no significant differences in the developmental time of 2nd to 6th instar larvae at 20 °C, but there was a significant reduction in developmental time on maize host plant at 30 °C. The duration of the larval stage of FAW fed four varieties of bermudagrass at 20 and 30 °C shows the opposite tendency [25] These differences in FAW development based on temperature and host plant type might help predict FAW occurrence in both normal and extreme climatic conditions.
Temperature and the type of host plant significantly influenced the reproduction parameters of FAW [25,26,28,29,38,40]. Increased temperature (30 °C) significantly reduced the longevity of FAW fed on corn, while there was no longevity reduction when the temperature was increased from 21 to 25 °C [26]. Our study found temperature triggered different responses of longevity, TPOP, and fecundity of FAW fed on host plants. No significant differences in longevity and TPOP were found between sorghum and coix seed at 20 and 25 °C, but at 30 °C, the longevity was significantly reduced in FAW fed on coix seed. Higher temperatures increased FAW fecundity when fed on maize. An ambient temperature of 30 °C significantly increased the fecundity of FAW across the three host plants, but there was no significant difference at 20 °C.
Assessment of population parameters is an essential method for evaluating the effect of temperatures and host plants on the fitness of invasive insects [41]. No previous studies have recorded the life table parameters of FAW under different temperatures [6,38]. Our study first reported that increasing temperature significantly affected the population parameters (R0, r, λ, and GRR) of FAW on each host plant, except for the R0 when fed coix seed and GRR when fed sorghum host plants. Host plants significantly influenced the population parameters for FAW [16,27,29,42]. The values of r and λ for FAW fed on maize are significantly higher than those for wheat, soybean, Chinese cabbage, tomato, and pepper [29,42]. FAW fed on millet have higher R0 and r than on cotton and soybean plants [27]. Our study found similar results that feeding on maize caused higher r, λ, and R0 for FAW at 25 and 30 °C than feeding on sorghum and coix seed, but at 20 °C, this trend did not reach statistical significance. Therefore, temperature also influenced the population parameter responses of FAW to the host plants.
Climate anomalies are important drivers for expanding habitat distribution and elevating the invasion risk of invasive species [43]. For example, temperature-driven changes caused insect outbreaks by affecting their life history traits to alter system stability [4]. FAW is a poikilothermic pest species that could regulate growth and development with a temperature change. In our study, the increased temperature caused changes in the life history of FAW in each host plant. However, relatively low temperatures (20 °C) appeared to eliminate temperature-mediated synergism in FAW performance. There were significant life stage effects at 20 °C [44,45]. Feeding on different host species also influenced the cold-tolerance strategy of Epiphyas postvittana (Walker) [46]. Furthermore, FAW collected in China may be the progeny of hybrid populations, which should have developed substantial adaptations to maize plants [16]. Currently, there is little information available on FAW adaptation to coix seeds throughout the world [19]. Adaptation to local host plants may influence the response of the oak specialist butterfly (Erynnis propertius) to climate change [47]. Therefore, strong adaptation to a local host plant (maize) might increase responses to increasing temperature. However, this research still had many limitations. We only determined the performance of FAW on different host plants at 20 °C, 25 °C, and 30 °C under the laboratory condition, and further study should focus on temperatures below 20 °C or above 30 °C. Meanwhile, fluctuations and constant temperatures have different effects on the development and longevity of insects. The insects reared at fluctuating profiles with lower average temperatures develop faster and survive longer than those reared at constant temperatures, while higher average fluctuating temperatures result in longer developmental duration and lower longevity of insects [48,49]. In our study, we only tested the constant temperatures on FAW without considering fluctuating temperatures in the laboratory. We hope that the result stimulates future study on this topic to evaluate the fluctuating temperatures on the occurrence of FAW on different host plants in the field.
5. Conclusions
In conclusion, our experiment carried out detailed observation of each stage after 12 h, and the results may be more accurate than those observed once in 24 h. Meanwhile, our study revealed that temperature and the type of host plant significantly influenced the performance of FAW, and the fitness of FAW on different host plants varied depending on temperature. Lower temperatures could eliminate temperature-mediated synergism of FAW on local host plants and speed up their adaptation to plants in new locales, which may also result in outbreaks of invasive species. In our study, we conducted detached leaf assays to detect the life history of FAW under different temperatures in laboratory conditions. Further studies should focus on the interactions between temperatures and host plant species outside of the lab; we recommend the next step being the greenhouse.
