
Two New Synonyms of Paraleuctra orientalis (Chu, 1928) (Plecoptera: Leuctridae) Based on Morphological and Molecular Data, with Notes on Paraleuctra cervicornis Du and Qian, 2012


Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Specimen Preparation
2.2. Observation and Description
2.3. DNA Extraction, Amplification and Sequencing
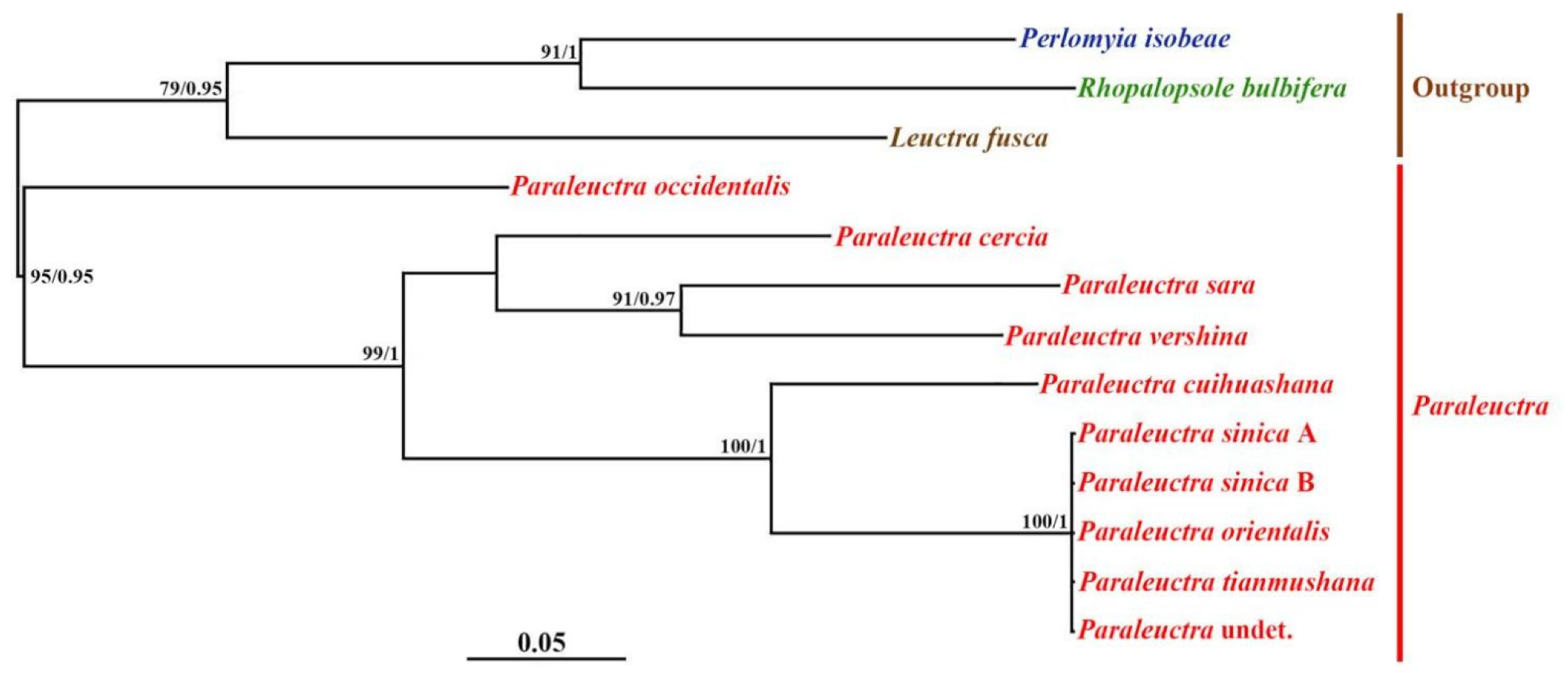
2.4. Phylogenetic Analyses
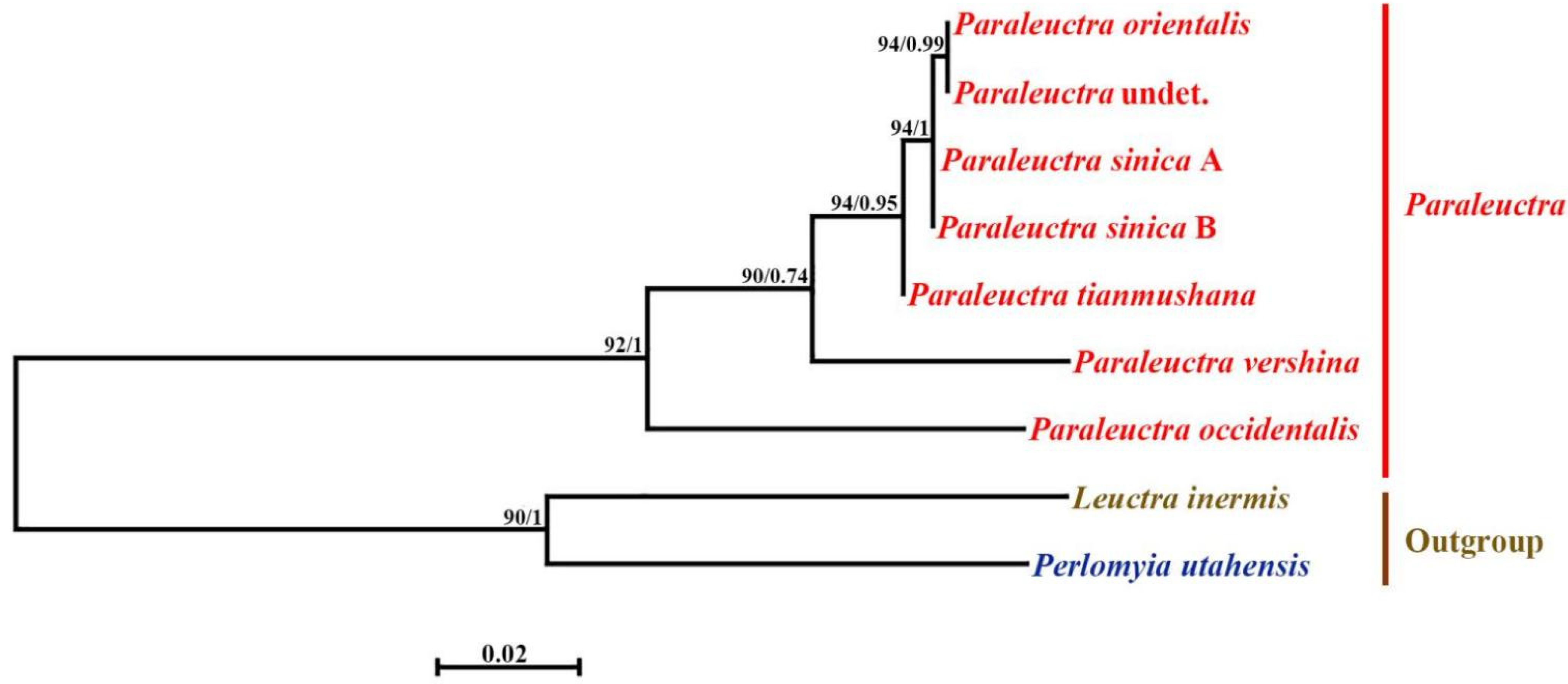
3. Results
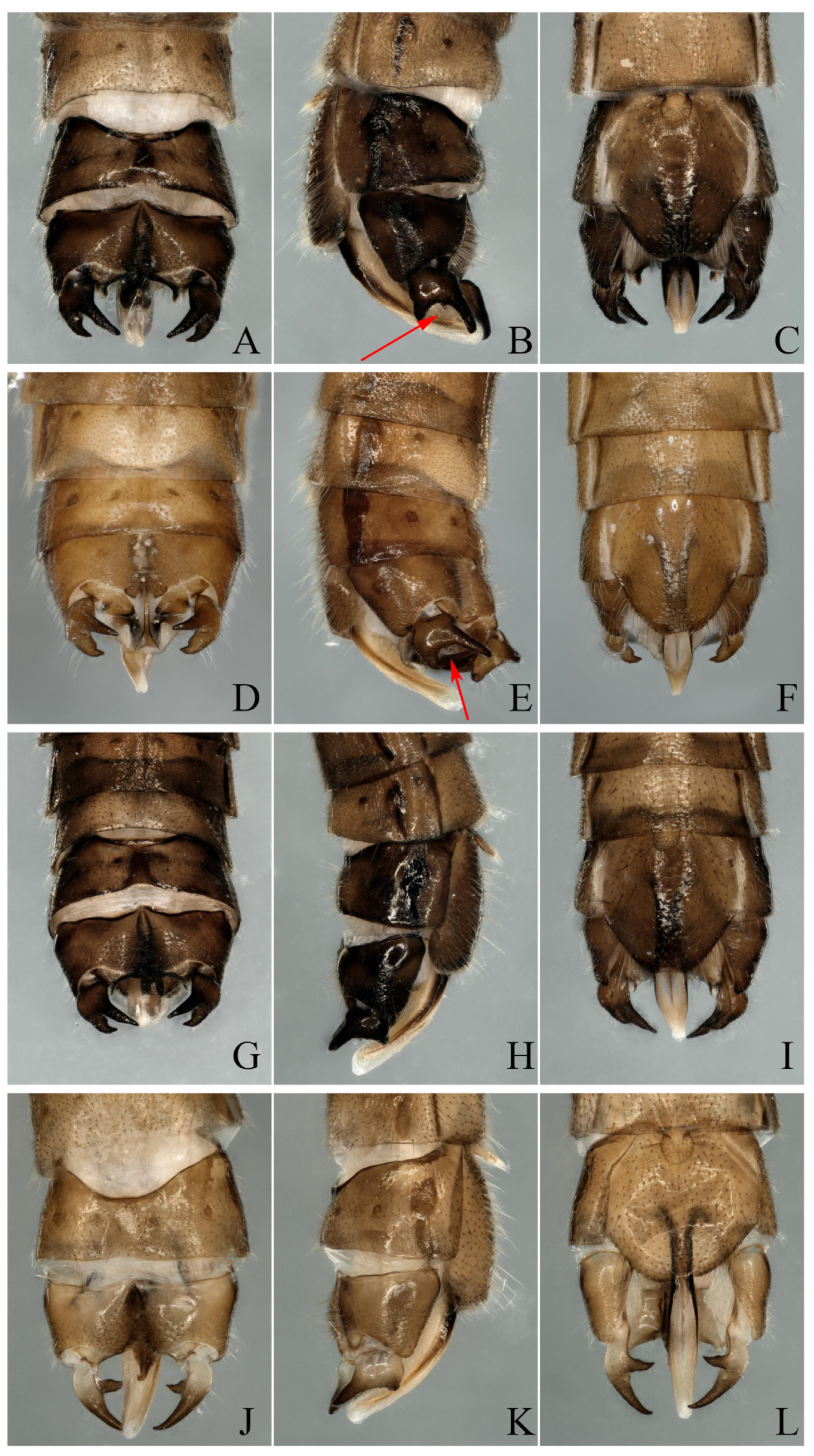
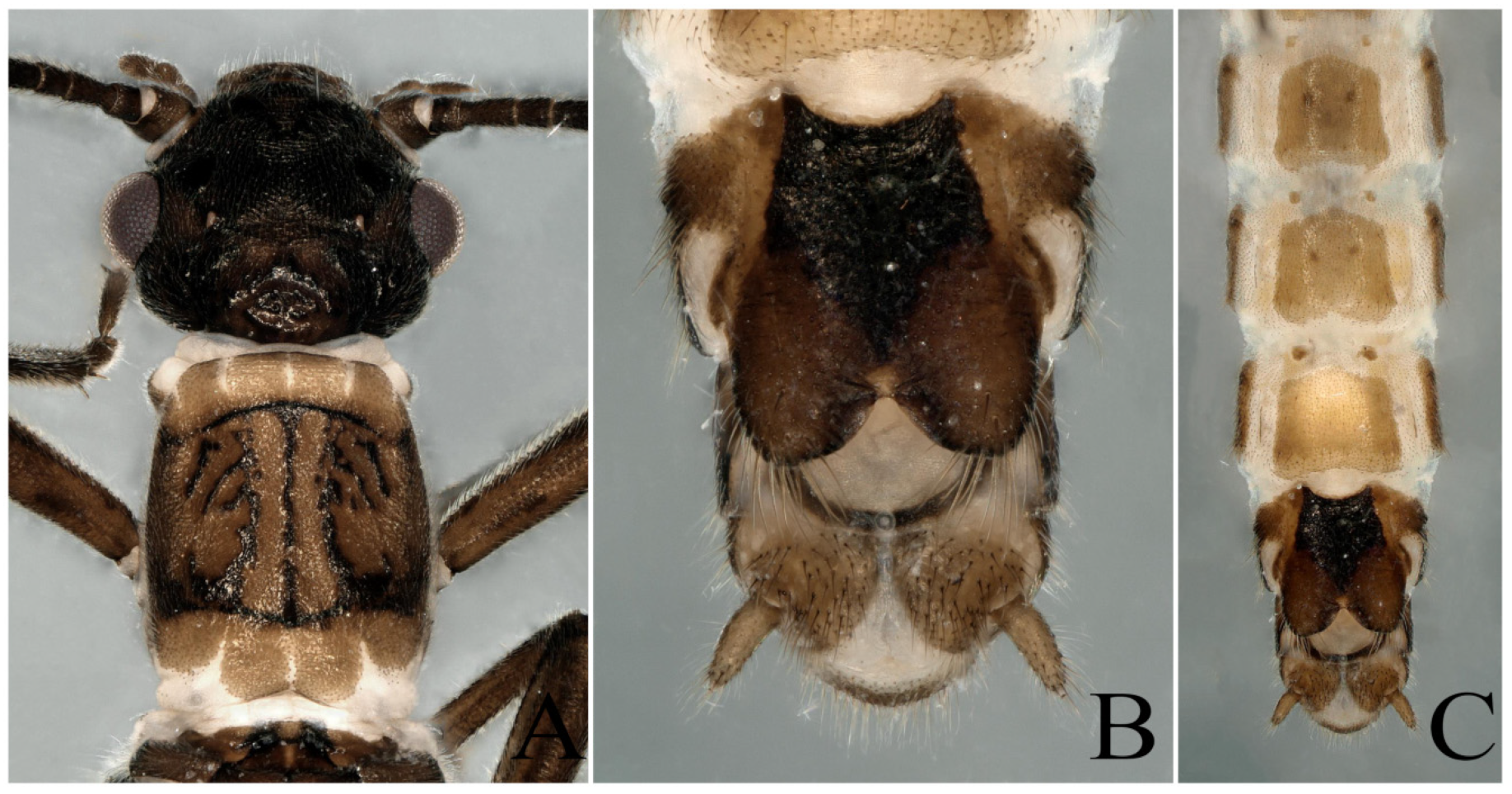
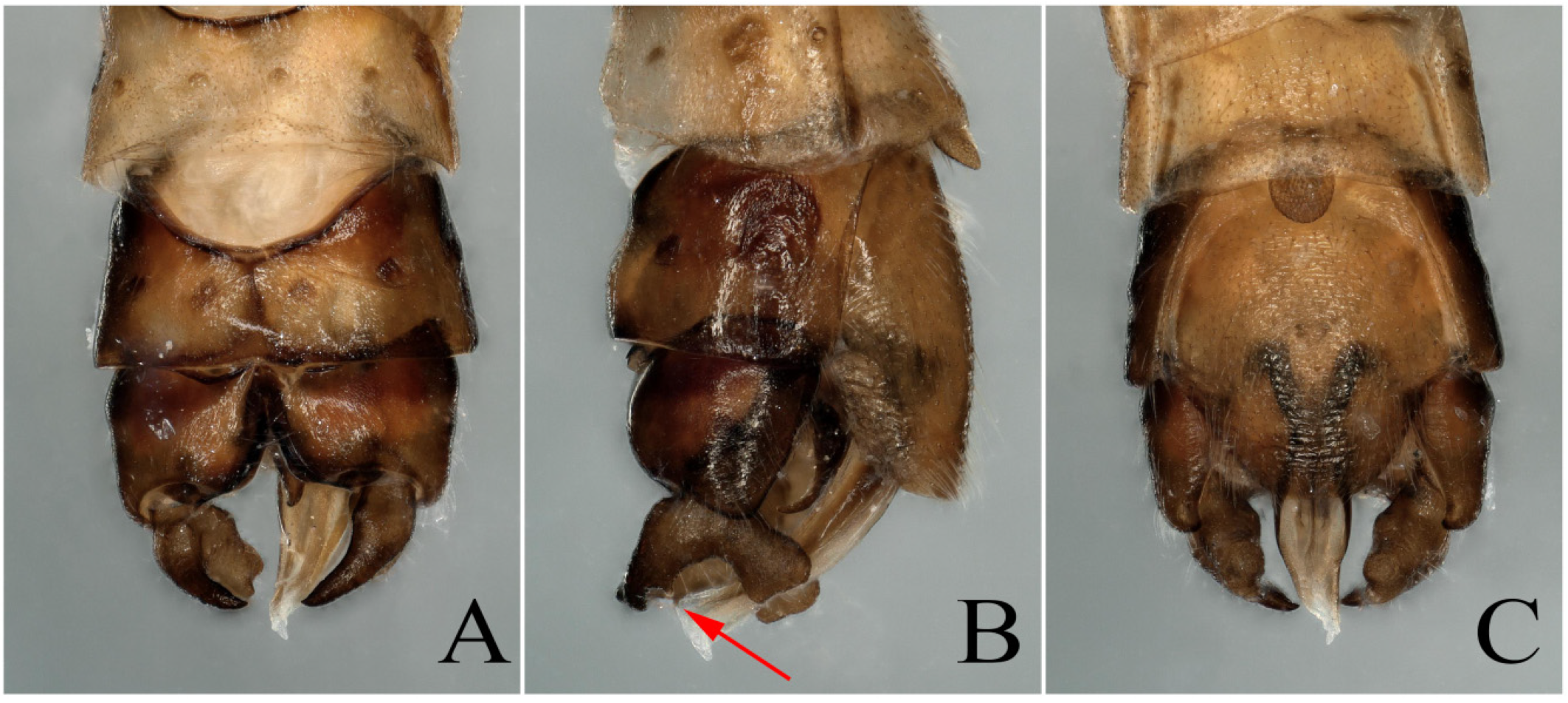
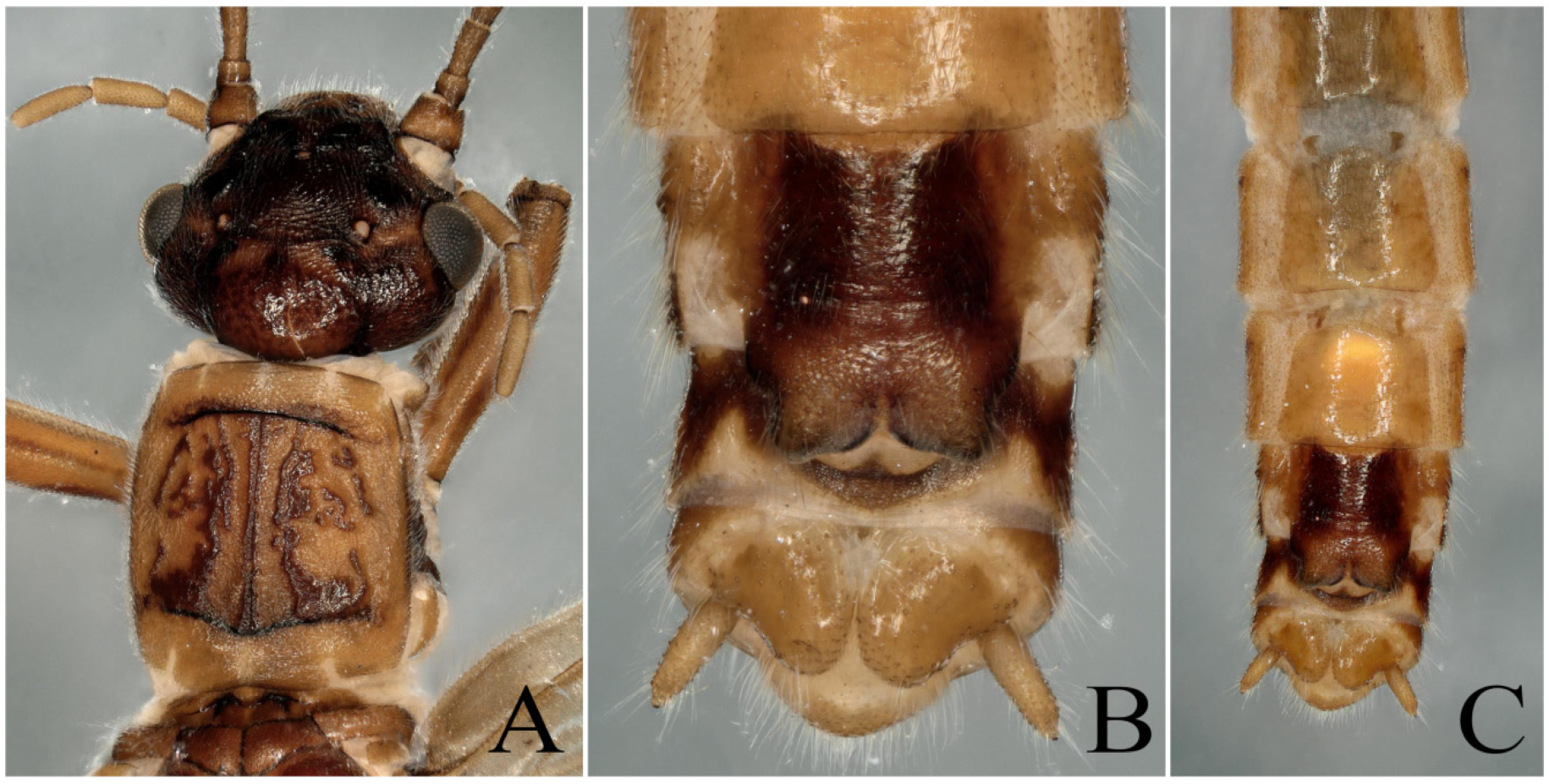
3.1. Paraleuctra orientalis (Chu, 1928)
3.2. Paraleuctra cervicornis Du and Qian, 2012
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- DeWalt, R.E.; Maehr, M.D.; Neu-Becker, U.; Stueber, G. Plecoptera Species File Online. Available online: http://Plecoptera.SpeciesFile.org (accessed on 26 February 2022).
- Chu, Y.T. Description of a new species of Leuctra and notes on Nemoura sinensis from Hangchow. China J. 1928, 9, 87–89. [Google Scholar]
- Hanson, J.F. Studies on the Plecoptera of North America. Bullet. Brooklyn Entomol. Soc. 1941, 36, 57–66. [Google Scholar]
