
Diverse Host Plants of the First Instars of the Invasive Lycorma delicatula: Insights from eDNA Metabarcoding


Department of Entomology, University of Maryland, College Park, MD 20742, USA
*
Author to whom correspondence should be addressed.
Insects 2022, 13(6), 534; https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060534
Submission received: 25 April 2022
/
Revised: 3 June 2022
/
Accepted: 9 June 2022
/
Published: 10 June 2022
(This article belongs to the Special Issue Advances on Invasive Insect Pests: Insect Behavior, Host Plant Usage, Biocontrol, and More)
Abstract
:Simple Summary
The globalization of human activities, especially in agriculture, has facilitated the range expansion of insect pests, promoting species invasions in new territories. Here, we focused on the invasive spotted lanternfly, Lycorma delicatula, an important economic pest, accidentally introduced in the United States and first detected in 2014. Our study investigated host-plant usage by the first nymphal instars, which are more challenging to find and monitor than other stages. Using DNA metabarcoding of nymphal gut contents (i.e., detection of DNA from multiple ingested plants) in this study we determined and characterized the ingested plants that could be included in a broad host-plant range of early nymphal stages of L. delicatula. This, in turn, will have important applications for developing effective management programs to control the invasive spotted lanternfly. The results from our study will be of great interest for all the forest managers and growers in the potential (national) range of the spotted lanternfly, and will help them ultimately minimize their efforts and expenses needed for managing this important pest.
Abstract
Identification of host plants of the invasive spotted lanternfly, Lycorma delicatula (Hemiptera: Fulgoridae), has been the focus of many studies. While the adults and late nymphs are relatively easy to observe on plants and to use for molecular gut-content analysis, studying the early instars is more challenging. This study is the continuation of our ongoing efforts to determine the host range for each developmental stage of L. delicatula. In the present study, we focused exclusively on the first nymphal instars, and we used a novel approach, utilizing “bulk” DNA extracts for DNA metabarcoding of nymphal gut contents, to identify all the detectable plants that the nymphs had ingested prior to being collected. We were able to obtain high-quality amplicons (up to 406 bp) of a portion of the rbcL gene and detect 27 unique ingested plant species belonging to 17 families. Both native and introduced plants with the prevalence of trees and grasses were present among the ingested plants. We also identified 13 novel host plants that have not been previously reported for L. delicatula on the U.S. territory. The results from our study have important applications for developing effective programs on early monitoring of invasive L. delicatula.
1. Introduction
The spotted lanternfly, Lycorma delicatula (Hemiptera: Fulgoridae), is a highly invasive sap-feeding insect in North America, and one of the most aggressive insect pests in the Mid-Atlantic region of the United States [1,2,3,4,5,6,7]. Since its first discovery in Berks County, Pennsylvania, from Asia in 2014, L. delicatula has quickly dispersed and established its populations in neighboring states and beyond. The invasion of L. delicatula continues on U.S. territory: to date, L. delicatula individuals have been detected in 15 states; of these, 9 states have heavy infestations, and 4 states have multiple internal quarantine zones [8].
The host-plant range of L. delicatula has been the focus of many studies as it is highly polyphagous, and current host lists span a large number of host plants of various taxonomic levels, life forms, morphology, and plant architecture, as well as both coevolved (i.e., originated in Asia) and noncoevolved hosts (i.e., originated in other regions than Asia, both native and introduced to the U.S.), including various forest trees (tree-of-heaven, birch, maple, walnut, oak, etc.), fruit trees (such as apple trees and grapes), as well as many ornamental plants [4,7,9,10,11]. Being highly polyphagous, L. delicatula poses a significant risk to forestry and agriculture; adults and four nymphal instars feed on host phloem tissues, causing plant injury including plant photosynthesis reduction, weeping wounds, and creating conditions for sooty mold that ultimately affects plant growth and reduces the fruit quality [1,3,5,6,12].
Management, and especially early monitoring of L. delicatula, is challenging, particularly due to its unusual use of plant hosts during the insect development; as the nymphs mature, their host plant range decreases, and by the time the insects reach the adult stage they have one or two preferred host plants [1]. To date, there are multiple ongoing efforts to decipher the host-plant range of L. delicatula at both the adult stage and nymphal stages with successful attempts to accurately confirm the consumed host plants by adults and late nymphal instars [9]. Early nymphal instars, however, and especially the first instar, received less attention in the experimental studies. Due to their small size, cryptic coloration, and a discreet sap-feeding behavior, the first instars are challenging to find and monitor. While most studies on L. delicatula feeding preferences still focus on the experimental observations [5,13], recent advances using PCR-based DNA analyses provide opportunities for rapid and reliable detection and identification of L. delicatula’s diet [6,14]. In our previous work on plant DNA detection from the gut contents of L. delicatula [6], we focused on late nymphal instars only (third and fourth nymphal instars). Using Sanger sequencing to detect the most abundant plant item in the lanternfly gut contents, we found that (a) ingested plants in ~93% of the nymphs did not correspond with the plants from which the nymphs were collected (possibly due to high mobility of nymphs), and (b) both coevolved (i.e., introduced into the U.S.) and noncoevolved (i.e., native to the U.S.) plants, as well as woody and nonwoody plants, were ingested. Additionally, to optimize our DNA work, we conducted a systematic review of published PCR approaches to detect the host plants ingested by insect pests [15].
DNA metabarcoding of insect gut contents, in particular, has been demonstrated as an effective approach that allows us to accurately determine a host-plant range of insect pests [16], to reconstruct the invasion route of insect pests [17], insect pest migration [18,19,20], as well as to record realized and novel plant–insect associations [21,22,23]. In general, the advantage of DNA metabarcoding over observation or morphology-based approaches to insect diet analysis is detection of ingested plant diversity and composition [24]. The DNA metabarcoding approach has important applications for biomonitoring of species in diverse communities [25], and is particularly applied to restoration monitoring [24]. Additionally, recent practices in environmental DNA (eDNA) barcoding (i.e., simultaneous identification of species from multiple taxa within one environmental sample, using a specific fragment of DNA) demonstrated the advantage of using environmental bulk DNA extracts over DNA barcoding of individual organisms [26,27,28]. Following these recent advances, in this study, for the first time, we apply a “bulk” extraction of DNA from multiple nymphal individuals of L. delicatula. We expected that this approach (a) would be beneficial for increasing DNA yield in each bulk sample (compared to a “single-insect” approach); (b) would be time- and cost-efficient; and most importantly, (c) would result in diverse gut-content samples which, in turn, would accurately represent the diversity of ingested plant species, which is the primary focus of our study. Finally, following suggested applications in Van Der Heyde et al. [24], we aimed to show the utility of the DNA metabarcoding of gut contents of L. delicatula in early monitoring of the first nymphal instars, for which there is a growing need. Such a DNA metabarcoding approach is necessary for effective monitoring of movement of L. delicatula populations on their host plants, as well as early detection of novel host plants and potential host switch.
To address these goals, the main focus of this study was to demonstrate the utility of eDNA metabarcoding for determining a host-plant range for the first nymphal instar of the invasive spotted lanternfly, L. delicatula. Additionally, we explore the diversity of ingested plants in terms of woodiness and perenniality, as well as the proportions of coevolved/noncoevolved host plants. We also expected to detect the ingested plant species that had not been previously recorded as host plants of L. delicatula. Based on these findings, we discussed important implications for potential host plants of early nymphal instars and their effective monitoring. Since we focused on the ingested plants only, similar to our previous research on diet analysis of insect pests [6,29], and for the purpose of this study only, we will continue using the term “host plants” to indicate consumption of insect food plants only, i.e., the plants that L. delicatula nymphs utilize as suitable food source, and not especially suitable as sources for reproduction and development.
2. Materials and Methods
2.1. Study Site
The plants and the first nymphal instars were collected in Cecil County (MD, USA) in May 2021 during two collection trips, a week apart. The collection site (30 × 73 m2) was a forested area adjacent to a grass field and was located in the Fair Hill State Natural Resource Management Area (FH-NRMA) (39°42′36.3′′ N, 75°51′02.98′′ W, Elkton, MD, USA) (Figure 1a). The weather conditions at the site, on both collection days, ranged from no clouds to partial coverage with a temperature between 26–29 °C and calm-to-light wind. This site was chosen due to (a) the known established population of L. delicatula in this area; (b) a high number of observed egg masses on tree trunks; and (c) its high plant diversity (especially in woody plants). The FH-NRMA is located in Fair Hill, northeastern MD, in a 12 ha forested area. Previous studies have indicated that the area contains sites with high tree density (225 trees per ha), relatively high tree-canopy height (27.8 m on average), and is dominated by Fagus grandifolia (American beech), Liriodendron tulipifera (yellow poplar), Acer rubrum (red maple), Betula lenta (sweet birch) and Quercus alba (white oak) [30], all of which were reported as preferred or suitable host plants of L. delicatula [9].