Author Contributions
Conceptualization: Y.-C.C., M.-F.Y. and J.-F.L.; methodology: Y.-C.C.; investigation: Y.-C.C. and D.-F.C.; writing—original draft: Y.-C.C.; data curation: D.-F.C.; supervision: M.-F.Y. and J.-F.L.; writing—reviewing and editing: M.-F.Y. and J.-F.L.; project administration: J.-F.L.; funding acquisition: J.-F.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (32060637), the Systematic & Applied Acarology Society, the Natural Science Special Project of Guizhou University (Special post, [2020]-02), and the Growth Project of Youth Talent in Ordinary Universities in Guizhou Province ([2021]079).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Dataset is available from the first author on reasonable request.
Acknowledgments
We would like to thank Hsin Chi (Zhongxing University, Taiwan) for his help in guiding data analysis and Jia-Yun Zhu (Guizhou University) for the assistance with the experiment.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, J.J.; Byers, J.E.; Bierwagen, B.G.; Dukes, J.S. Five potential consequences of climate change for invasive species. Conserv. Biol. 2008, 22, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.D.; Torchin, M.E.; Hufbauer, R.A.; Lemoine, N.P.; Alba, C.; Blumenthal, D.M.; Bossdorf, O.; Byers, J.E.; Dunn, A.M.; Heckman, R.W.; et al. Do invasive species perform better in their new ranges? Ecology 2013, 94, 985–994. [Google Scholar] [CrossRef] [Green Version]
- Nelson, W.A.; Bjornstad, O.N.; Yamanaka, T. Recurrent insect outbreaks caused by temperature-driven changes in system stability. Science 2013, 341, 796–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.W.; Wang, Y.C.; Feng, C.C.; Wan, P.H.M.; Chang, K.T.T. Potential distributional changes of invasive crop pest species associated with global climate change. Appl. Geogr. 2017, 82, 83–92. [Google Scholar] [CrossRef]
- Díaz-Álvarez, E.A.; Martínez-Zavaleta, J.P.; López-Santiz, E.E.; Barrera, E.D.A.; Larsen, J.; Del-Val, E. Climate change can trigger fall armyworm outbreaks: A developmental response experiment with two Mexican maize landraces. Int. J. Pest Manag. 2021, 1–9. [Google Scholar] [CrossRef]
- Desurmont, G.A.; Donoghue, M.J.; Clement, W.L.; Agrawal, A.A. Evolutionary history predicts plant defense against an invasive pest. Proc. Natl. Acad. Sci. USA 2011, 108, 7070–7074. [Google Scholar] [CrossRef] [Green Version]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host Plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Maino, J.L.; Schouten, R.; Overton, K.; Day, R.; Ekesi, S.; Bett, B.; Barton, M.; Gregg, P.C.; Umina, P.A.; Reynolds, O.L. Regional and seasonal activity predictions for fall armyworm in Australia. Curr. Res. Insect Sci. 2021, 1, 100010. [Google Scholar] [CrossRef]
- Wan, J.; Huang, C.; Li, C.Y.; Zhou, H.X.; Ren, Y.L.; Li, Z.Y.; Xing, L.S.; Zhang, B.; Qiao, X.; Liu, B.; et al. Biology, invasion and management of the agricultural invader: Fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Integr. Agric. 2021, 20, 646–663. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.J.W.; Colmenarez, Y.; Corniani, N.; Early, R.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Pashley, D.P. Host-associated genetic differentiation in fall armyworm (Lepidoptera: Noctuidae): A sibling species complex? Ann. Entomol. Soc. Am. 2013, 79, 898–904. [Google Scholar] [CrossRef]
- Dumas, P.; Legeai, F.; Lemaitre, C.; Scaon, E.; Orsucci, M.; Labadie, K.; Gimenez, S.; Clamens, A.L.; Henri, H.; Vavre, F.; et al. Spodoptera frugiperda (Lepidoptera: Noctuidae) host-plant variants: Two host strains or two distinct species? Genetica 2015, 143, 305–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, D.P.; Guo, J.F.; Jiang, Y.M.; Zhao, J.Z.; Sethi, A.; He, K.L.; Wang, Z.Y. Initial detections and spread of invasive Spodoptera frugiperda in China and comparisons with other noctuid larvae in cornfields using molecular techniques. Insect Sci. 2020, 27, 780–790. [Google Scholar] [CrossRef]
