
Molecular Oscillator Affects Susceptibility of Caterpillars to Insecticides: Studies on the Egyptian Cotton Leaf Worm—Spodoptera littoralis (Lepidoptera: Noctuidae)

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
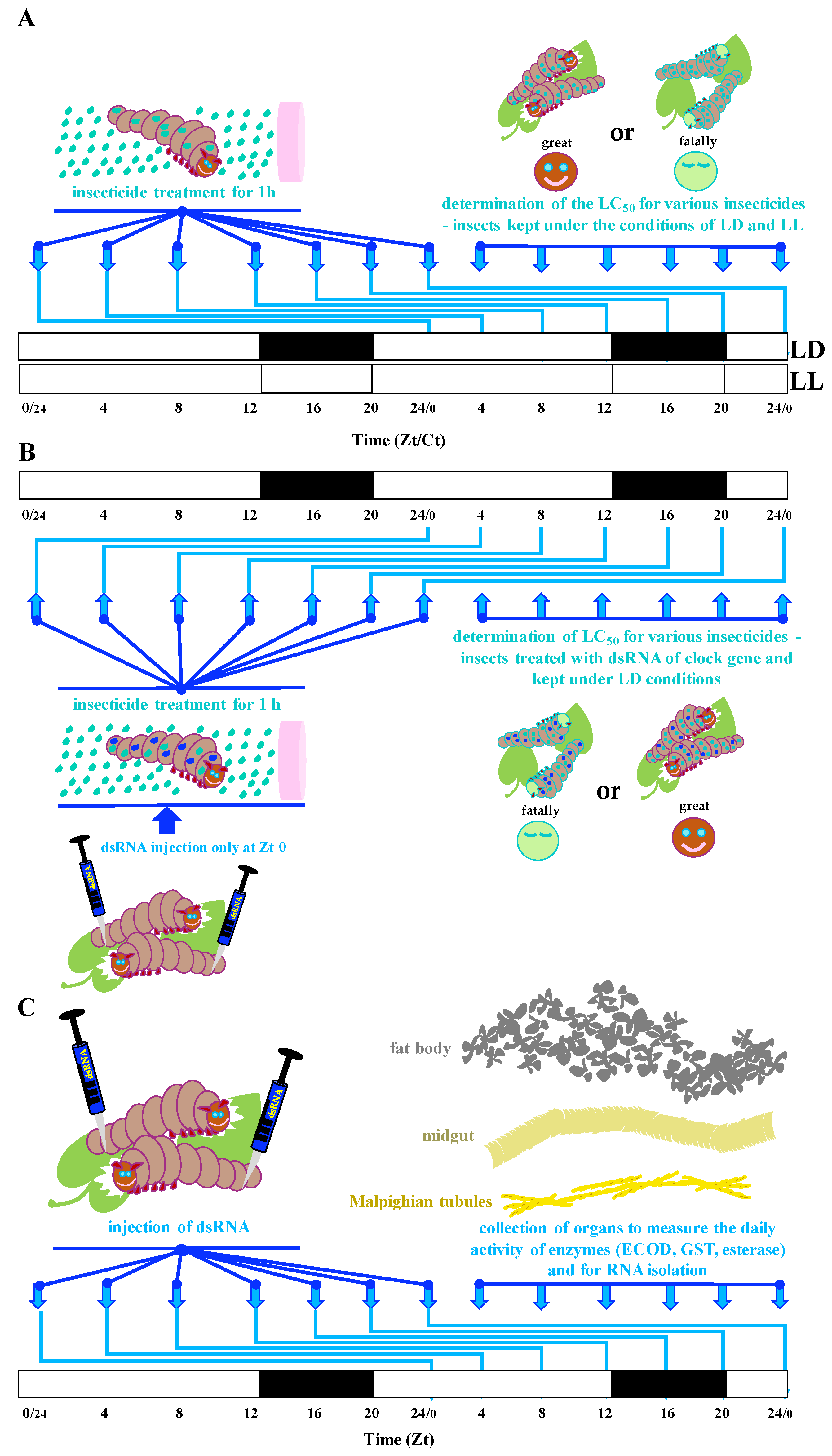
2.1. Insect Breeding
2.2. Insect Treatment with Insecticides
2.3. Insect Treatment with dsRNA
2.4. dsRNA Synthesis
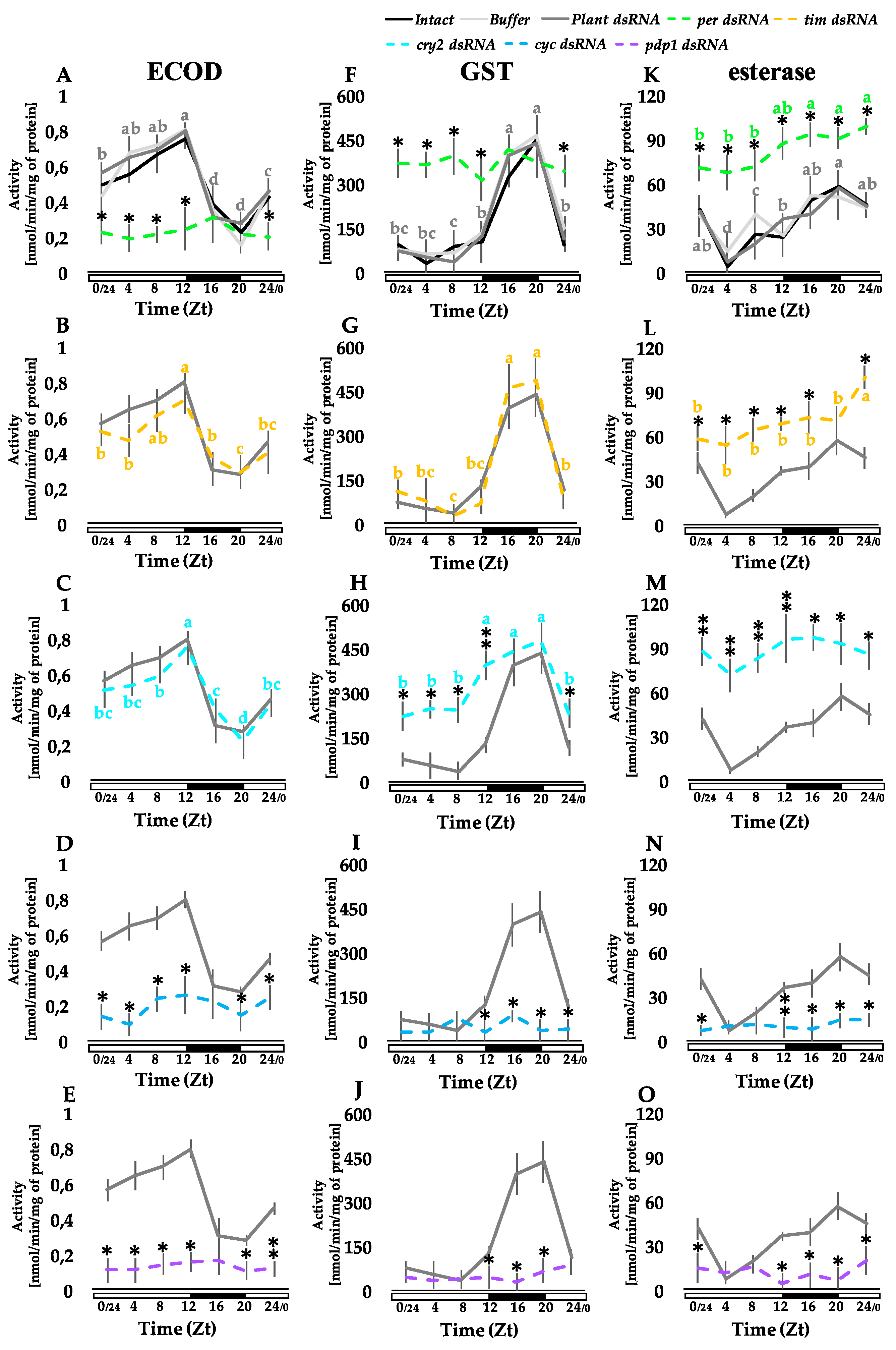
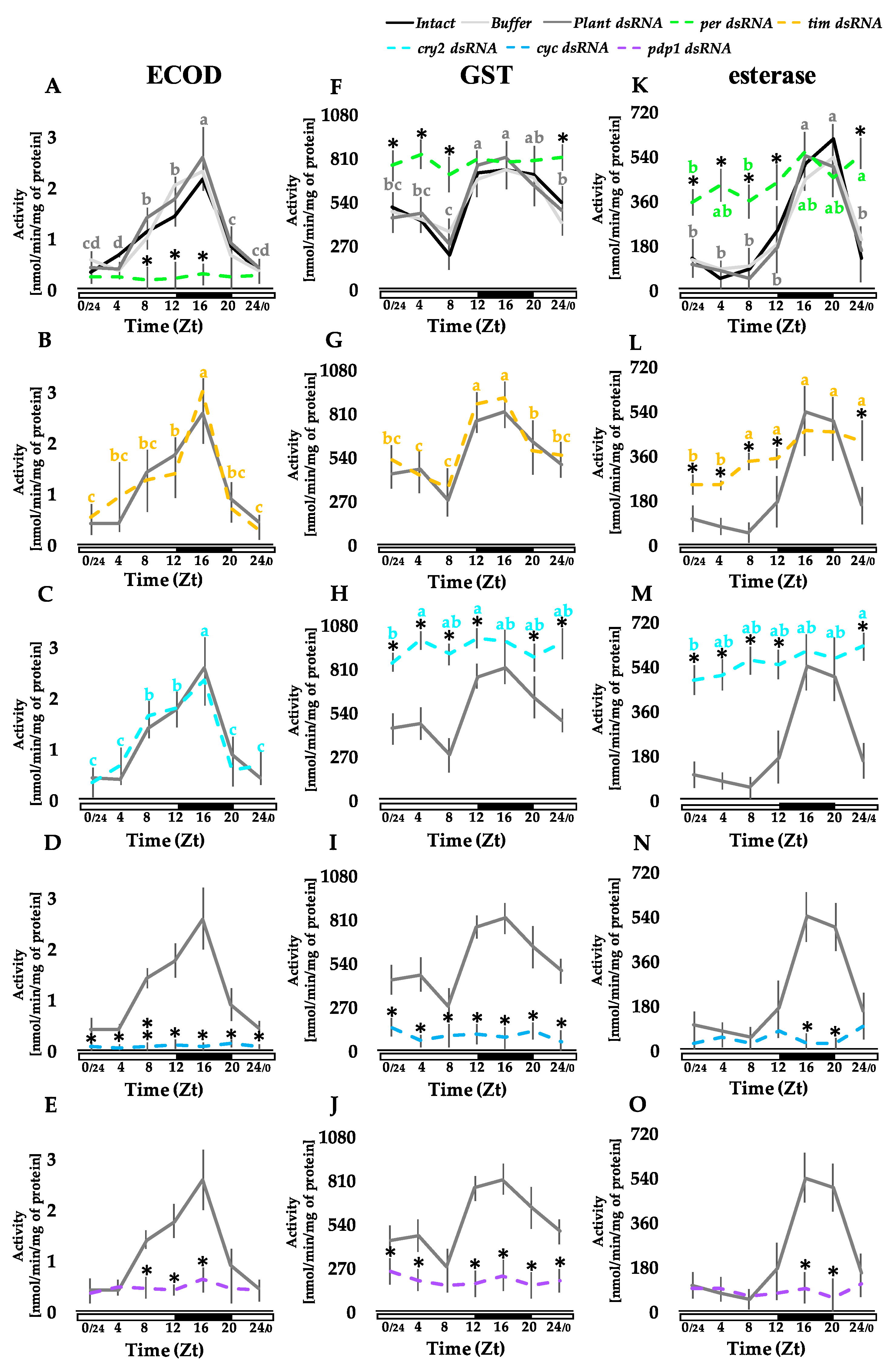
2.5. Enzymatic Assays
2.5.1. 7-Ethoxycoumarin-O-deethylase (ECOD) Activity
2.5.2. Glutathione-S-transferase (GST) Activity
2.5.3. Esterase Activity
3. Results
3.1. Exposure to Continuous Lighting Changes the Larvae’s Susceptibility to Insecticides
3.2. Knocking Out Clock Genes Expression through RNAi Affects the Detoxification Enzymes Activity in the Fat Body, Midgut, and Malpighian Tubules
3.2.1. Fat Body
3.2.2. Midgut
3.2.3. Malpighian Tubules
3.3. Knocking out of the Clock Genes Expression through RNAi Changes the Larvae’s Susceptibility to Insecticides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pilorz, V.; Helfrich-Forster, C.; Oster, H. The role of the circadian clock system in physiology. Pflügers Arch. Eur. J. Physiol. 2018, 470, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Young, M.W. Time Travels: A 40-Year Journey from Drosophila’s Clock Mutants to Human Circadian Disorders (Nobel Lecture). Angew. Chem. Int. Ed. 2018, 57, 11532–11539. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.M.; Loros, J.J.; Dunlap, J.C. Circadian Oscillators: Around the Transcription-Translation Feedback Loop and on to Output. Trends Biochem. Sci. 2016, 41, 834–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saini, R.; Jaskolski, M.; Davis, S.J. Circadian oscillator proteins across the kingdoms of life: Structural aspects. BMC Biol. 2019, 17, 13. [Google Scholar] [CrossRef] [Green Version]
- Tataroglu, O.; Emery, P. The molecular ticks of the Drosophila circadian clock. Curr. Opin. Insect Sci. 2015, 7, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Beer, K.; Helfrich-Forster, C. Model and Non-model Insects in Chronobiology. Front. Behav. Neurosci. 2020, 14, 601676. [Google Scholar] [CrossRef]
- Lim, C.; Chung, B.Y.; Pitman, J.L.; McGill, J.J.; Pradhan, S.; Lee, J.; Keegan, K.P.; Choe, J.; Allada, R. clockwork orange encodes a transcriptional repressor important for circadian-clock amplitude in Drosophila. Curr. Biol. 2007, 17, 1082–1089. [Google Scholar] [CrossRef] [Green Version]
- Tomioka, K.; Matsumoto, A. Circadian molecular clockworks in non-model insects. Curr. Opin. Insect Sci. 2015, 7, 58–64. [Google Scholar] [CrossRef]
- Zhan, S.; Merlin, C.; Boore, J.L.; Reppert, S.M. The Monarch Butterfly Genome Yields Insights into Long-Distance Migration. Cell 2011, 147, 1171–1185. [Google Scholar] [CrossRef] [Green Version]
- Sandrelli, F.; Costa, R.; Kyriacou, C.P.; Rosato, E. Comparative analysis of circadian clock genes in insects. Insect Mol. Biol. 2008, 17, 447–463. [Google Scholar] [CrossRef] [Green Version]