2.2. Sample Collection and Processing
In this study, a total of 37 first nymphal instars of L. delicatula and 28 reference plants (i.e., one clipped portion and one leaf sample per unique plant growing at the collection site) were collected from plants along the margin between the grass field and the forested area (Figure 1b–d). That was also the only location the nymphs were observed; all the nymphs were observed on woody plants facing south. The plant samples were collected from the collection site only (and not from the entire Fair Hill State Natural Resource Management Area), i.e., from the site where the nymphs were observed and collected. Of the 28 plant samples collected, L. delicatula nymphs were only observed and collected from four plants: Rosa sp., Rubus phoenicolasius, Ailanthus altissima, and Celastrus orbiculatus (Table 1; samples p001, p002, p005, and p007). Sequence analysis of DNA extracted from the collected plants showed 19 unique plant species. After collection, nymphs and leaf samples were immediately dry-frozen at the site and transported to our laboratory at the University of Maryland where they remained stored at −20 °C until DNA extraction. The clipped plant samples were used to create herbarium samples to aid morphological plant identification when necessary.
2.3. DNA Extraction, PCR Amplification, and Purification
For this study, we utilized our previously developed approach for identification of ingested plants from the gut contents of L. delicatula [6], the potato leafhopper Empoasca fabae [29], and earlier Melanoplus grasshoppers [31,32]. The entire body of L. delicatula nymph and 5–10 mm sized cuts of plant tissue from the leaf samples were used for DNA extraction. To prepare bulk samples, anywhere from six to ten nymphs at a time (depending on insect size) were placed in a 1.5 mL microcentrifuge tube (Fisher Scientific Co., Pittsburg, PA, USA) and their bodies were ground up using RNase-free disposable pellet pestle (Fisher Scientific Co., Pittsburg, PA, USA). The resulting tissue mixture was then evenly separated into other 1.5 mL microcentrifuge tubes, approximately 100 mL into each, to satisfy the manufacturer protocol for DNA extraction. Genomic DNA of the L. delicatula nymph bulk samples was extracted using the DNeasy Blood and Tissue kit (catalog no. 69506, Qiagen Inc., Germantown, MD, USA) following the manufacturer’s protocol. Once DNA was isolated, the samples were stored at −4 °C until PCR amplification. In addition, DNA from one individual first nymphal instar was also extracted; this was used as a reference sample.
On the next step of sample processing, genomic DNA extracts from both individual sample and bulk samples were used to detect a portion (~530 bp) of a coding region of the chloroplast DNA, rbcL gene (ribulose-1,5-biphosphate carboxylase-oxygenase). Primers rbcLaF and rbcLaR (purchased from Integrated DNA Technologies, Inc., San Diego, CA, USA) were used to amplify the targeted DNA region. PCR was run following the protocol described in Avanesyan and Lamp [6] using PCR components and conditions indicated in Table 2. PCR products were then purified using Exo-SAP-IT (catalog no. 78201.1.ML, Affymetrix Inc., Santa Clara, CA, USA) according to manufacturer’s protocol. Leaf samples were processed following Avanesyan et al. [29]; same portion of the rbcL gene was amplified from each DNA extract, and purified, followed by Sanger sequencing which was conducted at Azenta/GENEWIZ (Azenta US, Inc., South Plainfield, NJ, USA).
2.4. Sequence Analysis
The obtained PCR products from insect samples containing amplified plant regions were used for sequencing. Next-generation sequencing (“Amplicon-EZ” service), followed by unique sequence abundance analysis, were conducted at Azenta/GENEWIZ (Azenta US, Inc., South Plainfield, NJ, USA). Amplicon-EZ service provides sequencing and analysis of heterogeneous PCR products. This service processes amplicons up to 500 bp and produces 50,000+ reads per sample. This service was chosen due to (a) full coverage the amplicons of the portion of the rbcL-gene, which was utilized for DNA barcoding; and (b) interactive analysis report of detected unique sequences that can be conveniently used for subsequent species identification. Obtained raw reads were trimmed and merged fasta-files were generated at Azenta using an in-house script; forward and reverse primers (described above) were then used to generate the final unique consensus sequences for any DNA region nested by the pair of these primers. Analysis of sequence quality was conducted on raw reads, for each bulk sample, using FastQC tool in Galaxy platform [33].
Plant species identity for both insect samples (i.e., ingested plants) and plant samples (i.e., reference plants) was determined using BLAST engine in the National Center for Biotechnology Information (NCBI) GenBank database (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/genbank/, accessed on 7 November 2021–3 April 2022). The unique ingested plant species were then determined across all the bulk samples of L. delicatula nymphs. For leaf samples, one forward and one reverse sequence were obtained from Azenta; the sequences were then trimmed using 4Peaks v. 1.7 and aligned using Unipro UGENE platform. Each consensus sequence was then used to determine the plant identity using BLAST, the NCBI GenBank database (also accessed on 7 November 2021–3 April 2022).
Sequence reads obtained from the gut contents of L. delicatula nymphs were sorted by sequence length, and all the sequences longer than 100 bp were used for plant identification. Of these, sequences that showed >90% in matches were used for further analysis. This sequence length was chosen based on our preliminary work on sequence identity, during which the sequences shorter than 100 bp demonstrated a low percentage of identity (50–70%) in matches with sequences deposited in the NCBI GenBank database. Plant origin in relation to North America, and specifically to the eastern US (native vs. introduced), as well as plant life form of all identified ingested plant species was determined using the USDA Plant database (https://plants.usda.gov/home, accessed on 7 November 2021–3 April 2022).
2.5. Measurements and Statistical Analysis
The obtained data on the presence of various ingested plant species in the gut contents of L. delicatula nymphs were first synthesized using counts and proportions. For the reference samples and each bulk sample, a list of unique identified ingested plants with corresponding sequences was first compiled (please see Supplementary Material). Next, a combined list of unique ingested plants across all the samples was constructed, and proportions of various plant families, as well as plants with different origin and life form were calculated. Additionally, for the bulk samples, a proportion of each unique plant species was determined. The prevalence of plant species of different origin, family, as well as various life forms, was analyzed using an exact binomial test. For the purpose of this study, the null hypothesis used for the binomial test was that the types of plants (i.e., woody vs. nonwoody, etc.) were represented in the gut contents of L. delicatula nymphs in equal proportions.
Mean quality scores for each group of forward and reverse-sequence reads were retrieved using FastQC tool in Galaxy platform; only the scores for sequences longer than 100 bp (for the bulk samples) and 150 bp (for the reference samples) were included in data analysis. Mean quality scores were then compared among all the samples using a one-way ANOVA with a post hoc TukeyHSD. The Shapiro–Wilk and Bartlett tests were used to investigate the normality and heteroscedasticity of data, respectively. Data analysis, followed by creating pie charts and a boxplot, was conducted in R v.4.1.0 [34].
3. Results
3.1. Basic Sequence Statistics
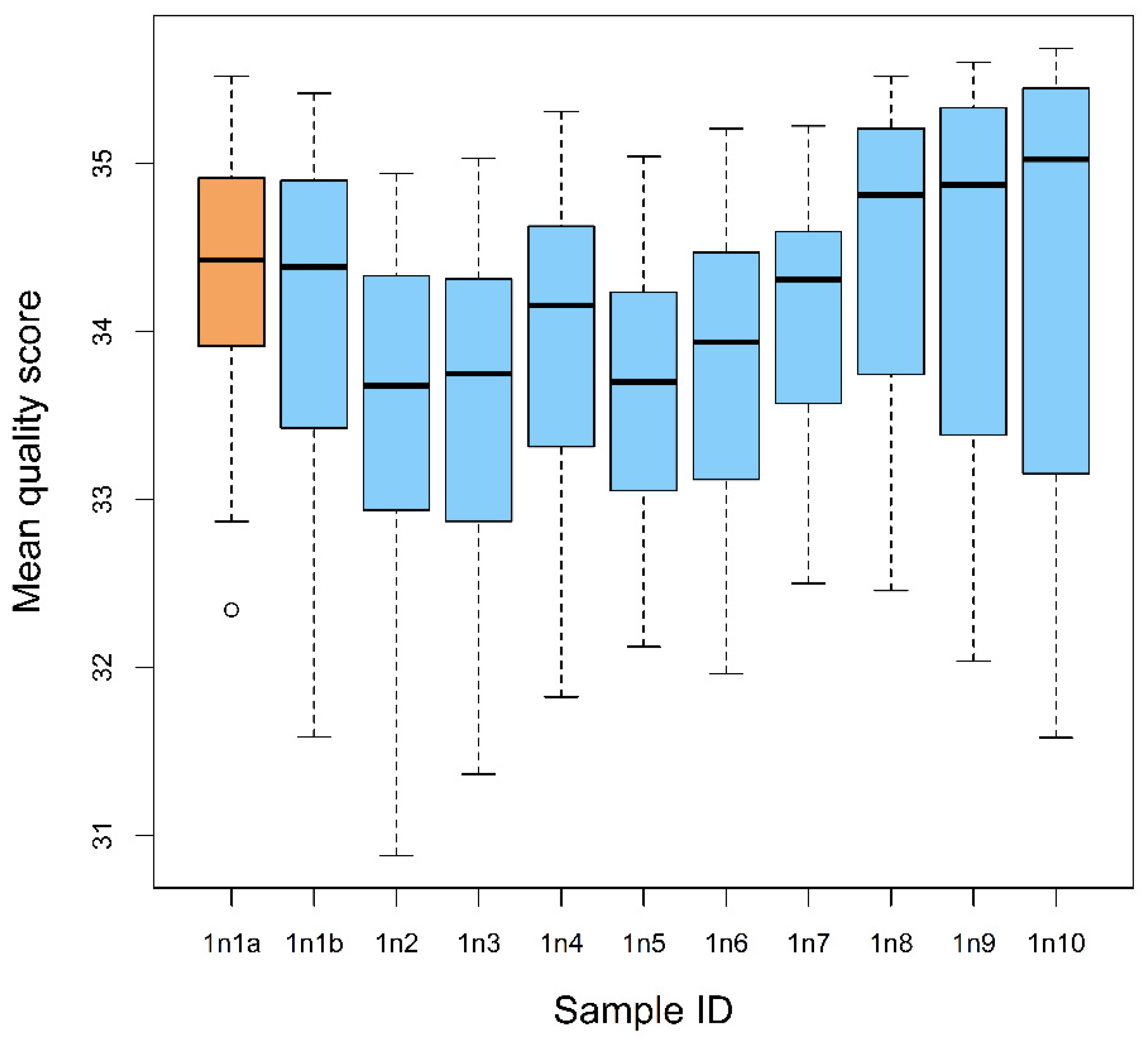
On average, 170,078 ± 9680 total sequence reads were retrieved from each bulk sample (samples 1n1-1n10) and 347,901 reads were obtained from a reference sample (one individual nymph) (Table 3). For the bulk samples, the analysis of consensus sequences revealed 6.9 ± 1.7 readable sequences longer than 100 bp, with the average quality scores >33 (high sequence quality); 65 readable sequences, 150 bp and longer, were obtained from the reference sample (Table 3, Figure 2).