- Tang, Y.L.; Gu, R.C.; Wu, Y.Y.; Zou, X.M.; Zhang, Z.; Niu, X.H.; Wang, Z.L.; Chen, J.; Li, T.; Li, C.F.; et al. Biotype identification of the population of Spodoptera frugiperda that migrated to Chongqing area. J. Southwest Univ. 2019, 41, 1–7. [Google Scholar]
- He, L.M.; Wu, Q.L.; Gao, X.W.; Wu, K.M. Population life tables for the invasive fall armyworm, Spodoptera frugiperda fed on major oil crops planted in China. J. Integr. Agric. 2020, 20, 745–754. [Google Scholar] [CrossRef]
- He, L.M.; Wang, T.L.; Chen, Y.C.; Ge, S.S.; Wyckhuys, K.A.G.; Wu, K.M. Larval diet affects development and reproduction of East Asian strain of the fall armyworm, Spodoptera frugiperda. J. Integr. Agric. 2021, 20, 736–744. [Google Scholar] [CrossRef]
- Lu, Q.S.; Zou, J.Q.; Zhu, K.; Zhang, Z.P. Discussion on the developmental of sorghum industry in China–On the national sorghum production advantage area. Rainfed Crop 2009, 29, 78–80. [Google Scholar]
- Zou, C.H.; Yang, J.J. Spodoptera frugiperda damages coix seed. China Plant Prot. 2019, 39, 47. [Google Scholar]
- Ziska, L.H.; Blumenthal, M.D.; Runion, C.B.; Hunt, E.R., Jr.; Diaz-Soltero, H. Invasive species and climate change: An agronomic perspective. Clim. Chang. 2011, 105, 13–42. [Google Scholar] [CrossRef]
- Forrest, J.R.K. Complex responses of insect phenology to climate change. Curr. Opin. Insect Sci. 2016, 17, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S.J. Migration and the life history strategy of the fall armyworm, Spodoptera frugiperda in the western hemisphere. Int. J. Trop Insect Sci. 1987, 8, 543–549. [Google Scholar] [CrossRef]
- Meagher, R.L. Nagoshi RN, Population dynamics and occurrence of Spodoptera frugiperda host strains in southern Florida. Ecol. Entomol. 2004, 29, 614–620. [Google Scholar] [CrossRef]
- Yang, X.M.; Song, Y.F.; Sun, X.X.; Shen, X.J.; Wu, Q.L.; Zhang, H.W.; Zhang, D.D.; Zhao, S.Y.; Liang, G.M.; Wu, K.M. Population occurrence of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), in the winter season of China. J. Integr. Agric. 2021, 20, 772–782. [Google Scholar] [CrossRef]
- Combs, R.L.; Valerio, J.R. Biology of the fall armyworm on four varieties of bermudagrass when held at constant temperatures. Environ. Entomol. 1980, 9, 393–396. [Google Scholar] [CrossRef]
- Barfield, C.S.; Ashley, T.R. Effects of corn phenology and temperature on the life cycle of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Fla. Entomol. 1987, 70, 110–116. [Google Scholar] [CrossRef]
- Barros, E.M.; Torres, J.B.; Ruberson, J.R.; Oliveira, M.D. Development of Spodoptera frugiperda on different hosts and damage to reproductive structures in cotton. Entomol. Exp. Appl. 2010, 137, 237–245. [Google Scholar] [CrossRef]
- Chen, Y.; Guo, J.F.; Gao, Z.P.; He, K.; Bai, S.X.; Zhang, T.T.; Wang, Z.Y. Performance of Spodoptera frugiperda (Lepidoptera: Noctuidae) fed on six host plants: Potential risks to mid-high latitude crops in China. J. Agric. Sci. 2020, 12, 16. [Google Scholar] [CrossRef]
- Wu, L.H.; Zhou, C.; Long, G.Y.; Yang, X.B.; Wei, Z.Y.; Liao, Y.J.; Yang, H.; Hu, C.X. Fitness of fall armyworm, Spodoptera frugiperda to three solanaceous vegetables. J. Integr. Agric. 2021, 20, 755–763. [Google Scholar] [CrossRef]
- Tian, T.A.; Yu, L.C.; Sun, G.J.; Yu, X.F.; Li, L.T.; Wu, C.X.; Chen, Y.C.; Yang, M.F.; Liu, J.F. Biological control efficiency of an ectoparasitic mite Pyemotes zhonghuajia on oriental armyworm Mythimna separata. Syst. Appl. Acarol. 2020, 25, 1683–1692. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull Inst. Zool Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, TWOSEX-MSChart: A Computer Program for the Age–Stage, Two-Sex Life Table Analysis. 2021. Available online: http://140.120.197.173/Ecology/prod02.htm (accessed on 1 December 2021).