- Derks, M.F.L.; Smit, S.; Salis, L.; Schijlen, E.; Bossers, A.; Mateman, C.; Pijl, A.S.; de Ridder, D.; Groenen, M.A.M.; Visser, M.E.; et al. The Genome of Winter Moth (Operophtera brumata) Provides a Genomic Perspective on Sexual Dimorphism and Phenology. Genome Biol. Evol. 2015, 7, 2321–2332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertossa, R.C.; van de Zande, L.; Beukeboom, L.W.; Beersma, D.G.M. Phylogeny and oscillating expression of period and cryptochrome in short and long photoperiods suggest a conserved function in Nasonia vitripennis. Chronobiol. Int. 2014, 31, 749–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Q.; Metterville, D.; Briscoe, A.D.; Reppert, S.M. Insect cryptochromes: Gene duplication and loss define diverse ways to construct insect circadian clocks. Mol. Biol. Evol. 2007, 24, 948–955. [Google Scholar] [CrossRef] [Green Version]
- Hanniger, S.; Dumas, P.; Schofl, G.; Gebauer-Jung, S.; Vogel, H.; Unbehend, M.; Heckel, D.G.; Groot, A.T. Genetic basis of allochronic differentiation in the fall armyworm. BMC Evol. Biol. 2017, 17, 68. [Google Scholar] [CrossRef] [Green Version]
- Suszczynska, A.; Kaniewska, M.M.; Bebas, P.; Giebultowicz, J.M.; Kotwica-Rolinska, J. Circadian regulation of caterpillar feeding and growth. J. Insect Physiol. 2017, 101, 113–122. [Google Scholar] [CrossRef]
- Lugena, A.B.; Zhang, Y.; Menet, J.S.; Merlin, C. Genome-wide discovery of the daily transcriptome, DNA regulatory elements and transcription factor occupancy in the monarch butterfly brain. PLoS Genet. 2019, 15, e1008265. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, S.; Shen, Z.J.; Liu, Y.; Li, Z.; Liu, X. Vrille is required for larval moulting and metamorphosis of Helicoverpa armigera (Lepidoptera: Noctuidae). Insect Mol. Biol. 2019, 28, 355–371. [Google Scholar] [CrossRef]
- Beaver, L.M.; Hooven, L.A.; Butcher, S.M.; Krishnan, N.; Sherman, K.A.; Chow, E.S.Y.; Giebultowicz, J.M. Circadian Clock Regulates Response to Pesticides in Drosophila via Conserved Pdp1 Pathway. Toxicol. Sci. 2010, 115, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Groot, A.T. Circadian rhythms of sexual activities in moths: A review. Front. Ecol. Evol. 2014, 2, 43. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, S.; Li, W.; Chen, Z.; Guo, H.; Liu, J.; Xu, Y.; Xiao, Y.; Zhang, L.; Arunkumar, K.P.; et al. Circadian regulation of night feeding and daytime detoxification in a formidable Asian pest Spodoptera litura. Commun. Biol. 2021, 4, 286. [Google Scholar] [CrossRef]
- Tomioka, K.; Matsumoto, A. A comparative view of insect circadian clock systems. Cell. Mol. Life Sci. 2010, 67, 1397–1406. [Google Scholar] [CrossRef] [PubMed]
- Numata, H.; Miyazaki, Y.; Ikeno, T. Common features in diverse insect clocks. Zool. Lett. 2015, 1, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomioka, K.; Uryu, O.; Kamae, Y.; Umezaki, Y.; Yoshii, T. Peripheral circadian rhythms and their regulatory mechanism in insects and some other arthropods: A review. J. Comp. Physiol. B 2012, 182, 729–740. [Google Scholar] [CrossRef]