We did not observe any pattern of sequence quality scores across the reference and bulk samples: three bulk samples showed the mean sequence scores that were significantly lower than that in the reference sample (ANOVA: F10,651 = 9.475, p < 0.001; TukeyHSD: p < 0.05), while the differences in the mean quality scores between the reference sample and other bulk samples were not significant. Similarly, pairwise comparisons of the bulk samples (46 pairs total) showed significant difference in the mean quality in 15 pairs, while the scores in the other pairs were not significantly different (Table 3).
3.2. Diversity of Ingested Plants
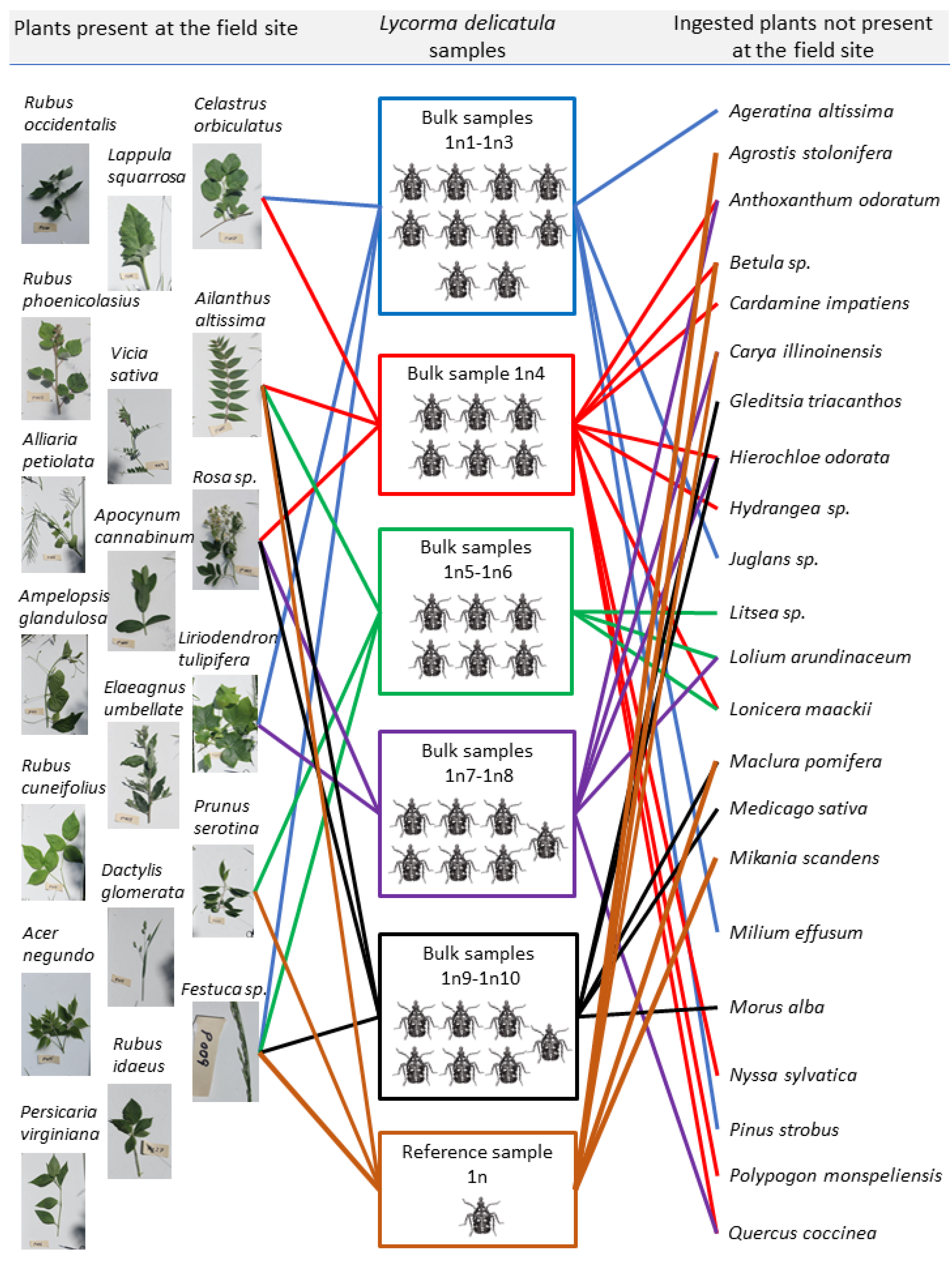
Sequence analysis of DNA extracted from the collected plants showed 19 unique plant species (Figure 3). DNA metabarcoding of gut contents of 37 first nymphal instars of L. delicatula revealed 27 unique ingested plant species; of these, six plant species were present at the collection site, two of which were the plants the nymphs were collected from (Table 4, Figure 3). From one to 12 unique plant species were identified from the bulk samples (samples 1n1-1n10; 4 ± 1.01 plant species per sample), and 11 unique plant species were identified from the reference sample, with Ailanthus altissima and Festuca sp. being the most common plants detected across all the samples (Figure 3). Interestingly, sample “1n4” containing a bulk DNA extract from six nymphs yielded the maximum number (12) of the unique ingested plants. Sequences from all the identified plant species demonstrated 99–100% (Mean% ± SE: 99.79 ± 0.06) match with sequences for corresponding plant species deposited in the NCBI GenBank database (Table 4).
Of the 27 unique ingested plants, 22 plants were identified to species and 5 plants to genus, with 2–3 best matches. The latter included the following genera: Betula sp. (with the highest match for Betula lenta, Betula pendula, and Betula papyrifera); Hydrangea sp. (high match with multiple species); Festuca sp. (with the highest match for Festuca brevipila and Festuca ovina); Litsea sp. (high match with multiple species); Prunus sp. (with the highest match for Prunus serotina and Prunus virginiana), and Rosa sp. (with the highest match for Rosa laevigata, Rosa multiflora, and Rosa rugosa). The presence of all of these identified ingested plant species in Cecil County, Maryland, USA was confirmed through the USDA PLANT database (https://plants.usda.gov/java/, accessed on 3 April 2022). Finally, when the list of identified ingested plants was compared with the latest published host plant list for L. delicatula [9], and with the recent findings on ingested plants from L. delicatula gut contents [14], 13 novel species/genera were identified (Table 4).