- Chi, H.; Su, H.Y. Age-stage, Two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Akca, I.; Ayvaz, T.; Yazici, E.; Smith, C.L.; Chi, H. Demography and population projection of aphis fabae (Hemiptera: Aphididae): With additional comments on life table research criteria. J. Econ. Entomol. 2015, 108, 1466–1478. [Google Scholar] [CrossRef] [PubMed]
- Plessis, H.D.; Schlemmer, M.L.; Berg, J.V.D. The effect of temperature on the development of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2020, 11, 228. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.G.; Godoy, W.A.C.; Thomas, J.M.G.; Nagoshi, R.N.; Meagher, R.L. Delimiting strategic zones for the development of fall armyworm (Lepidoptera: Noctuidae) on corn in the state of Florida. J. Econ. Entomol. 2017, 111, 120–126. [Google Scholar] [CrossRef]
- Huang, L.L.; Xue, F.S.; Chen, C.; Guo, X.; Tang, J.J.; Zhong, L.; He, H.M. Effects of temperature on life-history traits of the newly invasive fall armyworm, Spodoptera frugiperda in Southeast China. Ecol. Evol. 2021, 11, 5255–5264. [Google Scholar] [CrossRef]
- Guo, J.F.; Zhang, M.D.; Gao, Z.P.; Wang, D.; He, K. Comparison of larval performance and oviposition preference of Spodoptera frugiperda among three host plants: Potential risks to potato and tobacco crops. Insect Sci. 2020, 28, 602–614. [Google Scholar] [CrossRef]
- Mello, D.M.D.; Bueno, A.D.F.; Andrade, K.; Stecca, C.D.S.; Oliveira, M.C.N.D. Biology and nutrition of Spodoptera frugiperda (Lepidoptera: Noctuidae) fed on different food source. Sci. Agric. 2017, 74, 18–31. [Google Scholar]
- Aregbesola, O.Z.; Legg, J.P.; Lund, O.S.; Sigsgaard, L.; Sporleder, M.; Carhuapoma, P.; Rapisarda, C. Life history and temperature-dependence of cassava-colonising populations of Bemisia tabaci. J. Pest Sci. 2020, 93, 1225–1241. [Google Scholar] [CrossRef]
- Wang, W.W.; He, P.Y.; Zhang, Y.Y.; Liu, T.X.; Jing, X.F.; Zhang, S.Z. The population growth of Spodoptera frugiperda on six cash crop species and implications for its occurrence and damage potential in China. Insects 2020, 11, 639. [Google Scholar] [CrossRef] [PubMed]
- Winder, M.; Jassby, A.D.; Mac-Nally, R. Synergies between climate anomalies and hydrological modifications facilitate estuarine biotic invasions. Ecol. Lett. 2011, 14, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Keosentse, O.; Mutamiswa, R.; Plessis, H.D.; Nyamukondiwa, C. Developmental stage variation in Spodoptera frugiperda (Lepidoptera: Noctuidae) low temperature tolerance: Implications for overwintering. Austral Entomol. 2021, 60, 400–410. [Google Scholar] [CrossRef]
- Zhang, D.D.; Zhao, S.Y.; Wu, Q.L.; Li, Y.Y.; Wu, K.M. Cold hardiness of the invasive fall armyworm, Spodoptera frugiperda in China. J. Integr. Agric. 2021, 20, 764–771. [Google Scholar] [CrossRef]
- Morey, A.C.; Venette, R.C.; Nystrom, E.C.N.; Mosca, L.A.; Hutchison, W.D. Host-mediated shift in the cold tolerance of an invasive insect. Ecol. Evol. 2016, 6, 8267–8275. [Google Scholar] [CrossRef]
- Pelini, S.L.; Keppel, J.A.; Kelley, A.E.; Hellmann, J.J. Adaptation to host plants may prevent rapid insect responses to climate change. Glob. Chang. Biol. 2010, 16, 2923–2929. [Google Scholar] [CrossRef]
- McCalla, K.A.; Keçeci, M.; Milosavljević, I.; Ratkowsky, D.A.; Hoddle, M.S. The influence of temperature variation on life history parameters and thermal performance curves of Tamarixia radiata (Hymenoptera: Eulophidae), a parasitoid of the Asian citrus psyllid (Hemiptera: Liviidae). J. Econ. Entomol. 2019, 112, 1560–1572. [Google Scholar] [CrossRef]
- Milosavljević, I.; McCalla, K.A.; Ratkowsky, D.A.; Hoddle, M.S. Effects of constant and fluctuating temperatures on development rates and longevity of Diaphorencyrtus Aligarhensis (Hymenoptera: Encyrtidae). J. Econ.Entomol. 2019, 112, 1062–1072. [Google Scholar] [CrossRef]
Figure 1.
Age-stage specific survival rate (sxj) of each developmental stage of Spodoptera frugiperda feeding on different host plants at different temperatures.
Figure 1.
Age-stage specific survival rate (sxj) of each developmental stage of Spodoptera frugiperda feeding on different host plants at different temperatures.

Figure 2.
Age-stage specific survival rate (lx), fecundity (mx), and maternity (lxmx) of Spodoptera frugiperda on different hosts and at different temperatures.
Figure 2.
Age-stage specific survival rate (lx), fecundity (mx), and maternity (lxmx) of Spodoptera frugiperda on different hosts and at different temperatures.

Figure 3.