- Selcho, M.; Millan, C.; Palacios-Munoz, A.; Ruf, F.; Ubillo, L.; Chen, J.T.; Bergmann, G.; Ito, C.; Silva, V.; Wegener, C.; et al. Central and peripheral clocks are coupled by a neuropeptide pathway in Drosophila. Nat. Commun. 2017, 8, 15563. [Google Scholar] [CrossRef]
- Xu, K.Y.; Zheng, X.Z.; Sehgal, A. Regulation of Feeding and Metabolism by Neuronal and Peripheral Clocks in Drosophila. Cell Metab. 2008, 8, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Sauman, I.; Reppert, S.M. Brain control of embryonic circadian rhythms in the silkmoth Antheraea pernyi. Neuron 1998, 20, 741–748. [Google Scholar] [CrossRef] [Green Version]
- Hege, D.M.; Stanewsky, R.; Hall, J.C.; Giebultowicz, J.M. Rhythmic expression of a PER-reporter in the malpighian tubules of decapitated Drosophila: Evidence for a brain-independent circadian clock. J. Biol. Rhythm. 1997, 12, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Giebultowicz, J.M.; Stanewsky, R.; Hall, J.C.; Hege, D.M. Transplanted Drosophila excretory tubules maintain circadian clock cycling out of phase with the host. Curr. Biol. 2000, 10, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Ito, C.; Goto, S.G.; Tomioka, K.; Numata, H. Temperature Entrainment of the Circadian Cuticle Deposition Rhythm in Drosophila melanogaster. J. Biol. Rhythm. 2011, 26, 14–23. [Google Scholar] [CrossRef] [Green Version]
- Tanoue, S.; Krishnan, P.; Krishnan, B.; Dryer, S.E.; Hardin, P.E. Circadian clocks in antennal neurons are necessary and sufficient for olfaction rhythms in Drosophila. Curr. Biol. 2004, 14, 638–649. [Google Scholar] [CrossRef] [Green Version]
- Beaver, L.M.; Gvakharia, B.O.; Vollintine, T.S.; Hege, D.M.; Stanewsky, R.; Giebultowicz, J.M. Loss of circadian clock function decreases reproductive fitness in males of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2002, 99, 2134–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanchenko, M.; Stanewsky, R.; Giebultowicz, J.M. Circadian photoreception in Drosophila: Functions of cryptochrome in peripheral and central clocks. J. Biol. Rhythm. 2001, 16, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Bebas, P.; Cymborowski, B.; Giebultowicz, J.M. Circadian rhythm of sperm release in males of the cotton leafworm, Spodoptera littoralis: In vivo and in vitro studies. J. Insect Physiol. 2001, 47, 859–866. [Google Scholar] [CrossRef]
- Kotwica, J.; Bebas, P.; Gvakharia, B.O.; Giebultowicz, J.M. RNA Interference of the Period Gene Affects the Rhythm of Sperm Release in Moths. J. Biol. Rhythm. 2009, 24, 25–34. [Google Scholar] [CrossRef]
- Polanska, M.A.; Maksimiuk-Ramirez, E.; Ciuk, M.A.; Kotwica, J.; Bebas, P. Clock-controlled rhythm of ecdysteroid levels in the haemolymph and testes, and its relation to sperm release in the Egyptian cotton leafworm, Spodoptera littoralis. J. Insect Physiol. 2009, 55, 426–434. [Google Scholar] [CrossRef]
- Kotwica, J.; Ciuk, M.A.; Joachimiak, E.; Rowinski, S.; Cymborowski, B.; Bebas, P. Carbonic Anhydrase Activity in the Vas Deferens of the Cotton Leafworm—Spodoptera Littoralis (Lepidoptera: Noctuidae) Controlled by Circadian Clock. J. Physiol. Pharmacol. 2006, 57, 107–123. [Google Scholar]