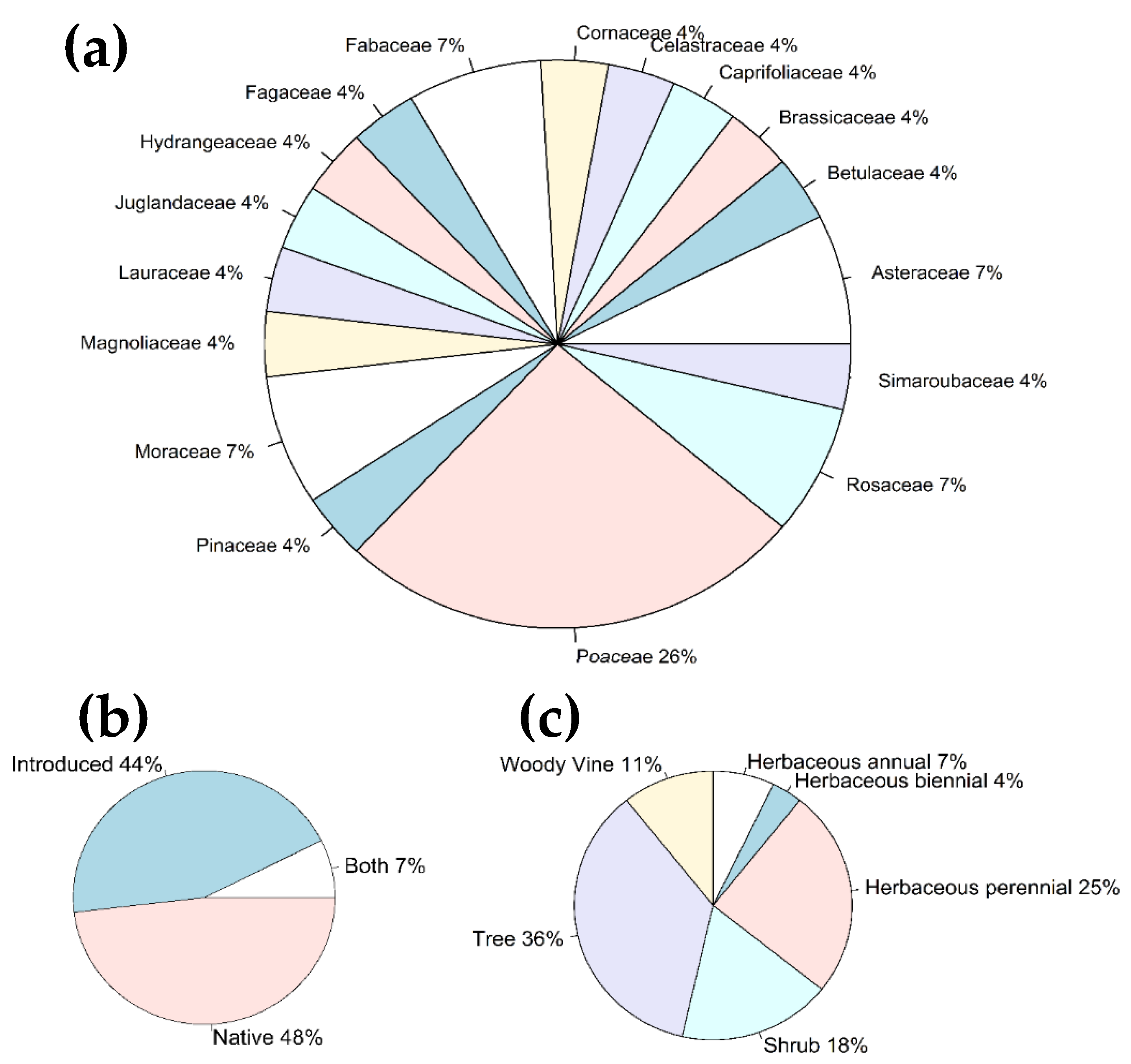
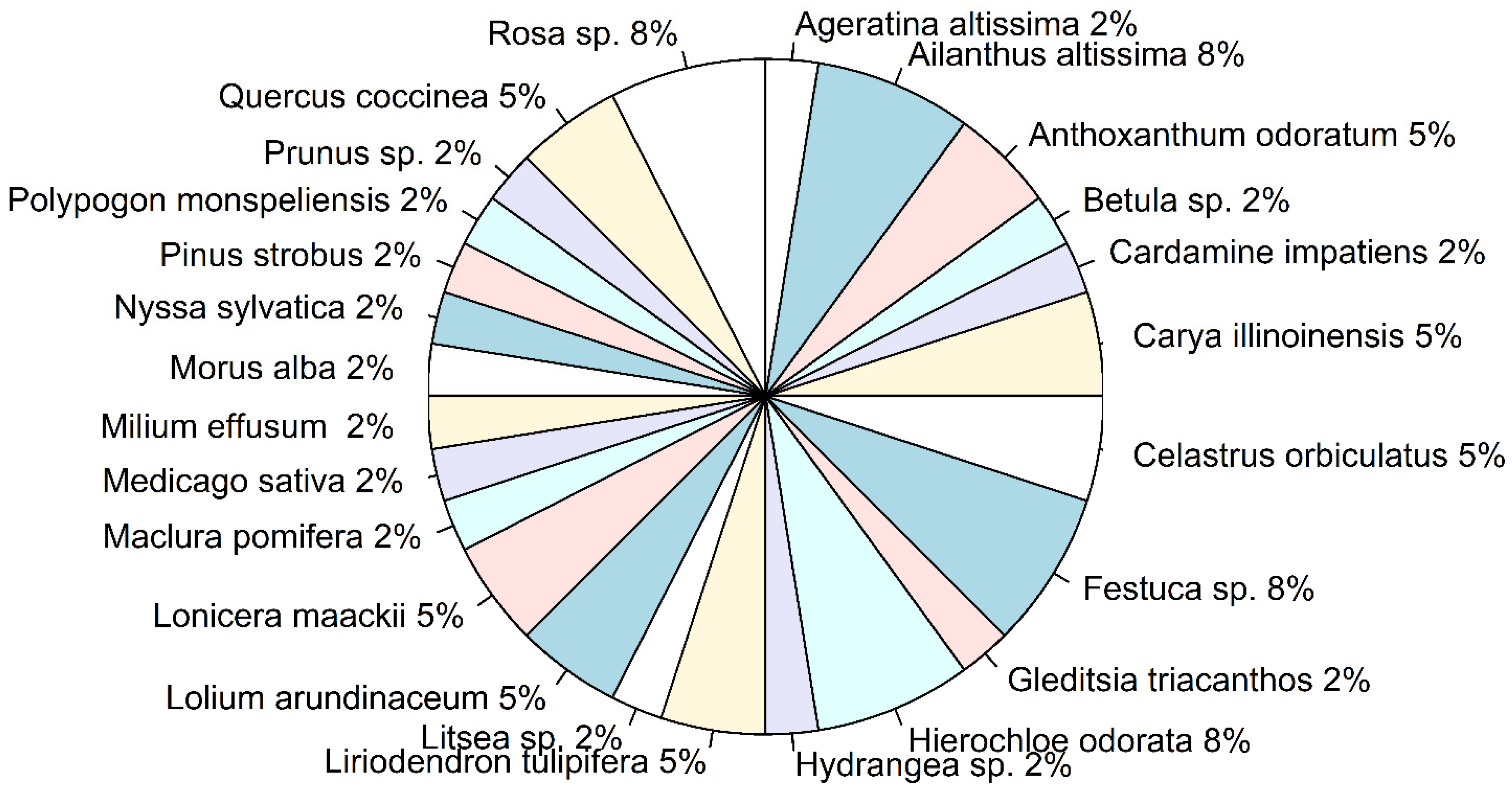
Identified ingested plants, from both the reference and bulk samples, belonged to 17 families (Figure 4a). Both native (48%) and introduced (44%) plants were identified among all the unique ingested plants, with 7% of plants (genera Betula sp. and Hydrangea sp.), which include both native and introduced species (Figure 4b). Trees and herbaceous plants were significantly prevalent among the ingested plants (Binomial test: p < 0.05)(Figure 4c); of all the herbaceous plants, grasses of the Poaceae family were dominant (Binomial test: p < 0.05). When the proportions of ingested plant species were analyzed across the bulk samples only, no significant prevalence of any of the species was observed (Binomial test: p > 0.05) (Figure 5).
4. Discussion
The results from this study demonstrated the successful effective application of eDNA metabarcoding to diet analysis of the first nymphal instars of L. delicatula. Amplicons of a portion of the chloroplast rbcL gene (up to 406bp) were reliably detected and identity of ingested plants was determined. Through the DNA metabarcoding of the gut contents of the fist nymphal instars of L. delicatula, we were able to (a) detect 27 unique ingested plant species belonging to 17 families; (b) demonstrate that up to 12 unique plant species can be retrieved per bulk sample containing 6–10 nymphs; (c) identify 13 novel host plants that have not been previously included in published host plant lists for L. delicatula in the U.S. territory; and (d) show that both native and introduced plant species with the prevalence of trees and grasses were suitable for feeding by the fist nymphal instars of L. delicatula. We discuss potential implications of these results for host interactions, ecology, and early monitoring of L. delicatula below.
4.1. Diversity of Ingested Plants
Our results support previous findings of polyphagous feeding behavior of early instars of L. delicatula [1,14,35]. Particularly, many authors reported the narrowing pattern of L. delicatula host plant use, from the first instars to adults [1,3,4,35,36]. Dechaine et al. [35] in their study observed first instars of L. delicatula on 33 different plant species; this number then decreases to 25 in late instars to 3 in adults. Nixon et al. [37] evaluated the survival of early nymphal instars on 10 host plants and showed significantly higher levels of survivorship on tree of heaven and black walnut. We detected Juglans sp. in one of the bulk samples, and a closely related pecan, Carya illinoinensis, from the same Juglandaceae family, was also commonly present in both the reference sample and bulk samples. Interestingly, even though C. illinoinensis was the best species match when GenBank was used for identification of the obtained sequence, the collection site was not located in the typical range of this species. It is possible that either these plants were ingested by the lanternfly nymphs while foraging at ornamental plantings near the collection site, or plant sequences for the obtained portion of the rbcL-gene are shared among other closely related Carya species.
Our findings also support, once again, the preference for tree of heaven, Ailanthus altissima, which was present at the collection site and was detected in 55% of all the samples. Cooper et al. [14] showed that up to 17 plant families can be detected from a combined 1st–3rd instars; of these, however, only 10 different taxa (belonging to 7 plant families) were reported for the first instars with 2.5 plant taxa per nymphal individual. The authors, however, used only three nymphal individuals and amplified portions of trnF and ITS genes. We demonstrated in our study that using a portion of rbcL gene in combination with bulk DNA extracts resulted in high DNA yield and detection up to 27 unique ingested plants with up to 12 unique taxa per sample. Future studies might further focus on differences in DNA yield from individual vs. bulk insect samples and optimization of eDNA metabarcoding protocols specifically for diet analysis using gut-content environmental samples.