- Yang, D.; Yang, C.K. Plecoptera: Leuctridae. Insects and Macrofungi of Gutianshan, Zhejiang; Zhu, T., Ed.; Zhejiang Science and Technology Press: Hangzhou, China, 1995; pp. 20–24. [Google Scholar]
- Li, W.H.; Wang, Y.B.; Yang, D. Synopsis of the genus Paraleuctra (Plecoptera: Leuctridae) from China. Zootaxa 2010, 2350, 46–52. [Google Scholar] [CrossRef]
- Du, Y.Z.; Qian, Y.H. Paraleuctra cervicornis (Plecoptera: Leuctridae), a new stonefly from China. J. Nat. Hist. 2012, 46, 1059–1063. [Google Scholar] [CrossRef]
- Kong, F.B.; Yang, D.; Li, W.H. Discovery of the genus Paraleuctra (Plecoptera: Leuctridae) from Palearctic China, with description of P. qilianshana sp. nov. Zootaxa 2013, 3702, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.T. The first formal report of brachypterous stonefly of Leuctridae (Plecoptera) from China. Zootaxa 2019, 4624, 230–240. [Google Scholar] [CrossRef]
- Zhiltzova, L.A. Rare genera of the family Leuctridae (Insecta: Plecoptera) in the fauna of the USSR. Zool. Zhurnal 1974, 53, 359–364. [Google Scholar]
- Harper, P.P. Capniidae, Leuctridae, and Perlidae (Plecoptera) from Nepal. Orient. Insects 1977, 11, 53–62. [Google Scholar] [CrossRef]