- Kotwica-Rolinska, J.; Gvakharia, B.O.; Kedzierska, U.; Giebultowicz, J.M.; Bebas, P. Effects of period RNAi on V-ATPase expression and rhythmic pH changes in the vas deferens of Spodoptera littoralis (Lepidoptera: Noctuidae). Insect Biochem. Mol. Biol. 2013, 43, 522–532. [Google Scholar] [CrossRef]
- Lim, F.L.; Currie, R.A.; Orphanides, G.; Moggs, J.G. Emerging evidence for the interrelationship of xenobiotic exposure and circadian rhythms: A review. Xenobiotica 2006, 36, 1140–1151. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, K.; Zhao, Y.; Fent, K. Environmental chemicals affect circadian rhythms: An underexplored effect influencing health and fitness in animals and humans. Environ. Int. 2021, 149, 106159. [Google Scholar] [CrossRef]
- Schibler, U. The daily timing of gene expression and physiology in mammals. Dialogues Clin. Neurosci. 2007, 9, 257–272. [Google Scholar] [CrossRef]
- Claudel, T.; Cretenet, G.; Saumet, A.; Gachon, F. Crosstalk between xenobiotics metabolism and circadian clock. FEBS Lett. 2007, 581, 3626–3633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmona-Antonanzas, G.; Santi, M.; Migaud, H.; Vera, L.M. Light- and clock-control of genes involved in detoxification. Chronobiol. Int. 2017, 34, 1026–1041. [Google Scholar] [CrossRef] [PubMed]
- Prokkola, J.M.; Nikinmaa, M. Circadian rhythms and environmental disturbances—Underexplored interactions. J. Exp. Biol. 2018, 221, jeb179267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.L.; Yue, Z.; Arnold, D.M.; Artiushin, G.; Sehgal, A. A Circadian Clock in the Blood-Brain Barrier Regulates Xenobiotic Efflux. Cell 2018, 173, 130–139. [Google Scholar] [CrossRef] [Green Version]
- Hooven, L.A.; Sherman, K.A.; Butcher, S.; Giebultowicz, J.M. Does the Clock Make the Poison? Circadian Variation in Response to Pesticides. PLoS ONE 2009, 4, e6469. [Google Scholar] [CrossRef]
- Yang, Y.Y.; Liu, Y.; Teng, H.J.; Sauman, I.; Sehnal, F.; Lee, H.J. Circadian control of permethrin-resistance in the mosquito Aedes aegypti. J. Insect Physiol. 2010, 56, 1219–1223. [Google Scholar] [CrossRef]
- Bebas, P.; Cymborowski, B. Effect of constant light on male sterility in the cotton leafworm Spodoptera littoralis. Physiol. Entomol. 1999, 24, 165–170. [Google Scholar] [CrossRef]
- Pszczolkowski, M.A.; Kuszczak, B. Effect of an ecdysone agonist, RH-5849, on wandering behaviour in Spodoptera littoralis. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1996, 113, 359–367. [Google Scholar] [CrossRef]
- Robertson, J.; Russell, R.M.; Preisler, H.; Savin, N.E. Bioassays with Arthropods, 2nd ed.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2007. [Google Scholar]
- Gvakharia, B.O.; Bebas, P.; Cymborowski, B.; Giebultowicz, J.M. Disruption of sperm release from insect testes by cytochalasin and ß-actin mRNA mediated interference. Cell. Mol. Life Sci. 2003, 60, 1744–1751. [Google Scholar] [CrossRef]