Barringer and Ciafré [9] in their review reported a broad global host range of L. delicatula. We specifically used this published list to determine whether our findings could potentially contribute to and expand the host plant list of L. delicatula. Thirteen novel plant taxa that we reported in this study were also determined by comparison with the findings by Cooper et al. [14], the only study that used DNA metabarcoding of gut contents of the first instars of L. delicatula. We emphasize that these are novel food plants, and whether they support the development and reproduction of L. delicatula could also be a focus for future studies. Our findings of 13 novel plant taxa have especially important applications for early monitoring of L. delicatula; currently, the control programs heavily focus on egg mass surveys [5]. However, monitoring potential food plants near or around the location of egg masses is also critical.
Our findings of many ingested plant species that were not present at the collection site are in agreement with our previous findings during detection of plant DNA from gut contents in late nymphal instars of L. delicatula [6]. We support our previous interpretation of these results by high mobility of nymphs, and possibly by sampling a number of plants before staying on a plant for feeding. Pearson et al. [38] have shown that psyllids use their stylets to sample the parenchyma cells before finding and ingesting phloem sap. DNA from such parenchyma cells, then, can be detected in insect gut contents. However, the feeding choice of the first instars is especially interesting. In the eastern U.S., L. delicatula lays egg masses once a year, typically from September to November or even December [39]. If the egg masses are laid on the tree trunk, they may be laid up to 17 m above the ground [38]. The eggs hatch in May; once hatched from the eggs (if they are on the tree trunk), the first nymphal instars move up along the tree trunk [1]. Due to various physical forces (such as wind) the nymphs fall to the ground; within 1–2 days the nymphs would ascend the tree again [1]. Considering this cyclic behavior on a host tree, the prevalence of grasses in the nymphal gut contents could be explained by insect feeding on grasses, or at least sampling the grasses, during those days while the nymphs are on or close to the ground before they repeat ascending the tree. It would be interesting for future studies to use eDNA metabarcoding and explore plant diversity in soil samples as DNA from seeds, roots, or plant parts can also be picked up during the nymphs probing or feeding.
4.2. Dispersal and Feeding of L. delicatula Nymphs
Dispersal is an important factor facilitating invasion of an introduced insect; thus, understanding the patterns of the insect dispersal is critical for effective management [40]. Flight dispersal patterns have been well-explored in L. delicatula adults. Previous studies showed that the adult can spontaneously fly up to 50 m at an average speed of 4.64 m/s before landing on the tree trunks of any available tree [41,42,43]. Baker et al. [42] suggested that L. delicatula can move until they meet the food sources needed to complete their development. Similarly, an interesting study by Domingue et al. [44] showed that the adults of L. delicatula which flew towards the open field (potentially attracted by the wavelengths detected in ambient light), often changed their direction and flew towards the shade and towards the trees, presumably for feeding.
While all stages of L. delicatula can contribute to its dispersal, the first instars initiate the pest movement in the certain areas; this, in turn, contributes to L. delicatula chances to find suitable hosts. Little is known about the dispersal of nymphs, though Keller et al. [39] conducted a mark-release-resight study in contiguous deciduous forest at a temporal level exploring the dispersal distances up to 10 days after the release. The authors observed some of the nymphs staying near the release point, while some of the nymphs moved to a variety of trees, shrubs, and understory plants up to 65 m away by day 10. On average, the nymphs were able to move up to 10 m during a single day. For the first instars, specifically, the authors indicated the median dispersal distance as 3.2 m, and up to 6.2 m 7 days after release. It was also shown that most of the nymphs could be recapture within 10 m in the presence of Ailanthus altissima [45]. It would be interesting for future studies to explore feeding of nymphs during their movement away from egg masses, and to assess the plant diversity, cover, and species richness at certain distances from the primary egg masses, to then compare these findings with the data on ingested plant species retrieved from the nymphal gut contents.
4.3. Potential Limitation
During this study we investigated ingested food plants, so we expected the DNA of ingested plants to be degraded to some extent. In case digestion of certain plants was fully completed, we did not expect to detect the DNA from those plants within the lanternfly gut contents. Thus, based on our previous work with L. delicatula and other sap-feeders, we expected to find detectable plant DNA within a few hours after ingestion. As a result, the host-plant range we aimed to identify reflected the host plants ingested over the past few hours at the collection site and might potentially exclude some of the suitable (previously known or unknown) host plants of L. delicatula. Therefore, the 27 ingested plants identified in this study represented a “screenshot” of the plant species that had been consumed and of which remains were still present in the gut contents of nymphs at the moment of collection. Given that three dozen individuals were processed, and that the DNA barcoding process was consistent across all the samples, we believe that this limitation did not affect the overall accuracy of our findings and the outcomes will be valuable for creating a host-plant range for each developmental stage of L. delicatula. This potential limitation could be addressed in future studies by increasing the number of collected nymphs and variety of collection sites to account for any other potential host plants.
Another potential limitation is a possible presence of pollen grains on the insect body surface, which could be detected from the unsterilized insect body surface. We indeed cleaned the insect bodies with 2% bleach solution in our previous studies [29,32]. However, we recently found that the body-surface contamination with plant DNA in sap-feeding potato leafhoppers was not significant (Avanesyan and Lamp, in prep.). Based on these findings, and due to high mobility of the first nymphal instars we did not expect the insect body surface to be contaminated with significant amount of DNA from pollen. In the previous studies, however, we only used Sanger sequencing, and it is possible that the NGS technology which we used in this study was more sensitive to detect any plant DNA fragments which might be present on the insect body surface. We plan to address this issue in our future DNA barcoding studies by a detailed comparison of concentration of plant DNA obtained from gut-content samples and from the insect body surface.
5. Conclusions
The egg masses of L. delicatula are typically laid on tree trunks and branches, which might serve as the first suitable hosts for newly hatched first nymphal instars. However, multiple plants can be sampled and ingested during the movement of the nymphs around their first host plant. Our study showed that plants from at least 27 different taxa, both native and introduced, as well as plants of various life forms, can be used as food plants for the first nymphal instars at a relatively small collection site. In combination with results of previous studies on the nymphal dispersal, our findings emphasize the importance of early monitoring of L. delicatula nymphs, not only on plants in close proximity to the egg masses, but also within several meters from the “initial” host plants.
Supplementary Materials
Raw sequences and sequence quality score data are deposited to https://github.com/alina42/SLF-1st-instar (accessed on 25 April 2022). This project is licensed under the terms of the MIT license.
Author Contributions
C.M., A.A. and W.O.L. designed the study; W.O.L. conducted insect and plant collections; C.M. and A.A. conducted data analysis; all authors contributed to writing and revising the paper. C.M. conducted DNA extractions, sequence analysis, and plant species identification, and deposited sequences to GenBank. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by Maryland Agricultural Experiment Station Competitive Grant Program MD-ENTM-201189 and by the Hatch Project, MD-ENTM-1802.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data obtained and used during this study are deposited to https://github.com/alina42/SLF-1st-instar (accessed on 25 April 2022). This project is licensed under the terms of the MIT license.
Acknowledgments
The authors thank David Hawthorne (Department of Entomology, University of Maryland) for continuous support with providing lab equipment for DNA work; Alireza Shokoohi for help with insect and plant collections and photography; and Aaron Shurtleff (Maryland Department of Agriculture) for help with locating the insect collection site.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kim, J.G.; Lee, E.H.; Seo, Y.M.; Kim, N.Y. Cyclic behavior of Lycorma delicatula (Insecta: Hemiptera: Fulgoridae) on host plants. J. Insect Behav. 2011, 24, 423–435. [Google Scholar] [CrossRef]
- Barringer, L.E.; Donovall, L.R.; Spichiger, S.-E.; Lynch, D.; Henry, D. The first New World record of Lycorma delicatula (Insecta: Hemiptera: Fulgoridae). Entomol. News 2015, 125, 20–23. [Google Scholar] [CrossRef]
- Dara, S.K.; Barringer, L.; Arthurs, S.P. Lycorma delicatula (Hemiptera: Fulgoridae): A new invasive pest in the United States. J. Integr. Pest Manag. 2015, 6, 20. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.H.; Park, Y.L.; Leskey, T.C. A review of biology and management of Lycorma delicatula (Hemiptera: Fulgoridae), an emerging global invasive species. J. Asia-Pac. Entomol. 2019, 22, 589–596. [Google Scholar] [CrossRef]
- Liu, H.; Hartlieb, R.J. Spatial Distribution of Lycorma delicatula (Hemiptera: Fulgoridae) Egg Masses on Tree-of-Heaven, Black Walnut, and Siberian Elm in North America. J. Econ. Entomol. 2019, 113, 1028–1032. [Google Scholar] [CrossRef]
- Avanesyan, A.; Lamp, W.O. Use of molecular gut content analysis to decipher the range of food plants of the invasive spotted lanternfly, Lycorma delicatula. Insects 2020, 11, 215. [Google Scholar] [CrossRef] [Green Version]
- Wakie, T.T.; Neven, L.G.; Yee, W.L.; Lu, Z. The establishment risk of Lycorma delicatula (Hemiptera: Fulgoridae) in the United States and globally. J. Econ. Entomol. 2020, 113, 306–314. [Google Scholar] [CrossRef] [PubMed]
- New York State Integrated Pest Management: Spotted Lanternfly. Available online: https://nysipm.cornell.edu/environment/invasive-species-exotic-pests/spotted-lanternfly/ (accessed on 7 April 2022).