- Shimizu, T. Paraleuctra (Insecta: Plecoptera: Leuctridae) from Japan, with taxonomic notes on the Japanese Leuctridae. Species Divers. 2000, 5, 285–303. [Google Scholar] [CrossRef] [Green Version]
- Stark, B.P.; Juliana, W.K. Systematics of Nearctic Paraleuctra with description of a new genus (Plecoptera: Leuctridae). Tijdschr. Entomol. 2001, 144, 119–135. [Google Scholar] [CrossRef] [Green Version]
- Cherchesova, S.K.; Zhiltzova, L.A. The stonefly fauna (Plecoptera) of North Ossetia and its zoogeographical characteristics. Entomol. Rev. 2003, 83, 451–454. [Google Scholar] [CrossRef] [Green Version]
- Baumann, R.W.; Stark, B.P. Paraleuctra alta (Plecoptera: Leuctridae), a new stonefly from Alberta, Canada. Illiesia 2009, 5, 30–33. [Google Scholar]
- Murányi, D.; Hwang, J.M. Four new species and further contributions to the Leuctridae (Plecoptera) of the Korean Peninsula. Zootaxa 2017, 4282, 43–61. [Google Scholar] [CrossRef]
- William, D.D.; Feltmate, B.W. Chapter V–Order Plecoptera. In Aquatic Insects; CAB International Publish: Wallingford, UK, 1992. [Google Scholar]
- Zwick, P. Variable egg development of Dinocras spp. (Plecoptera, Perlidae) and the stonefly seed banktheory. Freshw. Biol. 1996, 35, 81–100. [Google Scholar] [CrossRef]