- Aitio, A. A Simple and Sensitive Assay of 7-Ethoxycoumarin Deethylation. Anal. Biochem. 1978, 85, 488–491. [Google Scholar] [CrossRef]
- Heinrichhirsch, B.; Hofmann, D.; Webb, J.; Neubert, D. Activity of Aldrinepoxidase, 7-Ethoxycoumarin-O-Deethylase and 7-Ethoxyresorufin-O-Deethylase during the Development of Chick-Embryos In Ovo. Arch. Toxicol. 1990, 64, 128–134. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, G.; Cuany, A.; Brun, A.; Amichot, M.; Rahmani, R.; Berge, J.B. A microfluorometric method for measuring ethoxycoumarin-O-deethylase activity on individual Drosophila melanogaster abdomens: Interest for screening resistance in insect populations. Anal. Biochem. 1995, 229, 86–91. [Google Scholar] [CrossRef] [PubMed]
- Vidau, C.; Diogon, M.; Aufauvre, J.; Fontbonne, R.; Vigues, B.; Brunet, J.L.; Texier, C.; Biron, D.G.; Blot, N.; El Alaoui, H.; et al. Exposure to sublethal doses of fipronil and thiacloprid highly increases mortality of honeybees previously infected by Nosema ceranae. PLoS ONE 2011, 6, e21550. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wu, Y.; Chen, S.; Devine, G.J.; Denholm, I.; Jewess, P.; Moores, G.D. The involvement of microsomal oxidases in pyrethroid resistance in Helicoverpa armigera from Asia. Insect Biochem. Mol. Biol. 2004, 34, 763–773. [Google Scholar] [CrossRef] [PubMed]
- Su, J.Y.; Lai, T.C.; Li, J. Susceptibility of field populations of Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae) in China to chlorantraniliprole and the activities of detoxification enzymes. Crop. Prot. 2012, 42, 217–222. [Google Scholar] [CrossRef]
- Van Asperen, K. A study of housefly esterases by means of a sensitive colouring method. J. Insect Physiol. 1962, 8, 401–416. [Google Scholar] [CrossRef]
- Gamil, W.E.; Mariy, F.M.; Youssef, L.A.; Halim, S.M.A. Effect of Indoxacarb on some biological and biochemical aspects of Spodoptera littoralis (Boisd.) larvae. Ann. Agric. Sci. 2011, 56, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Ben-Yakir, D. Optical Manipulation of Arthropod Pests and Beneficials, 1st ed.; CAB International: Boston, MA, USA, 2020. [Google Scholar]
- Brady, J. The physiology of insect circadian rhythms. In Advances in Insect Physiology; Treherne, J.E., Treherne, V.E., Berridge, M.J., Wigglesworth, V.B., Eds.; Academic Press: London, UK, 1974; Volume 10, pp. 1–115. [Google Scholar]
- Onyeocha, F.A.; Fuzeaubraesch, S. Circadian-Rhythm Changes in Toxicity of the Insecticide Dieldrin on Larvae of the Migratory Locust Locusta migratoria migratorioides. Chronobiol. Int. 1991, 8, 103–109. [Google Scholar] [CrossRef]
- Piechowicz, B.; Stawarczyk, K.; Stawarczyk, M. Circadian changes in susceptibility of young honeybee workers to intoxication by pyrethroid, carbamate, organophosphorus, benzoyl urea and pyridine derivative insecticides. J. Plant Prot. Res. 2012, 52, 286–289. [Google Scholar] [CrossRef]
- Piechowicz, B.; Grodzicki, P.; Stawarczyk, M.; Stawarczyk, K. Circadian and Seasonal Changes in Honeybee (Apis mellifera) Worker Susceptibility to Diazinon, Teflubenzuron, Pirimicarb, and Indoxacarb. Pol. J. Environ. Stud. 2013, 22, 1457–1463. [Google Scholar]