- Barringer, L.; Ciafré, C.M. Worldwide feeding host plants of spotted lanternfly, with significant additions from North America. Environ. Entomol. 2020, 49, 999–1011. [Google Scholar] [CrossRef]
- Uyi, O.; Keller, J.A.; Swackhamer, E.; Hoover, K. Performance and Host Association of Spotted Lanternfly (Lycorma Delicatula) among Common Woody Ornamentals. Sci. Rep. 2021, 11, 15774. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.M.; Calvin, D.; Hills-Stevenson, J. Early Response (2018–2020) to the Threat of Spotted Lanternfly, Lycorma Delicatula (Hemiptera: Fulgoridae) in Pennsylvania. Ann. Entomol. Soc. Am. 2021, 114, 709–718. [Google Scholar] [CrossRef]
- Kim, H.; Kim, S.; Lee, Y.; Lee, H.S.; Lee, S.J.; Lee, J.H. Tracing the Origin of Korean Invasive Populations of the Spotted Lanternfly, Lycorma delicatula (Hemiptera: Fulgoridae). Insects 2021, 12, 539. [Google Scholar] [CrossRef]
- Liu, H. Oviposition substrate selection, egg mass characteristics, host preference, and life history of the spotted lanternfly (Hemiptera: Fulgoridae) in North America. Environ. Entomol. 2019, 48, 1452–1468. [Google Scholar] [CrossRef]
- Cooper, W.R.; Marshall, A.T.; Foutz, J.; Wildung, M.R.; Northfield, T.D.; Crowder, D.W.; Leach, H.; Leskey, T.C.; Halbert, S.E.; Snyder, J.B. Directed Sequencing of Plant Specific DNA Identifies the Dietary History of Four Species of Auchenorrhyncha (Hemiptera). Ann. Entomol. Soc. Am. 2021, 115, 275–284. [Google Scholar] [CrossRef]
- Avanesyan, A.; Sutton, H.; Lamp, W.O. Choosing an effective PCR-based approach for diet analysis of insect herbivores: A systematic review. J. Econ. Entomol. 2021, 114, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Mazzi, D.; Dorn, S. Movement of insect pests in agricultural landscapes. Ann. Appl. Biol. 2012, 160, 97–113. [Google Scholar] [CrossRef]
- Simonsen, T.J.; Brown, R.L.; Sperling, F.A. Tracing an invasion: Phylogeography of Cactoblastis cactorum (Lepidoptera: Pyralidae) in the United States based on mitochondrial DNA. Ann. Entomol. Soc. Am. 2008, 101, 899–905. [Google Scholar] [CrossRef]
- Wang, Q.; Bao, W.F.; Yang, F.; Xu, B.; Yang, Y.Z. The specific host plant DNA detection suggests a potential migration of Apolygus lucorum from cotton to mungbean fields. PLoS ONE 2017, 12, e0177789. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Bao, W.; Zhang, Q.; Fu, X.; Yang, Y.; Lu, Y. Host plant use of a polyphagous mirid, Apolygus lucorum: Molecular evidence from migratory individuals. Ecol. Evol. 2019, 9, 11518–11528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.; Kandemir, I.; Walsh, D.B.; Zalom, F.G.; Lavine, L.C. Identification of Lygus hesperus by DNA barcoding reveals insignificant levels of genetic structure among distant and habitat diverse populations. PLoS ONE 2012, 7, e34528. [Google Scholar] [CrossRef] [Green Version]
- Bafeel, S.O.; Arif, I.A.; Bakir, M.A.; Al Homaidan, A.A.; Al Farhan, A.H.; Khan, H.A. DNA barcoding of arid wild plants using rbcL gene sequences. Genet. Mol. Res. 2012, 11, 1934–1941. [Google Scholar] [CrossRef]
- De la Cadena, G.; Papadopoulou, A.; Maes, J.M.; Gómez-Zurita, J. Evaluation of bias on the assessment of diet breadth of herbivorous insects using molecular methods. Insect Sci. 2017, 24, 194–209. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Gravel, D.; He, F. Seeing is believing? Comparing plant–herbivore networks constructed by field co-occurrence and DNA barcoding methods for gaining insights into network structures. Ecol. Evol. 2019, 9, 1764–1776. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heyde, M.; Bunce, M.; Dixon, K.W.; Fernandes, K.; Majer, J.; Wardell-Johnson, G.; White, N.E.; Nevill, P. Evaluating Restoration Trajectories Using DNA Metabarcoding of Ground-dwelling and Airborne Invertebrates and Associated Plant Communities. Mol. Ecol. 2022, 31, 2172–2188. [Google Scholar] [CrossRef] [PubMed]
- Dugal, L.; Thomas, L.; Wilkinson, S.P.; Richards, Z.T.; Alexander, J.B.; Adam, A.A.S.; Kennington, W.J.; Jarman, S.; Ryan, N.M.; Bunce, M.; et al. Coral Monitoring in Northwest Australia with Environmental DNA Metabarcoding Using a Curated Reference Database for Optimized Detection. Environ. DNA 2022, 4, 63–76. [Google Scholar] [CrossRef]
- Stat, M.; Huggett, M.J.; Bernasconi, R.; DiBattista, J.D.; Berry, T.E.; Newman, S.J.; Harvey, E.S.; Bunce, M. Ecosystem Biomonitoring with EDNA: Metabarcoding across the Tree of Life in a Tropical Marine Environment. Sci. Rep. 2017, 7, 12240. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heyde, M.; Bunce, M.; Wardell-Johnson, G.; Fernandes, K.; White, N.E.; Nevill, P. Testing multiple substrates for terrestrial biodiversity monitoring using environmental DNA metabarcoding. Mol. Ecol. Res. 2020, 20, 732–745. [Google Scholar] [CrossRef] [PubMed]
- Van Der Heyde, M.; Bateman, P.W.; Bunce, M.; Wardell-Johnson, G.; White, N.E.; Nevill, P. Scat DNA provides important data for effective monitoring of mammal and bird biodiversity. Biodivers. Conserv. 2021, 30, 3585–3602. [Google Scholar] [CrossRef]
- Avanesyan, A.; Illahi, N.; Lamp, W.O. Detecting ingested host plant DNA in potato leafhopper (Hemiptera: Cicadellidae): Potential use of molecular markers for gut content analysis. J. Econ. Entomol. 2021, 114, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Siegert, C.M.; Levia, D.F. Seasonal and meteorological effects on differential stemflow funneling ratios for two deciduous tree species. J. Hydrol. 2014, 519, 446–454. [Google Scholar] [CrossRef]
- Avanesyan, A. Plant DNA detection from grasshopper gut contents: A step-by-step protocol, from tissues preparation to obtaining plant DNA sequences. Appl. Plant Sci. 2014, 2, 1300082. [Google Scholar] [CrossRef]
- Avanesyan, A.; Culley, T.M. Feeding preferences of Melanoplus femurrubrum grasshoppers on native and exotic grasses: Behavioral and molecular approaches. Entomol. Exp. Appl. 2015, 157, 152–163. [Google Scholar] [CrossRef]
- Goecks, J.; Nekrutenko, A.; Taylor, J. Galaxy: A comprehensive approach for supporting accessible, reproducible, and transparent computational research in the life sciences. Genome Biol. 2010, 11, R86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014; Available online: http://www.R-project.org/ (accessed on 1 April 2022).
- Dechaine, A.C.; Sutphin, M.; Leskey, T.C.; Salom, S.M.; Kuhar, T.P.; Pfeiffer, D.G. Phenology of Lycorma delicatula (Hemiptera: Fulgoridae) in Virginia, USA. Environ. Entomol. 2021, 50, 1267–1275. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Kim, S.; Kwon, S.W.; Lee, S.I.; Jablonski, P.G. Defense sequestration associated with narrowing of diet and ontogenetic change to aposematic colours in the spotted lanternfly. Sci. Rep. 2018, 8, 16831. [Google Scholar] [CrossRef] [PubMed]
- Nixon, L.J.; Jones, S.K.; Tang, L.; Urban, J.; Felton, K.; Leskey, T.C. Survivorship and development of the invasive Lycorma delicatula (Hemiptera: Fulgoridae) on wild and cultivated temperate host plants. Environ. Entomol. 2022, 51, 222–228. [Google Scholar] [CrossRef]
- Pearson, C.C.; Backus, E.A.; Shugart, H.J.; Munyaneza, J.E. Characterization and correlation of EPG waveforms of Bactericera cockerelli (Hemiptera: Triozidae): Variability in waveform appearance in relation to applied signal. Ann. Entomol. Soc. Am. 2014, 107, 650–666. [Google Scholar] [CrossRef] [Green Version]
- Essler, J.L.; Kane, S.A.; Collins, A.; Ryder, K.; DeAngelo, A.; Kaynaroglu, P.; Otto, C.M. Egg Masses as Training Aids for Spotted Lanternfly Lycorma Delicatula Detection Dogs. PLoS ONE 2021, 16, e0250945. [Google Scholar] [CrossRef]
- Keller, J.A.; Johnson, A.E.; Uyi, O.; Wurzbacher, S.; Long, D.; Hoover, K. Dispersal of Lycorma delicatula (Hemiptera: Fulgoridae) nymphs through contiguous, deciduous forest. Environ. Entomol. 2020, 49, 1012–1018. [Google Scholar] [CrossRef]
- Wolfin, M.S.; Binyameen, M.; Wang, Y.; Urban, J.M.; Roberts, D.C.; Baker, T.C. Flight dispersal capabilities of female spotted lanternflies (Lycorma delicatula) related to size and mating status. J. Insect Behav. 2019, 32, 188–200. [Google Scholar] [CrossRef]
- Baker, T.C.; Smyers, E.C.; Urban, J.M.; Meng, Z.; Damadaram, K.P.; Myrick, A.J.; Cooperband, M.F.; Domingue, M.J. Progression of seasonal activities of adults of the spotted lanternfly, Lycorma delicatula, during the 2017 season of mass flight dispersal behavior in eastern Pennsylvania. J. Asia-Pac. Entomol. 2019, 22, 705–713. [Google Scholar] [CrossRef]
- Baker, T.C.; Myrick, A.J.; Wolfin, M.S.; Wang, Y. Visual Responses of Flight-Dispersing Spotted Lanternflies, Lycorma delicatula toward a Tall Vertical Silhouette in a Vineyard. J. Insect Behav. 2021, 34, 49–60. [Google Scholar] [CrossRef]
- Domingue, M.J.; Baker, T.C. Orientation of flight for physically disturbed spotted lanternflies, Lycorma delicatula,(Hemiptera, Fulgoridae). J. Asia-Pac. Entomol. 2019, 22, 117–120. [Google Scholar] [CrossRef]
- Cooperband, M.; Murman, K.; Cannon, S.; Abreu, L.; Wallace, M. Dispersal and host preference of marked and released spotted lanternfly. In Otis Laboratory 2018 Annual Report; Trepanowski, N., Vieira, K., Heller, S., Booth, E., Eds.; United States Department of Agriculture: Buzzards Bay, MA, USA, 2019; p. 97. [Google Scholar]
Figure 1.