- Snellen, R.K.; Stewart, K.W. The life cycle and drumming behavior of Zealeuctra claasseni (Frison) and Zealeuctra hitei (Ricker and Ross) (Plecoptera: Leuctridae) in Texas, USA. Aquatic Insects 1979, 1, 65–89. [Google Scholar] [CrossRef]
- Terry, M.D.; Whiting, M.F. Mantophasmatodea and phylogeny of the lower neopterous insects. Cladistics 2005, 21, 240–257. [Google Scholar] [CrossRef]
- Yang, D.; Li, W.H.; Zhu, F. Fauna Sinica, Insecta. Plecoptera: Nemouroidea; Science Press: Beijing, China, 2015; Volume 58, 518p. [Google Scholar]
- Illies, J. Katalog der rezenten Plecoptera. Das Tierreich 1966, 82, 1–632. [Google Scholar]
- Zwick, P. Insecta: Plecoptera, Phylogenetisches System und Katalog. Das Tierreich 1973, 94, 410. [Google Scholar]
- Du, Y.Z.; Sivec, I. Insect Fauna of Middle-west Qinling Range and South Montains of Gansu Province; Yang, X.K., Ed.; Science Press: Beijing, China, 2005; Volume 40, pp. 38–54. [Google Scholar]
- Qian, Y.H.; Du, Y.Z. A new stonefly species, Rhopalopsole tricuspis (Leuctridae: Plecoptera), and three new records of stoneflies from the Qinling Mountains of Shaanxi, China. J. Insect Sci. 2012, 12, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Qian, Y.H.; Li, H.L.; Du, Y.Z. A Study of Leuctridae (Insecta: Plecoptera) from Shennongjia, Hubei Province, China. Fla. Entomol. 2014, 97, 605–610. [Google Scholar] [CrossRef]