Age-stage specific life expectancy (exj) of each development stage of Spodoptera frugiperda reared on different hosts at different temperatures.
Figure 3.
Age-stage specific life expectancy (exj) of each development stage of Spodoptera frugiperda reared on different hosts at different temperatures.

Figure 4.
Age-stage reproductive value (vxj) of each developmental stage of Spodoptera frugiperda on different hosts and at different temperatures.
Figure 4.
Age-stage reproductive value (vxj) of each developmental stage of Spodoptera frugiperda on different hosts and at different temperatures.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Mortality of mean (±SE) different developmental stages of Spodoptera frugiperda reared on different host plants at different temperatures.
Table 1.
Mortality of mean (±SE) different developmental stages of Spodoptera frugiperda reared on different host plants at different temperatures.
| Mortality Rate | ||||||
|---|---|---|---|---|---|---|
| Temperature (°C) | Egg | 1st instar Larva | ||||
| Maize | Sorghum | Coix seed | Maize | Sorghum | Coix seed | |
| 20 | 0.29 ± 0.045 aA | 0.35 ± 0.048 aA | 0.42 ± 0.049 aA | 0 ± 0 aA | 0 ± 0 aA | 0 ± 0 aA |
| 25 | 0.27 ± 0.044 aA | 0.37 ± 0.05 aA | 0.35 ± 0.048 aAB | 0 ± 0 aA | 0 ± 0 aA | 0 ± 0 aA |
| 30 | 0.21 ± 0.041 aA | 0.29 ± 0.046 aA | 0.28 ± 0.045 aB | 0 ± 0 aA | 0 ± 0 aA | 0 ± 0 aA |
| Temperature (°C) | 2nd instar larva | 3rd instar larva | ||||
| Maize | Sorghum | Coix seed | Maize | Sorghum | Coix seed | |
| 20 | 0.03 ± 0.017 aA | 0.06 ± 0.024 aA | 0.03 ± 0.017 aA | 0.03 ± 0.017 aA | 0.04 ± 0.02 aA | 0.03 ± 0.017 aA |
| 25 | 0.02 ± 0.014 aA | 0.05 ± 0.022 aA | 0.07 ± 0.026 aA | 0.01 ± 0.009 abA | 0 ± 0 bB | 0.04 ± 0.02 aA |
| 30 | 0.02 ± 0.014 aA | 0.05 ± 0.022 aA | 0.05 ± 0.022 aA | 0 ± 0 aA | 0.03 ± 0.017 aAB | 0.02 ± 0.014 aA |
| Temperature (°C) | 4th instar larva | 5th instar larva | ||||
| Maize | Sorghum | Coix seed | Maize | Sorghum | Coix seed | |
| 20 | 0.04 ± 0.02 bA | 0 ± 0 aB | 0 ± 0 aA | 0.02 ± 0.014 aAB | 0.04 ± 0.02 aA | 0.04 ± 0.02 aA |
| 25 | 0.03 ± 0.017 aAB | 0 ± 0 aB | 0 ± 0 aA | 0 ± 0 aB | 0 ± 0 aB | 0.02 ± 0.014 aA |
| 30 | 0 ± 0 bB | 0.06 ± 0.024 aA | 0.02 ± 0.014 bA | 0.05 ± 0.022 aA | 0.01 ± 0.01 bAB | 0.01 ± 0.01 bA |
| Temperature (°C) | 6th instar larva | Pre-pupa | ||||
| Maize | Sorghum | Coix seed | Maize | Sorghum | Coix seed | |
| 20 | 0 ± 0 aA | 0 ± 0 aA | 0.01 ± 0.01 aA | 0 ± 0 aA | 0.03 ± 0.017 aA | 0.02 ± 0.014 aB |
| 25 | 0 ± 0 bA | 0.03 ± 0.017 aA | 0.03 ± 0.017 aA | 0.01 ± 0.01 aA | 0.01 ± 0.01 aA | 0 ± 0 aB |
| 30 | 0 ± 0 aA | 0 ± 0 aA | 0 ± 0 aA | 0 ± 0 bA | 0.02 ± 0.014 bA | 0.07 ± 0.026 aA |
| Temperature (°C) | Pupa | Pre-adult | ||||
| Maize | Sorghum | Coix seed | Maize | Sorghum | Coix seed | |
| 20 | 0.03 ± 0.017 aA | 0 ± 0 aA | 0 ± 0 aB | 0.44 ± 0.05 aA | 0.52 ± 0.05 aA | 0.55 ± 0.05 aA |
| 25 | 0.01 ± 0.01 bA | 0.02 ± 0.014 bA | 0.08 ± 0.027 aA | 0.35 ± 0.048 bAB | 0.48 ± 0.05 bA | 0.59 ± 0.05 aA |
| 30 | 0 ± 0 aA | 0.02 ± 0.014 aA | 0.02 ± 0.014 aB | 0.28 ± 0.045 bB | 0.48 ± 0.05 aA | 0.47 ± 0.05 aA |
The lowercase letters (a–c) indicate significant differences in mortality at different developmental stages of Spodoptera frugiperda on different hosts under the same temperature. The capital letters (A–C) indicate significant differences in mortality at different developmental stages of S. frugiperda at different temperatures with the same host plant. A paired bootstrap test was used to detect statistical differences in the mortality at different stages of S. frugiperda on different host plants or different temperatures. Standard errors were estimated from 100,000 bootstrap resampling.
