Measurement of Multiple Cardiac Performance Endpoints in Daphnia and Zebrafish by Kymograph

 ,
,  ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish and Daphnia Maintenance and Sample Preparation
2.2. Instrument Setting for Temperature Control
2.3. Astemizole Treatment
2.4. Video Capturing and Kymograph Generation
2.5. Heart Rate and Heart Rate Variability Measurement
2.6. Cardiac Parameter Endpoint Calculation
2.7. Statistics
3. Results
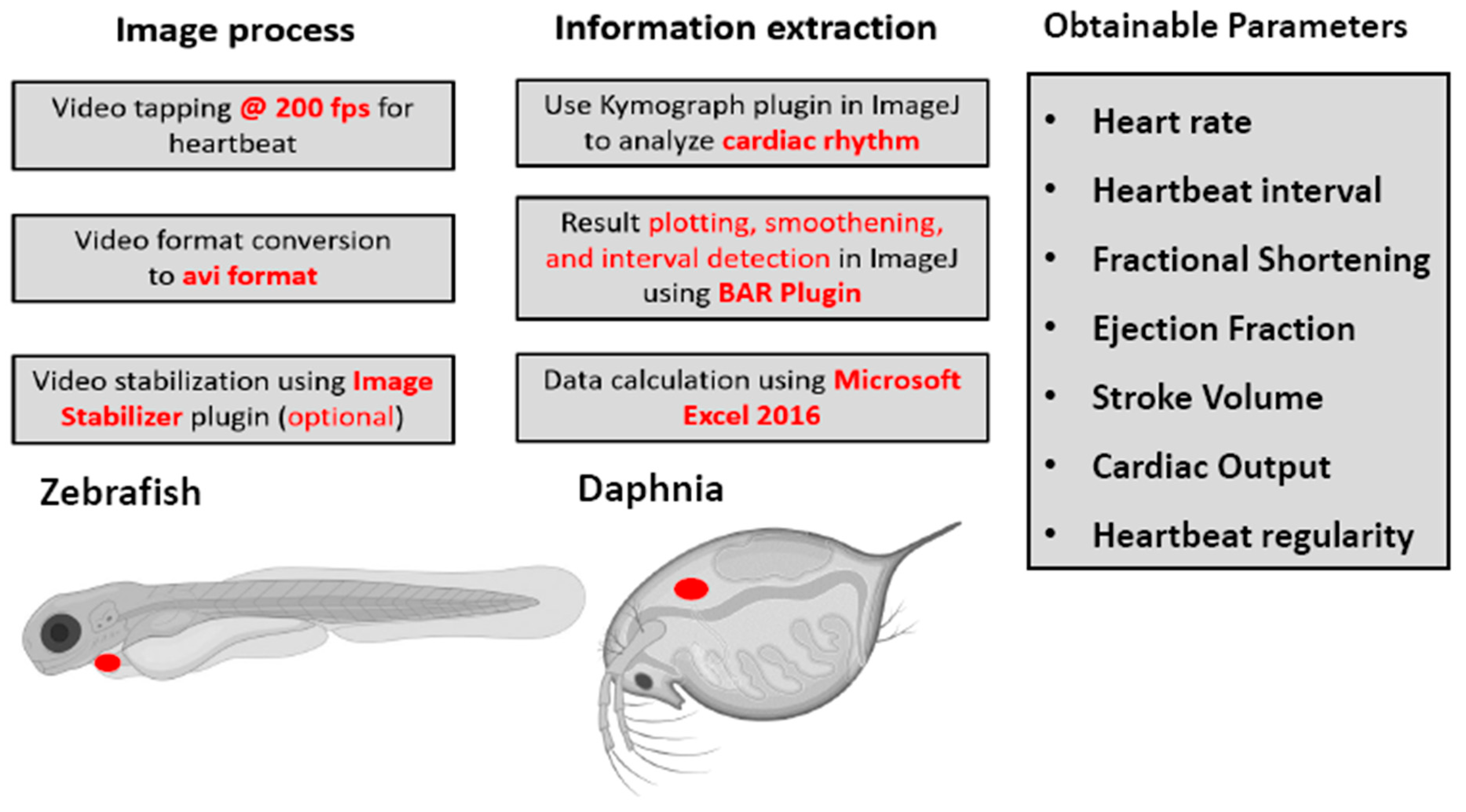
3.1. Overview of Experimental Workflow
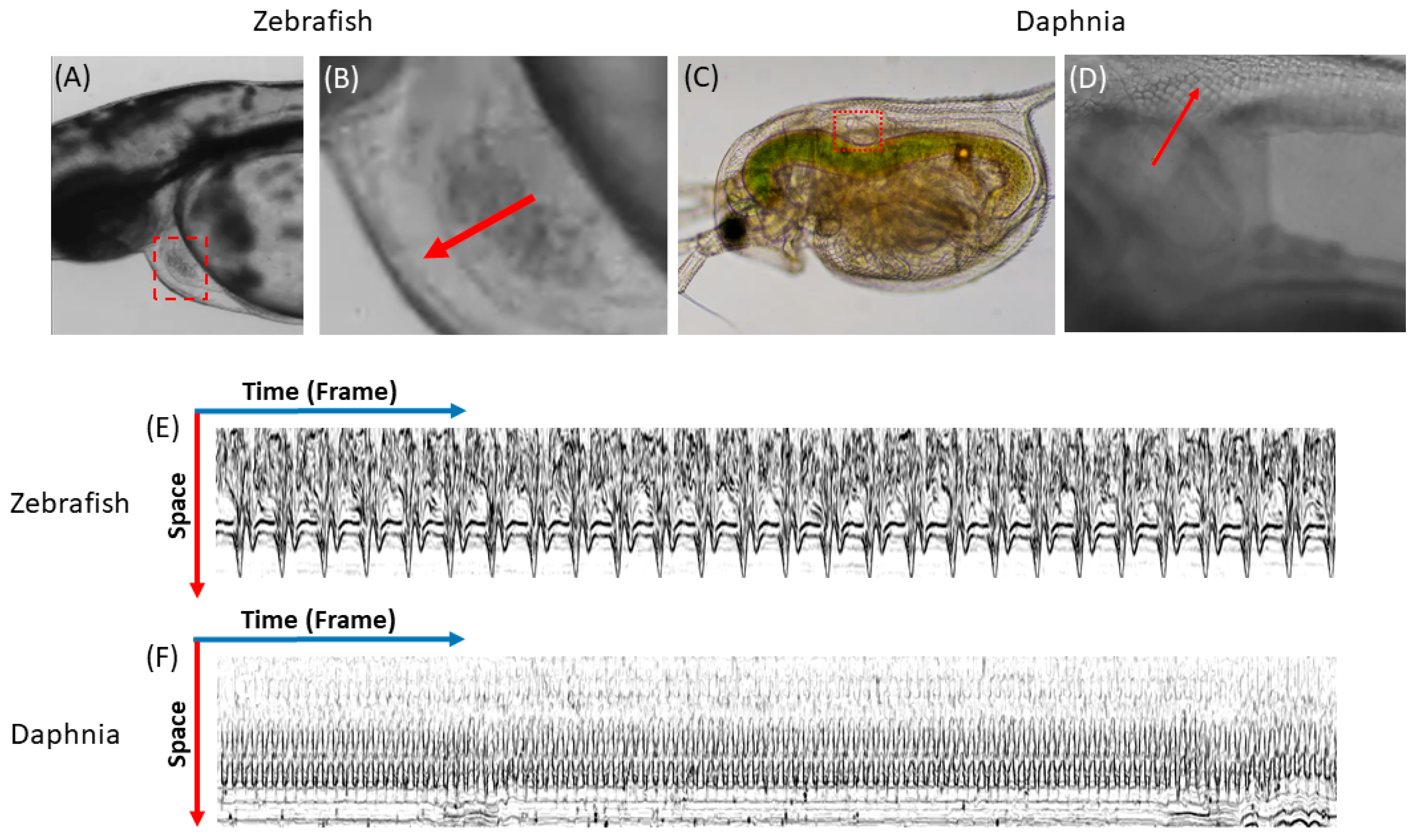
3.2. Measurement of Heart Rate by Kymograph in Zebrafish and Daphnia Magna
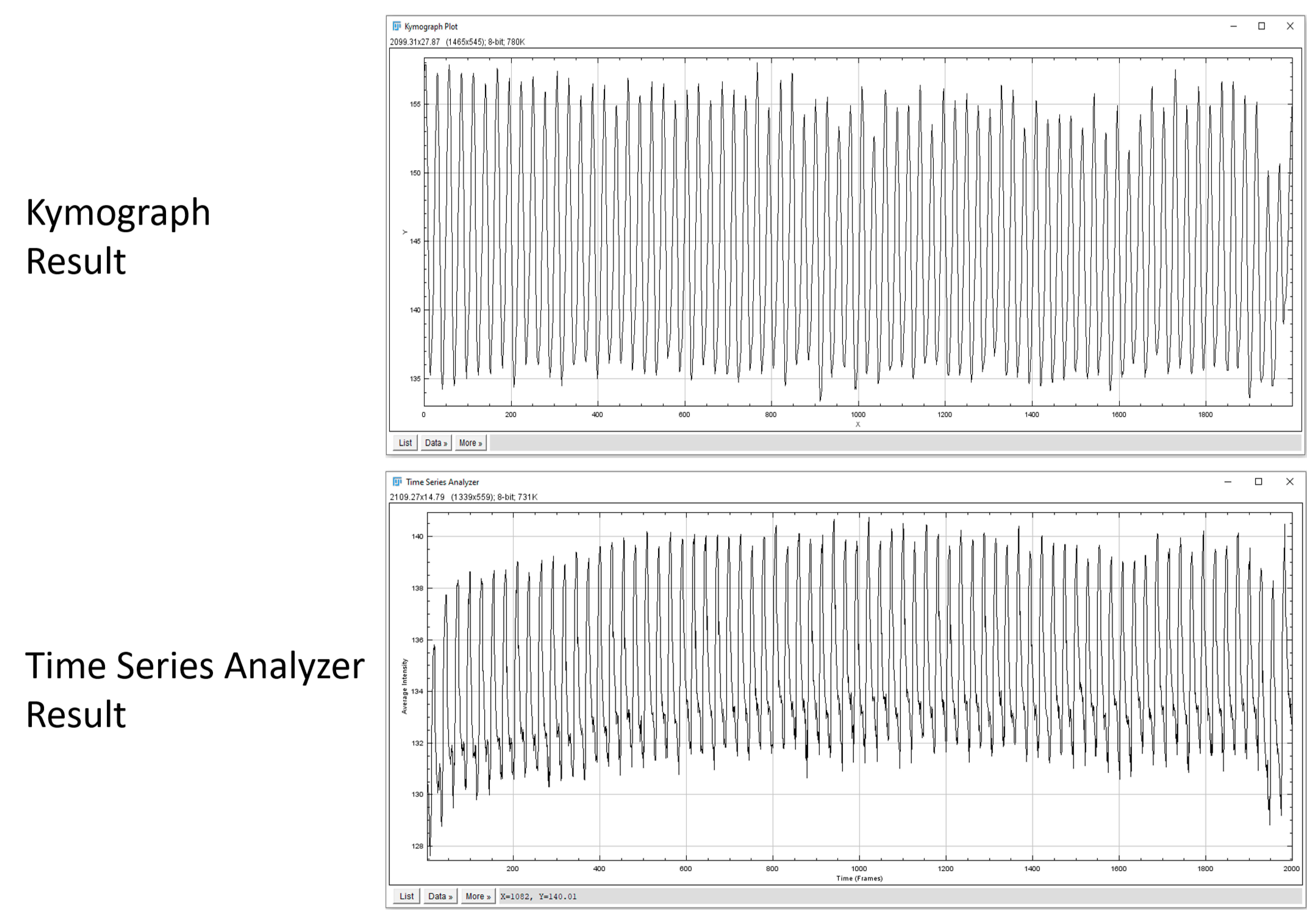
3.3. Comparison of Different Measuring Methods on Heart Rate Variability in Daphnia Magna
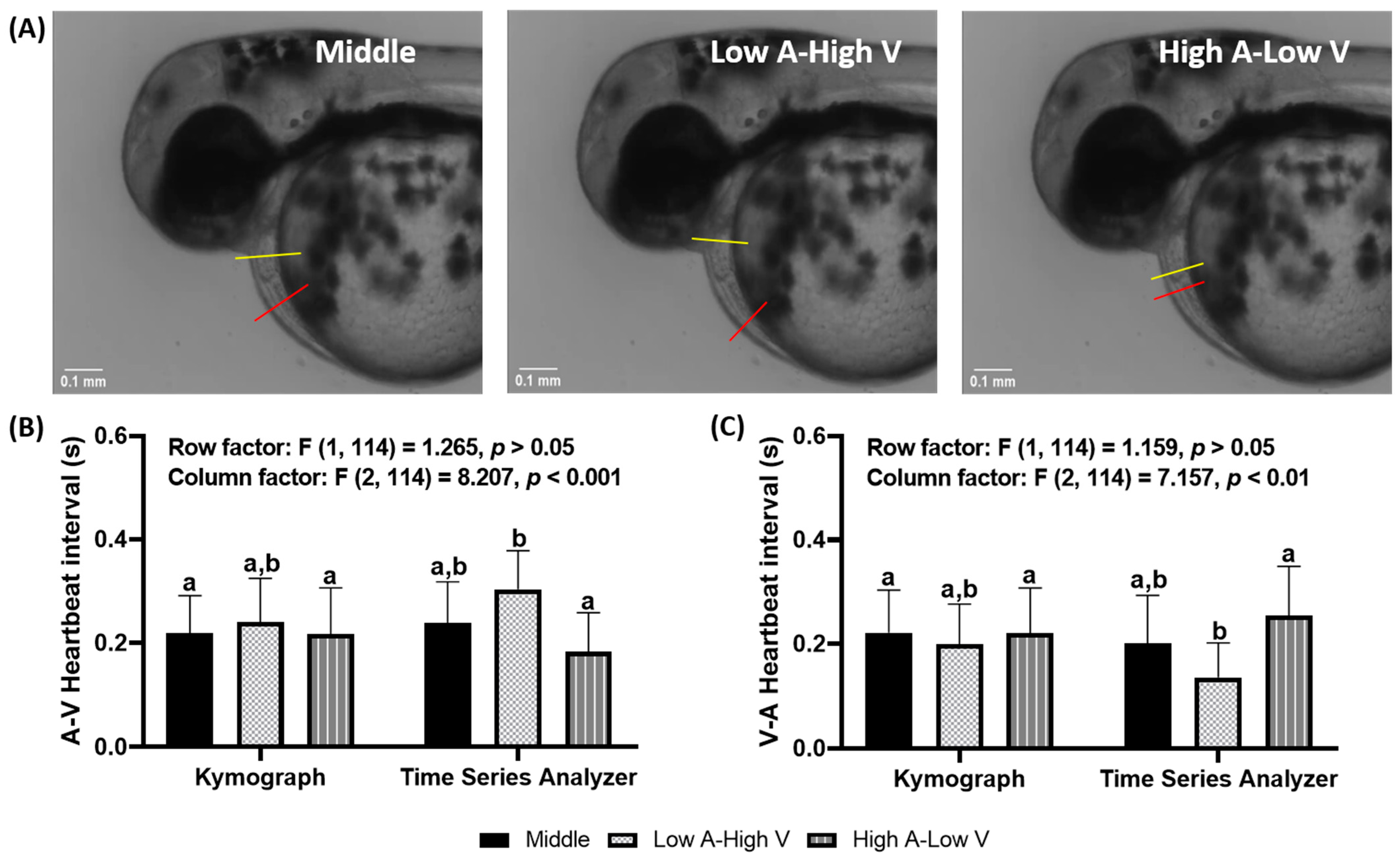
3.4. Effects of Region of Interest (ROI) Position on Heartbeat Regularity in Zebrafish
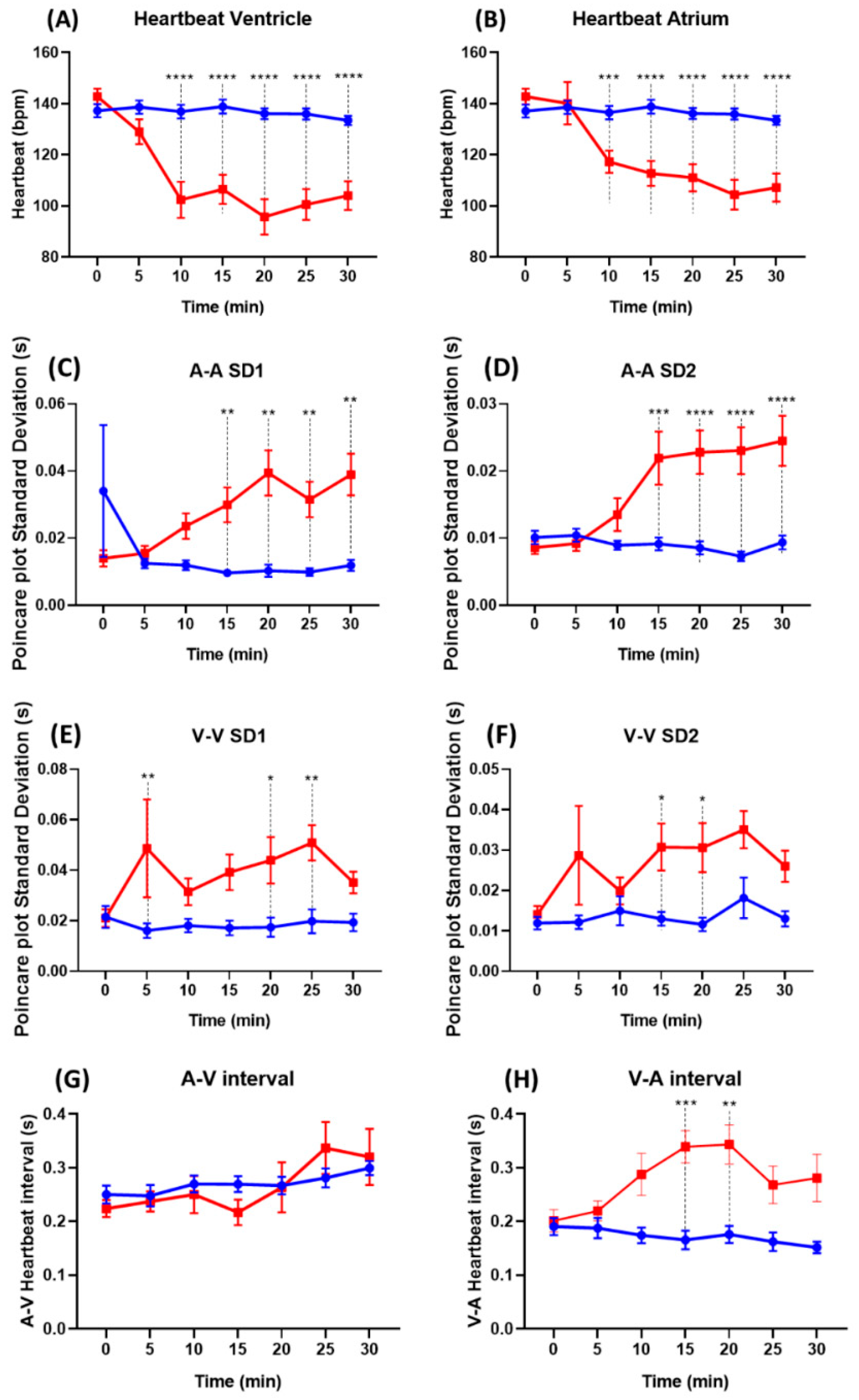
3.5. Comparison of the Heartbeat Regularity after Astemizole Treatment in Zebrafish
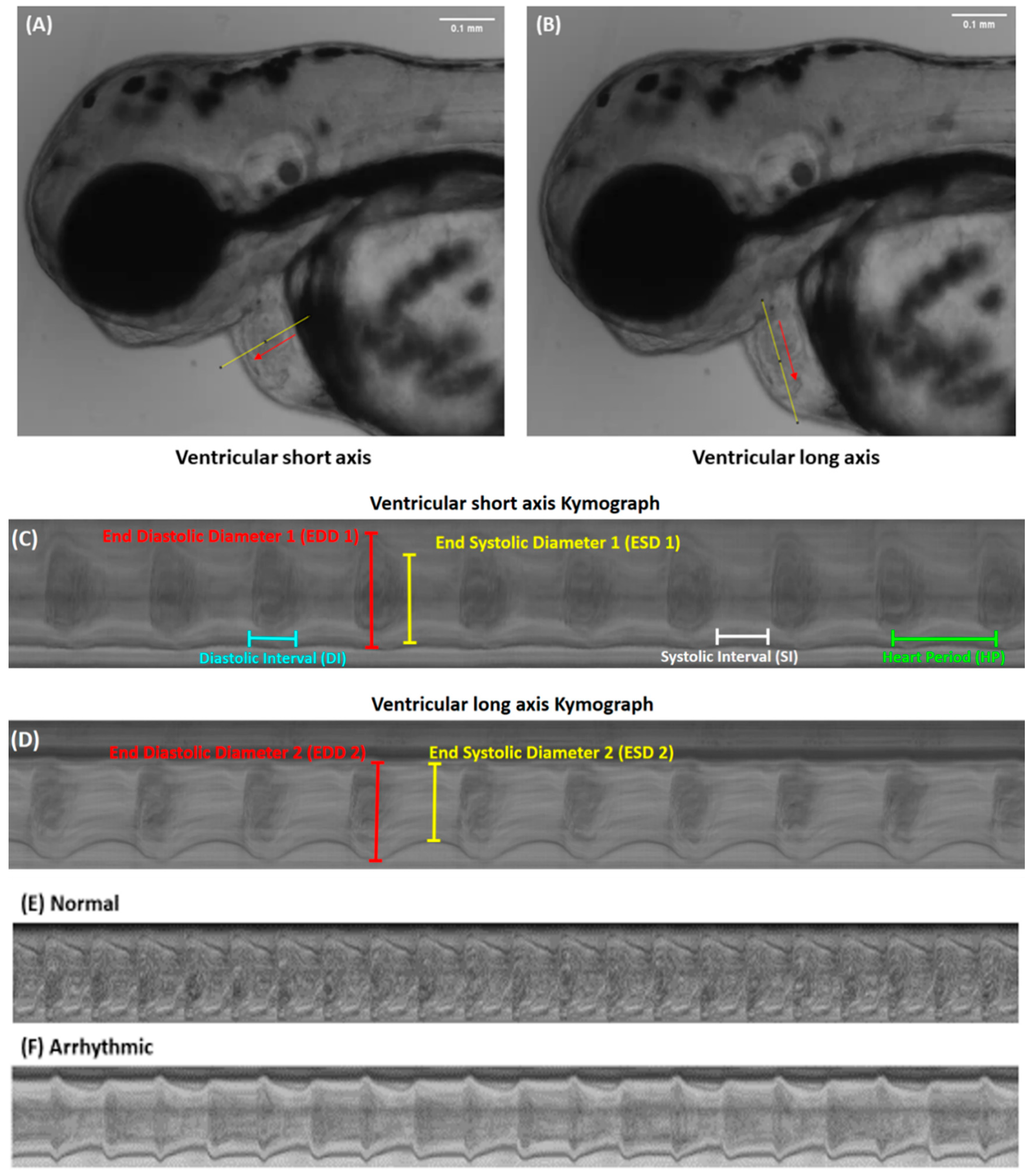
3.6. Measurement of Multiple Cardiac Performance Endpoints by Using Kymograph
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

References
- World Health Organization. Hearts: Technical Package for Cardiovascular Disease Management in Primary Health Care; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Gut, P.; Reischauer, S.; Stainier, D.Y.R.; Arnaout, R. Little fish, big data: Zebrafish as a model for cardiovascular and metabolic disease. Physiol. Rev. 2017, 97, 889–938. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bournele, D.; Beis, D. Zebrafish models of cardiovascular disease. Heart Fail. Rev. 2016, 21, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Guilhermino, L.; Diamantino, T.; Silva, M.C.; Soares, A. Acute toxicity test with daphnia magna: An alternative to mammals in the prescreening of chemical toxicity? Ecotoxicol. Environ. Saf. 2000, 46, 357–362. [Google Scholar] [CrossRef] [Green Version]