- Bagheri, F.; Talebi, K.; Hosseininaveh, V.; Allahyari, H.; Habibi-Rezaei, M.; Zare, S. Circadian Rhythmicity of Diazinon Susceptibility, Detoxifying Enzymes, and Energy Reserves in Aphis gossypii (Hemiptera: Aphididae). J. Econ. Entomol. 2016, 109, 1651–1659. [Google Scholar] [CrossRef] [PubMed]
- Egi, Y.; Sakamoto, K. Daily rhythm of insecticide susceptibility in the silkworm Bombyx mori. Biol. Rhythm Res. 2016, 47, 247–252. [Google Scholar] [CrossRef] [Green Version]
- Maliszewska, J.; Piechowicz, B.; Maciaga, G.; Zareba, L.; Marcinkowska, S. Pyrethroid residue dynamics in insects depends on the circadian clock. J. Environ. Sci. Health Part B 2018, 53, 441–446. [Google Scholar] [CrossRef]
- Ptitsyn, A.A.; Reyes-Solis, G.; Saavedra-Rodriguez, K.; Betz, J.; Suchman, E.L.; Carlson, J.O.; Black, W.C. Rhythms and synchronization patterns in gene expression in the Aedes aegypti mosquito. BMC Genom. 2011, 12, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meireles-Filho, A.C.; Kyriacou, C.P. Circadian rhythms in insect disease vectors. Mem. Inst. Oswaldo Cruz 2013, 108, 48–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Severson, D.W.; Behura, S.K. Mosquito Genomics: Progress and Challenges. Annu. Rev. Entomol. 2012, 57, 143–166. [Google Scholar] [CrossRef] [PubMed]
- Rund, S.S.; Hou, T.Y.; Ward, S.M.; Collins, F.H.; Duffield, G.E. Genome-wide profiling of diel and circadian gene expression in the malaria vector Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2011, 108, E421–E430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balmert, N.J.; Rund, S.S.; Ghazi, J.P.; Zhou, P.; Duffield, G.E. Time-of-day specific changes in metabolic detoxification and insecticide resistance in the malaria mosquito Anopheles gambiae. J. Insect Physiol. 2014, 64, 30–39. [Google Scholar] [CrossRef]
- Rund, S.S.; Bonar, N.A.; Champion, M.M.; Ghazi, J.P.; Houk, C.M.; Leming, M.T.; Syed, Z.; Duffield, G.E. Daily rhythms in antennal protein and olfactory sensitivity in the malaria mosquito Anopheles gambiae. Sci. Rep. 2013, 3, 2494. [Google Scholar] [CrossRef] [Green Version]
- Hamby, K.A.; Kwok, R.S.; Zalom, F.G.; Chiu, J.C. Integrating Circadian Activity and Gene Expression Profiles to Predict Chronotoxicity of Drosophila suzukii Response to Insecticides. PLoS ONE 2013, 8, e68472. [Google Scholar] [CrossRef] [Green Version]
- Khalid, M.F.; Lee, C.-Y.; Doggett, S.L.; Singham, G.V. Circadian rhythms in insecticide susceptibility, metabolic enzyme activity, and gene expression in Cimex lectularius (Hemiptera: Cimicidae). PLoS ONE 2019, 14, e0218343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela, G.M.; Stroppa, M.M.; Garcia, B.A. Daily Variations in the Expression of Genes Related to Insecticide Resistance in the Chagas Disease Vector Triatoma infestans (Hemiptera: Reduviidae). Am. J. Trop. Med. Hyg. 2019, 100, 1482–1485. [Google Scholar] [CrossRef] [PubMed]