Table 2.
Duration of the mean (±SE) each developmental stage of Spodoptera frugiperda reared on different host plants under different temperatures.
Table 2.
Duration of the mean (±SE) each developmental stage of Spodoptera frugiperda reared on different host plants under different temperatures.
| Developmental Time (d) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temperature (°C) | Egg | 1st instar larva | ||||||||||
| N | Maize | N | Sorghum | N | Coix seed | N | Maize | N | Sorghum | N | Coix seed | |
| 20 | 71 | 5.11 ± 0.02 aA | 65 | 5.15 ± 0.03 aA | 58 | 5.2 ± 0.03 aA | 71 | 3.97 ± 0.05 aA | 65 | 3.96 ± 0.06 aA | 58 | 3.92 ± 0.06 aA |
| 25 | 73 | 3.41 ± 0.04 aB | 63 | 3.30 ± 0.04 aB | 65 | 3.32 ± 0.04 aB | 73 | 2.01 ± 0.05 bB | 63 | 2.06 ± 0.04 bB | 65 | 2.53 ± 0.05 aB |
| 30 | 79 | 1.61 ± 0.02 aC | 71 | 1.62 ± 0.03 aC | 72 | 1.57 ± 0.02 aC | 79 | 1.61 ± 0.02 aC | 71 | 1.62 ± 0.03 aC | 72 | 1.57 ± 0.02 aC |
| Temperature (°C) | 2nd instar larva | 3rd instar larva | ||||||||||
| N | Maize | N | Sorghum | N | Coix seed | N | Maize | N | Sorghum | N | Coix seed | |
| 20 | 68 | 3.29 ± 0.04 aA | 59 | 3.21 ± 0.03 aA | 55 | 3.21 ± 0.03 aA | 65 | 3.58 ± 0.05 aA | 55 | 3.7 ± 0.06 aA | 52 | 3.71 ± 0.06 aA |
| 25 | 71 | 1.98 ± 0.04 aB | 58 | 1.91 ± 0.05abB | 58 | 2.01 ± 0.03 bB | 70 | 1.14 ± 0.03 bB | 58 | 2.63 ± 0.05 aB | 54 | 2.64 ± 0.05 aB |
| 30 | 77 | 1.23 ± 0.03 cC | 66 | 1.33 ± 0.03 bC | 67 | 1.46 ± 0.05 aC | 77 | 1.32 ± 0.03 bC | 63 | 1.24 ± 0.03 bC | 65 | 1.45 ± 0.04 aC |
| Temperature (°C) | 4th instar larva | 5th instar larva | ||||||||||
| N | Maize | N | Sorghum | N | Coix seed | N | Maize | N | Sorghum | N | Coix seed | |
| 20 | 61 | 3.32 ± 0.04 aA | 55 | 3.38 ± 0.05 aA | 52 | 3.38 ± 0.05 aA | 59 | 3.54 ± 0.04 aA | 51 | 3.54 ± 0.06 aA | 48 | 3.46 ± 0.06 aA |
| 25 | 67 | 1.84 ± 0.03 cB | 58 | 2.17 ± 0.03 bB | 54 | 2.58 ± 0.04 aB | 67 | 2.43 ± 0.06 bB | 58 | 2.78 ± 0.06 aB | 52 | 2.88 ± 0.06 aB |
| 30 | 77 | 0.96 ± 0.02 bC | 57 | 1.18 ± 0.03 aC | 63 | 1.18 ± 0.03 aC | 72 | 1.08 ± 0.02 bC | 56 | 1.12 ± 0.03 bC | 62 | 1.59 ± 0.03 aC |
| Temperature (°C) | 6th instar larva | Pre-pupa | ||||||||||
| N | Maize | N | Sorghum | N | Coix seed | N | Maize | N | Sorghum | N | Coix seed | |
| 20 | 59 | 3.18 ± 0.03 aA | 51 | 3.14 ± 0.05 aA | 47 | 3.2 ± 0.05 aA | 59 | 3.01 ± 0.04 bA | 48 | 3.17 ± 0.05 aA | 45 | 3.2 ± 0.05 aA |
| 25 | 67 | 2.67 ± 0.05 aB | 55 | 2.81 ± 0.07 aB | 49 | 2.8 ± 0.07 aB | 66 | 2.8 ± 0.06 aB | 54 | 2.96 ± 0.06 aB | 49 | 2.89 ± 0.07 aB |
| 30 | 72 | 1.34 ± 0.05 bC | 56 | 1.93 ± 0.04 aC | 62 | 1.93 ± 0.02 aC | 72 | 1.76 ± 0.04 bC | 54 | 2.36 ± 0.07 aC | 55 | 2.27 ± 0.07 aC |
| Temperature (°C) | Pupa | Pre-adult | ||||||||||
| N | Maize | N | Sorghum | N | Coix seed | N | Maize | N | Sorghum | N | Coix seed | |
| 20 | 56 | 18.36 ± 0.17 bA | 48 | 21.47 ± 0.24 aA | 45 | 21.53 ± 0.23 aA | 56 | 47.34 ± 0.19 bA | 48 | 50.75 ± 0.3 aA | 45 | 50.82 ± 0.28 aA |
| 25 | 65 | 12.83 ± 0.08 bB | 52 | 13.92 ± 0.13 aB | 41 | 13.56 ± 0.33 aB | 65 | 31.09 ± 0.17 bB | 52 | 34.59 ± 0.2 aB | 41 | 35.02 ± 0.32 aB |