- Greene, M.; Pitts, W.; Dewprashad, B. Using videography to study the effects of stimulants on daphnia magna. Am. Biol. Teach. 2017, 79, 35–40. [Google Scholar] [CrossRef]
- Campbell, A.K.; Wann, K.T.; Matthews, S.B. Lactose causes heart arrhythmia in the water flea daphnia pulex. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 139, 225–234. [Google Scholar] [CrossRef]
- Villegas-Navarro, A.; Rosas-L, E.; Reyes, J.L. The heart of daphnia magna: Effects of four cardioactive drugs. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2003, 136, 127–134. [Google Scholar] [CrossRef]
- Gaikwad, N.; Panat, A.; Deshpande, M.; Ramya, K.; Khalid, P.; Augustine, P. Effect of shilajit on the heart of daphnia: A preliminary study. J. Ayurveda Integr. Med. 2012, 3, 3. [Google Scholar]
- Hoage, T.; Ding, Y.; Xu, X. Quantifying cardiac functions in embryonic and adult zebrafish. In Cardiovascular Development; Peng, X., Antonyak, M., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 11–20. [Google Scholar]
- Sampurna, B.P.; Audira, G.; Juniardi, S.; Lai, Y.-H.; Hsiao, C.-D. A simple imagej-based method to measure cardiac rhythm in zebrafish embryos. Inventions 2018, 3, 21. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, C.-D.; Wu, H.-H.; Malhotra, N.; Liu, Y.-C.; Wu, Y.-H.; Lin, Y.-N.; Saputra, F.; Santoso, F.; Chen, K.H.-C. Expression and purification of recombinant ghk tripeptides are able to protect against acute cardiotoxicity from exposure to waterborne-copper in zebrafish. Biomolecules 2020, 10, 1202. [Google Scholar] [CrossRef]