- Subala, S.P.R.R.; Shivakumar, M.S. Changes in light and dark periods affect the arylalkylamine N-acetyl transferase, melatonin activities and redox status in the head and hemolymph of nocturnal insect Spodoptera litura. Biol. Rhythm Res. 2018, 49, 13–28. [Google Scholar] [CrossRef]
- Villanueva, O.K.; Ponce, G.; Lopez, B.; Gutierrez, S.M.; Rodriguez, I.P.; Reyes, G.; Saavedra, K.J.; Black, W.C.; Garcia, J.; Beaty, B.; et al. Effect of Photoperiod on Permethrin Resistance in Aedes Aegypti. J. Am. Mosq. Control Assoc. 2016, 32, 308–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kola, V.S.R.; Renuka, P.; Madhav, M.S.; Mangrauthia, S.K. Key enzymes and proteins of crop insects as candidate for RNAi based gene silencing. Front. Physiol. 2015, 6, 119. [Google Scholar] [CrossRef] [Green Version]
- Perkin, L.C.; Adrianos, S.L.; Oppert, B. Gene Disruption Technologies Have the Potential to Transform Stored Product Insect Pest Control. Insects 2016, 7, 46. [Google Scholar] [CrossRef] [Green Version]
- Niu, J.Z.; Taning, C.N.T.; Christiaens, O.; Smagghe, G.; Wang, J.J. Rethink RNAi in Insect Pest Control: Challenges and Perspectives. Adv. Insect Physiol. 2018, 55, 1–17. [Google Scholar]
- Kim, Y.H.; Issa, M.S.; Cooper, A.M.W.; Zhu, K.Y. RNA interference: Applications and advances in insect toxicology and insect pest management. Pestic. Biochem. Physiol. 2015, 120, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Chung, S.H.; Feng, H.L.; Jander, G. Engineering pest tolerance through plant-mediated RNA interference. Curr. Opin. Plant Biol. 2021, 60, 102029. [Google Scholar] [CrossRef]
- Dias, N.P.; Cagliari, D.; dos Santos, E.A.; Smagghe, G.; Jurat-Fuentes, J.L.; Mishra, S.; Nava, D.E.; Zotti, M.J. Insecticidal Gene Silencing by RNAi in the Neotropical Region. Neotrop. Entomol. 2020, 49, 1–11. [Google Scholar] [CrossRef]
- Amezian, D.; Nauen, R.; Le Goff, G. Comparative analysis of the detoxification gene inventory of four major Spodoptera pest species in response to xenobiotics. Insect Biochem. Mol. Biol. 2021, 138, 103646. [Google Scholar] [CrossRef] [PubMed]
- Amezian, D.; Nauen, R.; Le Goff, G. Transcriptional regulation of xenobiotic detoxification genes in insects—An overview. Pestic. Biochem. Physiol. 2021, 174, 104822. [Google Scholar] [CrossRef] [PubMed]
- Hilliou, F.; Chertemps, T.; Maibeche, M.; Le Goff, G. Resistance in the Genus Spodoptera: Key Insect Detoxification Genes. Insects 2021, 12, 544. [Google Scholar] [CrossRef] [PubMed]
- Dalla Bona, A.C.; Chitolina, R.F.; Fermino, M.L.; Poncio, L.D.; Weiss, A.; Lima, J.B.P.; Paldi, N.; Bernardes, E.S.; Henen, J.; Maori, E. Larval application of sodium channel homologous dsRNA restores pyrethroid insecticide susceptibility in a resistant adult mosquito population. Parasites Vectors 2016, 9, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Li, T.; Feng, X.C.; Li, M.; Liu, S.K.; Liu, N.N. Multiple cytochrome P450 genes: Conferring high levels of permethrin resistance in mosquitoes, Culex quinquefasciatus. Sci. Rep. 2021, 11, 9041. [Google Scholar] [CrossRef] [PubMed]