- Li, W.H.; Li, K.F.; Wang, R.F.; Yang, D. The first description of the larvae of the Chinese species Paraleuctra tianmushana Li & Yang (Plecoptera: Leuctridae). Zootaxa 2016, 4061, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.; Ratnasingham, S.; de Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270 (Suppl. 1), 96–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Adamowicz, S.J.; Jacobus, L.M.; DeWalt, R.E.; Hebert, P.D. Towards a comprehensive barcode library for Arctic life-Ephemeroptera, Plecoptera, and Trichoptera of Churchill, Manitoba, Canada. Front. Zool. 2009, 6, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baur, A.; Buschinger, A.; Zimmermann, F.K. Molecular cloning and sequencing of 18S rDNA gene fragments from six different ant species. Insectes Sociaux 1993, 40, 325–335. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | GenBank Accession No. | |
|---|---|---|---|
| COI Gene | 18S rDNA | ||
| Paraleuctra | orientalis | OM836700 | ON130265 |
| Paraleuctra | sinica A | OM836702 | ON130263 |
| Paraleuctra | sinica B | OM203120 | ON130264 |
| Paraleuctra | tianmushana | OM836701 | ON130266 |
| Paraleuctra | sp. F | OM836704 | ON130267 |
| Paraleuctra | cuihuashana | MK995183 | |
| Paraleuctra | cercia | MK492251 | |
| Paraleuctra | vershina | MH840222 | AY521878 |
| Paraleuctra | sara | HQ938243 | |
| Paraleuctra | occidentalis | MG380092 | EF622723 |
| Perlomyia | isobeae | MK492252 | |
| Perlomyia | utahensis | EF622724 | |
| Rhopalopsole | bulbifera | MK111419 | |
| Leuctra | fusca | MK568475 | |
| Leuctra | inermis | EF622721 | |
| Genetic Distance (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Name | P. orientalis | P. sinica A | P. sinica B | P. tianmushana | P. undet. | P. cuihuashana | P. cercia | P. vershina | P. sara | P. occidentalis | P. isobeae | R. bulbifera |
| P. orientalis | ||||||||||||
| P. sinica A | 0.0 | |||||||||||
| P. sinica B | 0.0 | 0.0 | ||||||||||
| P. tianmushana | 0.0 | 0.0 | 0.0 | |||||||||
| P. undet. | 0.0 | 0.0 | 0.0 | 0.0 | ||||||||
| P. cuihuashana | 11.7 | 11.7 | 11.7 | 11.7 | 11.7 | |||||||
| P. cercia | 18.3 | 18.3 | 18.3 | 18.3 | 18.3 | 15.3 | ||||||
| P. vershina | 17.4 | 17.4 | 17.4 | 17.4 | 17.4 | 17.1 | 14.8 | |||||
| P. sara | 19.5 | 19.5 | 19.5 | 19.5 | 19.5 | 19.4 | 14.7 | 13.0 | ||||
| P. occidentalis | 20.4 | 20.4 | 20.4 | 20.4 | 20.4 | 20.7 | 18.8 | 19.3 | 19.8 | |||