- Jeong, T.-Y.; Yoon, D.; Kim, S.; Kim, H.Y.; Kim, S.D. Mode of action characterization for adverse effect of propranolol in daphnia magna based on behavior and physiology monitoring and metabolite profiling. Environ. Pollut. 2018, 233, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.-L.; Lin, Y.-H.; Tsai, I.-J.; Hsieh, F.-J.; Tsai, H.-J. In vivo assessment of cardiac morphology and function in heart-specific green fluorescent zebrafish. J. Formos. Med. Assoc. 2007, 106, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Xing, Q.; Huynh, V.; Parolari, T.G.; Maurer-Morelli, C.V.; Peixoto, N.; Wei, Q. Zebrafish larvae heartbeat detection from body deformation in low resolution and low frequency video. Med. Biol. Eng. Comput. 2018, 56, 2353–2365. [Google Scholar] [CrossRef]
- Gierten, J.; Pylatiuk, C.; Hammouda, O.T.; Schock, C.; Stegmaier, J.; Wittbrodt, J.; Gehrig, J.; Loosli, F. Automated high-throughput heartbeat quantification in medaka and zebrafish embryos under physiological conditions. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Akerberg, A.A.; Burns, C.E.; Burns, C.G.; Nguyen, C. Deep learning enables automated volumetric assessments of cardiac function in zebrafish. Dis. Models Mech. 2019, 12, 40188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pylatiuk, C.; Sanchez, D.; Mikut, R.; Alshut, R.; Reischl, M.; Hirth, S.; Rottbauer, W.; Just, S. Automatic zebrafish heartbeat detection and analysis for zebrafish embryos. Zebrafish 2014, 11, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Lenning, M.; Fortunato, J.; Le, T.; Clark, I.; Sherpa, A.; Yi, S.; Hofsteen, P.; Thamilarasu, G.; Yang, J.; Xu, X. Real-time monitoring and analysis of zebrafish electrocardiogram with anomaly detection. Sensors 2018, 18, 61. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.Y.; Wu, S.Q.; Guo, S.Y.; Yang, H.; Xia, B.; Li, P.; Li, C.Q. A zebrafish heart failure model for assessing therapeutic agents. Zebrafish 2018, 15, 243–253. [Google Scholar] [CrossRef]