Location of the collection site at the Fair Hill State Natural Resource Management Area (39°42′36.3′′ N, 75°51′02.98′′ W, Elkton, MD, USA) (a), and first nymphal instars of Lycorma delicatula observed and collected from various plants (b–d) (Photos by Alireza Shokoohi).
Figure 1.
Location of the collection site at the Fair Hill State Natural Resource Management Area (39°42′36.3′′ N, 75°51′02.98′′ W, Elkton, MD, USA) (a), and first nymphal instars of Lycorma delicatula observed and collected from various plants (b–d) (Photos by Alireza Shokoohi).

Figure 2.
Mean quality scores of the readable sequences obtained from each bulk sample and from the reference sample of gut contents of the first nymphal instars of Lycorma delicatula. Sample “1n1a” indicates the reference sample; samples 1n1b-1n10 are the bulk samples.
Figure 2.
Mean quality scores of the readable sequences obtained from each bulk sample and from the reference sample of gut contents of the first nymphal instars of Lycorma delicatula. Sample “1n1a” indicates the reference sample; samples 1n1b-1n10 are the bulk samples.

Figure 3.
The species identity of the ingested plants obtained from both bulk samples and reference sample of the gut contents of the first nymphal instars of Lycorma delicatula. The lines show the trophic interactions between the nymphs and plants that were present at the field site (DNA detected; on the left: 6 out of 19 plants at the site) and the plants that were absent at the field site (on the right). The plants that were present at the field site refer to the plants that were present at the collection site where the insect nymphs were collected, and not to the plants growing in the Fair Hill State Natural Resource Management Area (Elkton, MD, USA). Each box with L. delicatula nymphs shows a separate group of nymphs; the number of nymphs in each box corresponds to the number of nymphs which were ground together to create bulk DNA extracts. The bulk sample labels show the number of bulk samples obtained from each group of nymphs (e.g., 1n1-1n3 represent three bulk samples created from 10 nymphs). Trophic interactions are shown and were analyzed for each group of nymphs separately (i.e., each line of a different color coming from a different box represents separate nymphs), and then the results on ingested plants were summarized to include all the bulk samples and the reference sample. (Photos of the herbarium samples by W. Lamp; drawings of the nymphs by A. Avanesyan; the chart design by C. McPherson).
Figure 3.
The species identity of the ingested plants obtained from both bulk samples and reference sample of the gut contents of the first nymphal instars of Lycorma delicatula. The lines show the trophic interactions between the nymphs and plants that were present at the field site (DNA detected; on the left: 6 out of 19 plants at the site) and the plants that were absent at the field site (on the right). The plants that were present at the field site refer to the plants that were present at the collection site where the insect nymphs were collected, and not to the plants growing in the Fair Hill State Natural Resource Management Area (Elkton, MD, USA). Each box with L. delicatula nymphs shows a separate group of nymphs; the number of nymphs in each box corresponds to the number of nymphs which were ground together to create bulk DNA extracts. The bulk sample labels show the number of bulk samples obtained from each group of nymphs (e.g., 1n1-1n3 represent three bulk samples created from 10 nymphs). Trophic interactions are shown and were analyzed for each group of nymphs separately (i.e., each line of a different color coming from a different box represents separate nymphs), and then the results on ingested plants were summarized to include all the bulk samples and the reference sample. (Photos of the herbarium samples by W. Lamp; drawings of the nymphs by A. Avanesyan; the chart design by C. McPherson).

Figure 4.
Ingested plant species, obtained from both bulk samples and reference sample of the gut contents of the first nymphal instars of Lycorma delicatula. (a) Overall proportion of unique plant families; (b) overall proportion of ingested plants with different plant origin (“Both” represents Betula sp. and Hydrangea sp.); and (c) overall proportion of ingested plants with various life forms.
Figure 4.
Ingested plant species, obtained from both bulk samples and reference sample of the gut contents of the first nymphal instars of Lycorma delicatula. (a) Overall proportion of unique plant families; (b) overall proportion of ingested plants with different plant origin (“Both” represents Betula sp. and Hydrangea sp.); and (c) overall proportion of ingested plants with various life forms.

Figure 5.
Ingested plant species, obtained from the bulk samples only. The pie chart shows the proportion of unique plant species across the bulk samples shown on Figure 2 which represented separate groups of Lycorma delicatula nymphs (i.e., “1n1-1n3”, “1n5”, “1n5-1n6”, “1n7-1n8”, and “1n9-1n10). The percentage of plant species was calculated across unique plant species identified for each group of bulk samples.
Figure 5.
Ingested plant species, obtained from the bulk samples only. The pie chart shows the proportion of unique plant species across the bulk samples shown on Figure 2 which represented separate groups of Lycorma delicatula nymphs (i.e., “1n1-1n3”, “1n5”, “1n5-1n6”, “1n7-1n8”, and “1n9-1n10). The percentage of plant species was calculated across unique plant species identified for each group of bulk samples.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Identified reference plant species collected at the field site at the Fair Hill State Natural Resource Management Area (Elkton, MD, USA). Both the plants on which the nymphs of Lycorma delicatula were observed, and the plants that were free from infestation were collected.
Table 1.
Identified reference plant species collected at the field site at the Fair Hill State Natural Resource Management Area (Elkton, MD, USA). Both the plants on which the nymphs of Lycorma delicatula were observed, and the plants that were free from infestation were collected.
| Sample ID | Plant Family | Plant Species ID | Common Name | Nymphs Presence | Date Submitted to GenBank | GenBank Accession Number |
|---|---|---|---|---|---|---|
| p001 | Rosaceae | Rosa sp. | Rose | + | 12/27/2021 | OM038103 |
| p002 | Rosaceae | Rubus phoenicolasius | Wine raspberry | + | 12/27/2021 | OM038104 |
| p003 | Elaeagnaceae | Elaeagnus umbellata | Autumn olive | − | 12/27/2021 | OM038105 |
| p004 | Apocynaceae | Apocynum cannabinum | Indian hemp | − | 12/27/2021 | OM038106 |
| p005 | Simaroubaceae | Ailanthus altissima | Tree of heaven | + | 1/31/2022 | OM470964 |
| p006 | Polygonaceae | Persicaria virginiana | Jumpseed | − | 1/31/2022 | OM470965 |
| p007 | Celastraceae | Celastrus orbiculatus | Oriental bittersweet | + | 11/7/2021 | OL539536 |
| p008 | Brassicaceae | Alliaria petiolata | Garlic mustard | − | 11/21/2021 | OL603937 |
| p009 | Poaceae | Festuca sp. | Fescue | − | 11/24/2021 | OL657222 |
| p010 | Fabaceae | Vicia sativa | Common vetch | − | 11/24/2021 | OL657223 |
| p011 | Vitaceae | Ampelopsis glandulosa | Amur peppervine | − | 1/31/2022 | OM470966 |
| p012 | Celastraceae | Celastrus orbiculatus | Oriental bittersweet | − | 1/31/2022 | OM470967 |
| p013 | Poaceae | Dactylis glomerata | Cat grass | − | 2/4/2022 | OM523099 |
| p014 | Rosaceae | Rubus cuneifolius | Sand blackberry | − | 2/4/2022 | OM523098 |
| p015 | Sapindaceae | Acer negundo | Boxelder maple | − | 2/11/2022 | OM672995 |
| p016 | Celastraceae | Celastrus orbiculatus | Oriental bittersweet | − | 2/11/2022 | OM672996 |
| p017 | Celastraceae | Celastrus orbiculatus | Oriental bittersweet | − | 2/4/2022 | OM523100 |
| p018 | Boraginaceae | Lappula squarrosa | Bristly sheepburr | − | 2/4/2022 | OM523101 |
| p019 | Poaceae | Dactylis glomerata | Cat grass | − | 2/11/2022 | OM672997 |
| p020 | Rosaceae | Rubus occidentalis | Black Raspberry | − | 2/11/2022 | OM672998 |
| p021 | Elaeagnaceae | Elaeagnus umbellata | Autumn olive | − | 2/11/2022 | OM672999 |
| p022 | Elaeagnaceae | Elaeagnus umbellata | Autumn olive | − | 3/8/2022 | OM964639 |
| p023 | Vitaceae | Ampelopsis glandulosa | Amur peppervine | − | 2/11/2022 | OM673000 |
| p024 | Sapindaceae | Acer negundo | Boxelder maple | − | 2/11/2022 | OM673001 |
| p025 | Magnoliaceae | Liriodendron tulipifera | Tulip tree | − | 2/11/2022 | OM673002 |
| p026 | Rosaceae | Prunus serotina | Black cherry | − | 2/11/2022 | OM673003 |
| p027 | Rosaceae | Rubus idaeus | Red raspberry | − | 2/11/2022 | OM673004 |
| p028 | Boraginaceae | Lappula squarrosa | Bristly sheepburr | − | 2/11/2022 | OM673005 |
Table 2.