| 30 | 72 | 7.69 ± 0.11 bC | 52 | 8.87 ± 0.09 aC | 53 | 7.48 ± 0.09 bC | 72 | 18.59 ± 0.15 cC | 52 | 21.3 ± 0.13 aC | 53 | 20.42 ± 0.17 bC |
The lowercase letters (a–c) indicate significant differences in the developmental duration of Spodoptera frugiperda on different hosts at the same temperature. The capital letters (A–C) indicate significant differences in the developmental duration of S. frugiperda at different temperatures on the same host. A paired bootstrap test was used to detect statistical differences in the mortality at different stages of S. frugiperda on different host plants or different temperatures. Standard errors were estimated using 100,000 bootstrap resampling.
Table 3.
Adult longevity and reproductive parameters mean (±SE) of Spodoptera frugiperda reared on different host plants under different temperatures.
Table 3.
Adult longevity and reproductive parameters mean (±SE) of Spodoptera frugiperda reared on different host plants under different temperatures.
| Temperature (°C) | Adult Longevity/d (Female) | Adult Longevity/d (Male) | ||||
| Maize | Sorghum | Coix seed | Maize | Sorghum | Coix seed | |
| 20 | 57.2 ± 0.54 bA | 60.53 ± 0.64 aA | 60.34 ± 0.94 aA | 56.1 ± 0.63 bA | 59.95 ± 0.65 aA | 59.4 ± 0.62 aA |
| 25 | 39.87 ± 0.33 bB | 43.94 ± 0.5 aB | 43.4 ± 0.46 aB | 39.41 ± 0.37 bB | 41.87 ± 0.41 aB | 42.19 ± 0.62 aB |
| 30 | 27.18 ± 0.28 cC | 29.48 ± 0.33 aC | 28.34 ± 0.4 bC | 26.1 ± 0.32 cC | 29.05 ± 0.41 aC | 27.89 ± 0.41 bC |
| Temperature (°C) | Total pre-oviposition period (TPOP)/d | Adult pre-oviposition period (APOP)/d | ||||
| Maize | Sorghum | Coix seed | Maize | Sorghum | Coix seed | |
| 20 | 51.72 ± 0.33 bA | 56 ± 0.26 aA | 55.14 ± 1.14 aA | 5.22 ± 0.15 aA | 5.5 ± 0.34 aA | 5.14 ± 0.14 aA |
| 25 | 32.62 ± 0.5 bB | 36.35 ± 0.65 aB | 36.67 ± 0.49 aB | 1.83 ± 0.21 aB | 2.3 ± 0.4 aB | 1.67 ± 0.21 aB |
| 30 | 21.14 ± 0.17 cC | 23.86 ± 0.39 aC | 22.67 ± 0.42 bC | 2.17 ± 0.19 aB | 2.43 ± 0.25 aB | 2.42 ± 0.26 aB |
| Temperature (°C) | Oviposition days/Od | Fecundity (no. of eggs) | ||||
| Maize | Sorghum | Coix seed | Maize | Sorghum | Coix seed | |
| 20 | 1.78 ± 0.15 aC | 1.83 ± 0.17 aB | 1.86 ± 0.14 aB | 404.56.95 ± 49.16 aB | 426.83 ± 76.68 aA | 415.71 ± 73.41 aB |
| 25 | 3.92 ± 0.29 aA | 3.4 ± 0.34 aA | 3.33 ± 0.21aA | 954.92 ± 52.83 aA | 626.87 ± 95.21 bA | 628.17 ± 52.01 bA |
| 30 | 2.61 ± 0.31 aB | 2.71 ± 0.27 aA | 2.83 ± 0.27 aA | 956.83 ± 93.5 aA | 615 ± 77.97 bA | 558.42 ± 66.22 bAB |
The lowercase letters (a–c) indicate significant differences in the developmental duration of Spodoptera frugiperda on different hosts at the same temperature. The capital letters (A–C) indicate significant differences in the developmental duration of S. frugiperda at different temperatures on the same host. A paired bootstrap test was used to detect statistical differences in the mortality at different stages of S. frugiperda on different host plants or different temperatures. Standard errors were estimated using 100,000 bootstrap resampling.