- Martin, W.K.; Tennant, A.H.; Conolly, R.B.; Prince, K.; Stevens, J.S.; DeMarini, D.M.; Martin, B.L.; Thompson, L.C.; Gilmour, M.I.; Cascio, W.E. High-throughput video processing of heart rate responses in multiple wild-type embryonic zebrafish per imaging field. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Schutera, M.; Just, S.; Gierten, J.; Mikut, R.; Reischl, M.; Pylatiuk, C. Machine learning methods for automated quantification of ventricular dimensions. Zebrafish 2019, 16, 542–545. [Google Scholar] [CrossRef]
- Nasrat, S.; Marcato, D.; Hirth, S.; Reischl, M.; Pylatiuk, C. Semi-automated detection of fractional shortening in zebrafish embryo heart videos. Curr. Dir. Biomed. Eng. 2016, 2, 233–236. [Google Scholar] [CrossRef]
- Gaur, H.; Pullaguri, N.; Nema, S.; Purushothaman, S.; Bhargava, Y.; Bhargava, A. Zebrapace: An open-source method for cardiac-rhythm estimation in untethered zebrafish larvae. Zebrafish 2018, 15, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Welzel, O.; Knörr, J.; Stroebel, A.M.; Kornhuber, J.; Groemer, T.W. A fast and robust method for automated analysis of axonal transport. Eur. Biophys. J. 2011, 40, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Smal, I.; Grigoriev, I.; Akhmanova, A.; Niessen, W.J.; Meijering, E. Microtubule dynamics analysis using kymographs and variable-rate particle filters. IEEE Trans. Image Process. 2010, 19, 1861–1876. [Google Scholar] [CrossRef] [Green Version]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. Imagej2: Imagej for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef]
- Lee, L.; Genge, C.E.; Cua, M.; Sheng, X.; Rayani, K.; Beg, M.F.; Sarunic, M.V.; Tibbits, G.F. Functional assessment of cardiac responses of adult zebrafish (danio rerio) to acute and chronic temperature change using high-resolution echocardiography. PLoS ONE 2016, 11, e0145163. [Google Scholar] [CrossRef] [Green Version]