| P. isobeae | 24.3 | 24.3 | 24.3 | 24.3 | 24.3 | 22.8 | 19.5 | 21.7 | 21.1 | 19.0 | ||
| R. bulbifera | 24.9 | 24.9 | 24.9 | 24.9 | 24.9 | 23.2 | 20.0 | 22.7 | 23.3 | 20.5 | 15.4 | |
| L. fusca | 23.0 | 23.0 | 23.0 | 23.0 | 23.0 | 22.6 | 22.6 | 24.4 | 23.6 | 19.6 | 19.9 | 20.1 |
| Genetic Distance (%) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Name | P. orientalis | P. sinica A | P. sinica B | P. tianmushana | P.undet. | P. vershina | P. occidentalis | P. utahensis |
| P. orientalis | ||||||||
| P. sinica A | 0.2 | |||||||
| P. sinica B | 0.2 | 0.0 | ||||||
| P. tianmushana | 0.6 | 0.4 | 0.4 | |||||
| P. undet. | 0.0 | 0.2 | 0.2 | 0.6 | ||||
| P. vershina | 5.6 | 5.4 | 5.4 | 4.9 | 5.6 | |||
| P. occidentalis | 9.5 | 9.3 | 9.3 | 8.8 | 9.5 | 9.0 | ||
| P. utahensis | 30.3 | 30.0 | 30.0 | 29.3 | 30.3 | 30.9 | 29.3 | |
| L. inermis | 28.0 | 27.7 | 27.7 | 27.0 | 28.0 | 24.8 | 26.9 | 14.8 |
| Species Name | Methods | ||||
|---|---|---|---|---|---|
| Morphology | Genetic Distance of the COI Gene | Genetic Distance of the 18S rDNA | Phylogenetic Trees of COI Gene | Phylogenetic Trees of 18S rDNA | |