PCR components and conditions used to amplify a portion of rbcL gene in this study.
| Primers and Other PCR Components | Concentration/Volume (for 20 μL of PCR Reaction) | PCR Conditions |
|---|---|---|
| rbcLaF (5′-ATG TCA CCA CAA ACA GAG ACT AAA GC-3′) | 2 μM/2 μL | initial denaturation: 94 °C for 4 min 35 cycles of 94 °C for 30 s, 57 °C for 30 s, and 72 °C for 30 s final extension: 72 °C for 2 min |
| rbcLaR (5′-GTA AAA TCA AGT CCA CCR CG-3′) | 2 μM/2 μL | |
| 2X PCR PreMix with Dye (Syd Labs Inc., Natick, MA, USA) | 10 μL | |
| ddH2O | 5.2 μL | |
| template DNA | 0.8 μL |
Table 3.
Basic summary of sequence reads obtained from gut contents of first nymphal instars of Lycorma delicatula and used for identification of ingested plants. Both the reference sample (1n1a-ref) and bulk samples (1n1b-1n10) are included.
Table 3.
Basic summary of sequence reads obtained from gut contents of first nymphal instars of Lycorma delicatula and used for identification of ingested plants. Both the reference sample (1n1a-ref) and bulk samples (1n1b-1n10) are included.
| Sample ID | Total Number of Sequence Reads | Sequence Read Length (bp) | Sequence Length Screened (bp; Consensus Sequences) | Number of Unique Readable Sequences Passed the Screening | Number of Unique Plant Species | Sequence Quality Score (Mean ± SE) * | |
|---|---|---|---|---|---|---|---|
| Forward | Reverse | ||||||
| 1n1a-ref | 347,901 | 347,901 | 35–250 | >150 | 65 | 11 | 34.37 ± 0.11 ac |
| 1n1b | 223,629 | 223,629 | 35–250 | >100 | 3 | 3 | 34.11 ± 0.12 ac |
| 1n2 | 117,181 | 117,181 | 35–250 | >100 | 1 | 1 | 33.53 ± 0.12 bc |
| 1n3 | 176,223 | 176,223 | 35–250 | >100 | 9 | 3 | 33.59 ± 0.11 c |
| 1n4 | 157,468 | 157,468 | 35–250 | >100 | 19 | 12 | 33.95 ± 0.10 c |
| 1n5 | 169,266 | 169,266 | 35–250 | >100 | 7 | 3 | 33.67 ± 0.09 bc |
| 1n6 | 138,802 | 138,802 | 35–250 | >100 | 5 | 3 | 33.84 ± 0.10 bc |
| 1n7 | 163,404 | 163,404 | 35–250 | >100 | 11 | 6 | 34.11 ± 0.08 ac |
| 1n8 | 160,706 | 160,706 | 35–250 | >100 | 3 | 1 | 34.53 ± 0.11 a |
| 1n9 | 195,474 | 195,474 | 35–250 | >100 | 7 | 5 | 34.46 ± 0.14 a |
| 1n10 | 198,622 | 198,622 | 35–250 | >100 | 4 | 3 | 34.43 ± 0.16 ac |
* The mean sequence quality scores that do not share a letter are significantly different at α = 0.05.
Table 4.
The species identity, origin, and life form of all the ingested plants obtained from the gut contents of the first nymphal instars of Lycorma delicatula. Both reference sample and bulk samples are included.
Table 4.
The species identity, origin, and life form of all the ingested plants obtained from the gut contents of the first nymphal instars of Lycorma delicatula. Both reference sample and bulk samples are included.
| Plant Family | Plant Species | Common Name | Seq. Length (bp) | Highest Match, % | Plant Origin | Plant Life Form | Presence at the Site | Reported As a Host * | |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Asteraceae | Ageratina altissima | White snakeroot | 285 | 99.65 | Native | Herbaceous Perennial | − | − |
| 2 | Asteraceae | Mikania scandens | Climbing hempvine | 225 | 99.55 | Native | Woody Vine | − | − |
| 3 | Betulaceae | Betula sp. | Birch | 406 | 100 | Both | Tree | − | + |
| 4 | Brassicaceae | Cardamine impatiens | Narrowleaf bittercress | 258 | 100 | Introduced | Herbaceous Biennial | − | − |
| 5 | Caprifoliaceae | Lonicera maackii | Amur honeysuckle | 247 | 100 | Introduced | Shrub | − | + |
| 6 | Celastraceae | Celastrus orbiculatus | Oriental bittersweet | 404 | 100 | Introduced | Woody Vine | + | + |
| 7 | Cornaceae | Nyssa sylvatica | Blackgum | 362 | 100 | Native | Tree | − | + |
| 8 | Fabaceae | Gleditsia triacanthos | Honeylocust | 260 | 99.2 | Native | Shrub | − | − |
| 9 | Fabaceae | Medicago sativa | Alfalfa | 184 | 100 | Introduced | Herbaceous Perennial | − | + |
| 10 | Fagaceae | Quercus coccinea | Scarlet oak | 336 | 99.11 | Native | Tree | − | + |
| 11 | Hydrangeaceae | Hydrangea sp. | Hydrangea | 115 | 100 | Both | Shrub | − | − |
| 12 | Juglandaceae | Carya illinoinensis | Pecan | 367 | 100 | Native | Tree | − | + |
| 13 | Lauraceae | Litsea sp. | Litsea | 260 | 99.6 | Native | Shrub | − | − |
| 14 | Magnoliaceae | Liriodendron tulipifera | Tuliptree | 292 | 99 | Native | Tree | + | + |
| 15 | Moraceae | Maclura pomifera | Osage-orange | 190 | 100 | Native | Tree | − | − |
| 16 | Moraceae | Morus alba | White mulberry | 357 | 99.72 | Introduced | Tree | − | + |
| 17 | Pinaceae | Pinus strobus | Eastern white pine | 239 | 100 | Native | Tree | − | + |
| 18 | Poaceae | Agrostis stolonifera | Creeping bentgrass | 158 | 100 | Introduced | Herbaceous perennial | − | − |
| 19 | Poaceae | Anthoxanthum odoratum | Sweet vernalgrass | 150 | 100 | Introduced | Herbaceous Perennial | − | − |
| 20 | Poaceae | Festuca sp. | Fescue | 206 | 100 | Introduced | Herbaceous Perennial | + | + |
| 21 | Poaceae | Hierochloe odorata | Sweetgrass | 218 | 100 | Native | Herbaceous Perennial | − | − |
| 22 | Poaceae | Lolium arundinaceum | Tall fescue | 346 | 99.7 | Introduced | Herbaceous Annual | − | − |
| 23 | Poaceae | Milium effusum | American milletgrass | 110 | 100 | Native | Herbaceous Perennial | − | − |
| 24 | Poaceae | Polypogon monspeliensis | Annual rabbitsfoot grass | 127 | 99.21 | Introduced | Herbaceous Annual | − | − |
| 25 | Rosaceae | Prunus sp. | Plum | 200 | 100 | Native | Tree/Shrub | + | + |
| 26 | Rosaceae | Rosa sp. | Rose | 259 | 100 | Introduced | Woody Vine | + | + |
| 27 | Simaroubaceae | Ailanthus altissima | Tree of heaven | 300 | 99.67 | Introduced | Tree | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
McPherson, C.; Avanesyan, A.; Lamp, W.O. Diverse Host Plants of the First Instars of the Invasive Lycorma delicatula: Insights from eDNA Metabarcoding. Insects 2022, 13, 534. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060534
AMA Style
McPherson C, Avanesyan A, Lamp WO. Diverse Host Plants of the First Instars of the Invasive Lycorma delicatula: Insights from eDNA Metabarcoding. Insects. 2022; 13(6):534. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060534
Chicago/Turabian StyleMcPherson, Cameron, Alina Avanesyan, and William O. Lamp. 2022. "Diverse Host Plants of the First Instars of the Invasive Lycorma delicatula: Insights from eDNA Metabarcoding" Insects 13, no. 6: 534. https://0-doi-org.brum.beds.ac.uk/10.3390/insects13060534
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.