- Müller, M.F.; Colomer, J.; Serra, T. Temperature-driven response reversibility and short-term quasi-acclimation of daphnia magna. PLoS ONE 2018, 13, e0209705. [Google Scholar] [CrossRef]
- Avdesh, A.; Chen, M.; Martin-Iverson, M.T.; Mondal, A.; Ong, D.; Rainey-Smith, S.; Taddei, K.; Lardelli, M.; Groth, D.M.; Verdile, G. Regular care and maintenance of a zebrafish (danio rerio) laboratory: An introduction. J. Vis. Exp. 2012, 69, e4196. [Google Scholar] [CrossRef] [Green Version]
- Hoshi, R.A.; Pastre, C.M.; Vanderlei, L.C.M.; Godoy, M.F. Poincaré plot indexes of heart rate variability: Relationships with other nonlinear variables. Auton. Neurosci. 2013, 177, 271–274. [Google Scholar] [CrossRef]
- Shin, J.T.; Pomerantsev, E.V.; Mably, J.D.; MacRae, C.A. High-resolution cardiovascular function confirms functional orthology of myocardial contractility pathways in zebrafish. Physiol. Genom. 2010, 42, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.X.; Zdanowicz, M.; Young, L.; Kumiski, D.; Leatherbury, L.; Kirby, M.L. Cardiac neural crest in zebrafish embryos contributes to myocardial cell lineage and early heart function. Dev. Dyn. 2003, 226, 540–550. [Google Scholar] [CrossRef]
- Bagatto, B.; Burggren, W. A three-dimensional functional assessment of heart and vessel development in the larva of the zebrafish (danio rerio). Physiol. Biochem. Zool. 2006, 79, 194–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, K. The Image Stabilizer Plugin for Imagej. Available online: https://www.cs.cmu.edu/~kangli/code/Image_Stabilizer.html (accessed on 15 January 2021).
- Ferreira, T.; Hiner, M.; Rueden, C.; Miura, K.; Eglinger, J.; Chef, B. Bar Plugin for Imagej. Available online: https://0-doi-org.brum.beds.ac.uk/10.5281/zenodo.495245 (accessed on 15 January 2021).