| P. orientalis | Share similar features such as head, pronotum, male terminalia and female terminalia. | Genetic distance of the COI gene among the three species is below 2%. | Genetic distance of the 18S rDNA is below 2%. | The three species are grouped in the same clade with high support values. | The three species are grouped in the same clade with high support values. |
| P. sinica A | |||||
| P. sinica B | |||||
| P. tianmushana | |||||
| P. undet. | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.-B.; Du, Y.-Z. Two New Synonyms of Paraleuctra orientalis (Chu, 1928) (Plecoptera: Leuctridae) Based on Morphological and Molecular Data, with Notes on Paraleuctra cervicornis Du and Qian, 2012. Insects 2022, 13, 468. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050468
Yang Y-B, Du Y-Z. Two New Synonyms of Paraleuctra orientalis (Chu, 1928) (Plecoptera: Leuctridae) Based on Morphological and Molecular Data, with Notes on Paraleuctra cervicornis Du and Qian, 2012. Insects. 2022; 13(5):468. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050468
Chicago/Turabian StyleYang, Yu-Ben, and Yu-Zhou Du. 2022. "Two New Synonyms of Paraleuctra orientalis (Chu, 1928) (Plecoptera: Leuctridae) Based on Morphological and Molecular Data, with Notes on Paraleuctra cervicornis Du and Qian, 2012" Insects 13, no. 5: 468. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050468