- Killiny, N.; Hajeri, S.; Tiwari, S.; Gowda, S.; Stelinski, L.L. Double-Stranded RNA Uptake through Topical Application, Mediates Silencing of Five CYP4 Genes and Suppresses Insecticide Resistance in Diaphorina citri. PLoS ONE 2014, 9, e110536. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Wei, X.G.; Yang, J.; Du, T.H.; Yin, C.; Fu, B.L.; Huang, M.J.; Liang, J.J.; Gong, P.P.; Liu, S.N.; et al. Epitranscriptomic regulation of insecticide resistance. Sci. Adv. 2021, 7, eabe5903. [Google Scholar] [CrossRef]
- Palli, S.R. CncC/Maf-mediated xenobiotic response pathway in insects. Arch. Insect Biochem. Physiol. 2020, 104, e21674. [Google Scholar] [CrossRef]
- Fletcher, S.J.; Reeves, P.T.; Hoang, B.T.; Mitter, N. A Perspective on RNAi-Based Biopesticides. Front. Plant Sci. 2020, 11, 51. [Google Scholar] [CrossRef] [Green Version]
- Szekacs, A.; Ammour, A.S.; Mendelsohn, M.L. Editorial: RNAi Based Pesticides. Front. Plant Sci. 2021, 12, 714116. [Google Scholar] [CrossRef]
- Zhao, M.J.; Zhao, H.; Deng, J.M.; Guo, L.X.; Wu, B.J. Role of the CLOCK protein in liver detoxification. Brit. J. Pharmacol. 2019, 176, 4639–4652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gachon, F.; Firsov, D. The role of circadian timing system on drug metabolism and detoxification. Expert Opin. Drug Metab. Toxicol. 2011, 7, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Gachon, F. Physiological function of PARbZip circadian clock-controlled transcription factors. Ann. Med. 2007, 39, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Kotwica, J.; Larson, M.K.; Bebas, P.; Giebultowicz, J.M. Developmental profiles of PERIOD and DOUBLETIME in Drosophila melanogaster ovary. J. Insect Physiol. 2009, 55, 419–425. [Google Scholar] [CrossRef]
- Nelson, W.; Tong, Y.L.; Lee, J.K.; Halberg, F. Methods for cosinor-rhythmometry. Chronobiologia 1979, 6, 305–323. [Google Scholar]
- Refinetti, R.; Cornelissen, G.; Halberg, F. Procedures for numerical analysis of circadian rhythms. Biol. Rhythm Res. 2007, 38, 275–325. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haj Darwich, C.M.; Chrzanowski, M.M.; Bernatowicz, P.P.; Polanska, M.A.; Joachimiak, E.; Bebas, P. Molecular Oscillator Affects Susceptibility of Caterpillars to Insecticides: Studies on the Egyptian Cotton Leaf Worm—Spodoptera littoralis (Lepidoptera: Noctuidae). Insects 2022, 13, 488. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050488
Haj Darwich CM, Chrzanowski MM, Bernatowicz PP, Polanska MA, Joachimiak E, Bebas P. Molecular Oscillator Affects Susceptibility of Caterpillars to Insecticides: Studies on the Egyptian Cotton Leaf Worm—Spodoptera littoralis (Lepidoptera: Noctuidae). Insects. 2022; 13(5):488. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050488
Chicago/Turabian StyleHaj Darwich, Choukri M., Marcin M. Chrzanowski, Piotr P. Bernatowicz, Marta A. Polanska, Ewa Joachimiak, and Piotr Bebas. 2022. "Molecular Oscillator Affects Susceptibility of Caterpillars to Insecticides: Studies on the Egyptian Cotton Leaf Worm—Spodoptera littoralis (Lepidoptera: Noctuidae)" Insects 13, no. 5: 488. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13050488