- Jeong, T.-Y.; Kim, H.Y.; Kim, S.D. Multi-generational effects of propranolol on daphnia magna at different environmental concentrations. Environ. Pollut. 2015, 206, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Santoso, F.; Farhan, A.; Castillo, A.L.; Malhotra, N.; Saputra, F.; Kurnia, K.A.; Chen, K.H.-C.; Huang, J.-C.; Chen, J.-R.; Hsiao, C.-D. An overview of methods for cardiac rhythm detection in zebrafish. Biomedicines 2020, 8, 329. [Google Scholar] [CrossRef] [PubMed]
- Ott, C.; Lippincott-Schwartz, J. Visualization of live primary cilia dynamics using fluorescence microscopy. Curr. Protoc. Cell Biol. 2012, 57, 4.26.1–4.26.22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, T.; Welch, D.; Nicastro, D.; Dogic, Z. Cilia-like beating of active microtubule bundles. Science 2011, 333, 456–459. [Google Scholar] [CrossRef] [Green Version]
- O’Hagan, R.; Barr, M.M. Kymographic analysis of transport in an individual neuronal sensory cilium in caenorhabditis elegans. In Cilia; Satir, P., Christensen, S., Eds.; Humana Press: New York, NY, USA, 2016; pp. 107–122. [Google Scholar]
- Mukherjee, A.; Jenkins, B.; Fang, C.; Radke, R.J.; Banker, G.; Roysam, B. Automated kymograph analysis for profiling axonal transport of secretory granules. Med. Image Anal. 2011, 15, 354–367. [Google Scholar] [CrossRef] [Green Version]
- Lagerspetz, K.; Perttunen, V. Effect of temperature on the periodic heart beat reversal and heart rate in corethra plumicornis (diptera). J. Insect Physiol. 1962, 8, 621–625. [Google Scholar] [CrossRef]
- Florey, E. Studies on the nervous regulation of the heart beat in decapod crustacea. J. Gen. Physiol. 1960, 43, 1061–1081. [Google Scholar] [CrossRef] [Green Version]
- Motley, H.L. Physiological studies concerning the regulation of heartbeat in freshwater mussels. Physiol. Zool. 1934, 7, 62–84. [Google Scholar] [CrossRef]