Table 4.
The intrinsic rate of increase (r), finite rate of increase (λ), net reproductive rate (R0), mean generation time (T), and gross reproductive rate (GRR) (±SE) of Spodoptera frugiperda reared on different host plants at different temperatures.
Table 4.
The intrinsic rate of increase (r), finite rate of increase (λ), net reproductive rate (R0), mean generation time (T), and gross reproductive rate (GRR) (±SE) of Spodoptera frugiperda reared on different host plants at different temperatures.
| Temperature (°C) | r (d−1) | λ (d−1) | ||||
| Maize | Sorghum | Coix seed | Maize | Sorghum | Coix seed | |
| 20 | 0.068 ± 0.007 aC | 0.057 ± 0.009 aC | 0.06 ± 0.009 aC | 1.07 ± 0.008 aC | 1.058 ± 0.009 aC | 1.062 ± 0.009 aC |
| 25 | 0.137 ± 0.009 aB | 0.108 ± 0.009 bB | 0.094 ± 0.012 bB | 1.146 ± 0.01 aB | 1.114 ± 0.01 bB | 1.099 ± 0.013 bB |
| 30 | 0.224 ± 0.011 aA | 0.176 ± 0.012 bA | 0.173 ± 0.013 bA | 1.251 ± 0.013 aA | 1.192 ± 0.014 bA | 1.189 ± 0.016 bA |
| Temperature (°C) | R0 (offspring) | T (d) | ||||
| Maize | Sorghum | Coix seed | Maize | Sorghum | Coix seed | |
| 20 | 36.41 ± 12.309 aB | 25.61 ± 10.913 aB | 29.1 ± 11.64 aA | 52.843 ± 0.383 bA | 57.38 ± 0.316 aA | 55.743 ± 1.47 aA |
| 25 | 114.59 ± 31.535 aA | 62.687 ± 20.821 abAB | 37.69 ± 15.065 bA | 34.7 ± 0.496 bB | 38.39 ± 0.599 aB | 38.48 ± 0.494 aB |
| 30 | 172.23 ± 40.121 aA | 86.1 ± 23.755 abA | 67.01 ± 19.699 bA | 23.017 ± 0.176 bC | 25.338 ± 0.294 aC | 24.353 ± 0.429 aB |
| Temperature (°C) | GRR (offspring) | |||||
| Maize | Sorghum | Coix seed | ||||
| 20 | 76.67 ± 25.008 aB | 67.04 ± 27.34 aB | 96.65 ± 38.96 aA | |||
| 25 | 193.03 ± 51.027 aA | 141.88 ± 46.353 aAB | 100.01 ± 38.056 aA | |||
| 30 | 265.07 ± 59.009 aA | 179.88 ± 45.955 aA | 161.22 ± 47.385 aA | |||
The lowercase letters (a–c) indicate significant differences in the developmental duration of Spodoptera frugiperda on different hosts at the same temperature. The capital letters (A–C) indicate significant differences in the developmental duration of S. frugiperda at different temperatures on the same host. A paired bootstrap test was used to detect statistical differences in the mortality at different stages of S. frugiperda on different host plants or different temperatures. Standard errors were estimated using 100,000 bootstrap resampling.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Chen, Y.-C.; Chen, D.-F.; Yang, M.-F.; Liu, J.-F. The Effect of Temperatures and Hosts on the Life Cycle of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 211. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020211
AMA Style
Chen Y-C, Chen D-F, Yang M-F, Liu J-F. The Effect of Temperatures and Hosts on the Life Cycle of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects. 2022; 13(2):211. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020211
Chicago/Turabian StyleChen, Yi-Chai, De-Fei Chen, Mao-Fa Yang, and Jian-Feng Liu. 2022. "The Effect of Temperatures and Hosts on the Life Cycle of Spodoptera frugiperda (Lepidoptera: Noctuidae)" Insects 13, no. 2: 211. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13020211
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.