- Chakraborty, M.; Llamusi, B.; Artero, R. Modeling of myotonic dystrophy cardiac phenotypes in drosophila. Front. Neurol. 2018, 9, 473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, M.M.; Ahlberg, G.; Hansen, C.V.; Guenther, S.; Marin-Juez, R.; Sokol, A.M.; El-Sammak, H.; Piesker, J.; Hellsten, Y.; Olesen, M.S.; et al. Early sarcomere and metabolic defects in a zebrafish pitx2c cardiac arrhythmia model. Proc. Natl. Acad. Sci. USA 2019, 116, 24115–24121. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Lee, W.-N.; Wang, S.; Konofagou, E.E. P4a-2 an in-vivo study of frame rate optimization for myocardial elastography. In Proceedings of the 2007 IEEE Ultrasonics Symposium, New York, NY, USA, 31 October 2007; IEEE: New York, NY, USA, 2007; pp. 1933–1936. [Google Scholar]
- Gubin, D.G.; Weinert, D.; Rybina, S.V.; Danilova, L.A.; Solovieva, S.V.; Durov, A.M.; Prokopiev, N.Y.; Ushakov, P.A. Activity, sleep and ambient light have a different impact on circadian blood pressure, heart rate and body temperature rhythms. Chronobiol. Int. 2017, 34, 632–649. [Google Scholar] [CrossRef] [PubMed]
- Stauss, H.M. Heart rate variability. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 285, R927–R931. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoso, F.; Krylov, V.V.; Castillo, A.L.; Saputra, F.; Chen, H.-M.; Lai, H.-T.; Hsiao, C.-D. Cardiovascular performance measurement in water fleas by utilizing high-speed videography and imagej software and its application for pesticide toxicity assessment. Animals 2020, 10, 1587. [Google Scholar] [CrossRef]
- Pirow, R.; Wollinger, F.; Paul, R.J. The sites of respiratory gas exchange in the planktonic crustacean daphnia magna: An in vivo study employing blood haemoglobin as an internal oxygen probe. J. Exp. Biol. 1999, 202, 3089–3099. [Google Scholar]
- Smith, S.J. Cardiovascular toxicity of antihistamines. Otolaryngol. Head Neck Surg. 1994, 111, 348–354. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Published Methods | Main Region of Interest Recording Instrument | Tracking Software | Animal Model | Obtainable Result |
|---|---|---|---|---|
| Our study | Inverted Microscope (ICX41, Sunny Optical Technology, Ningbo, Zhejiang, China) + High- Speed CCD (Zgenebio, Taipei, Taiwan) | ImageJ (Fiji) | Zebrafish, AB strain and Daphnia magna | Heart rate, fractional shortening, ejection fraction, stroke volume, cardiac output, heartbeat interval, and chamber-to-chamber interval |
| Sampurna et al., 2018 [11] | CCD mounted onto dissecting microscope | ImageJ | Zebrafish, AB strain | Heart rate |
| Jeong et al., 2018 [13] | Olympus-CKX41SF Microscope (Olympus corporation, Shinjuku, Tokyo, Japan) | Manual counting (Toupview 3.7) | Daphnia magna | Heart rate |
| Ho et al., 2007 [14] | Axioplan microscope (Zeiss, Jeny Germany) with Normaski optics and fluorescence, captured using Sony digital camera | Viewed frame by frame (manual) | Transgenic zebrafish with Green fluorescence protein (GFP) expression | Heart rate, stroke volume, cardiac output |
| Xing et al., 2018 [15] | C3313/RHD2216 + C3100/RHD2000 (Intan Technologies LLC, Los Angeles, CA, USA) and Opticam + Stereo Microscope | MATLAB | Zebrafish, strain not mentioned | Heart rate |
| Gierten et al., 2020 [16] | Brightfield microscope | HeartBeat software (MATLAB) | Unhatched medaka and zebrafish embryos | Heart rate (single and multiple ROI) |
| Akerberg et al., 2019 [17] | Nikon Eclipse 80i compound microscope and Zeiss Lightsheet Z.1 microscope | Deep learning (MATLAB) | Zebrafish strain Tg(myl7:GFP) | Heart rate, fractional shortening, ejection fraction, and cardiac output |
| Pylatiuk et al., 2014 [18] | Automated microscope and Inverted microscope Leica DMIL LED (Leica Camera, Wetzlar, Germany) | MATLAB | Zebrafish AB, Shox2-MO and Isl1l-MO strains | Heart rate (automatic detection) |
| Lenning et al., 2018 [19] | Electrocardiogram recording using MEA membranes | LabVIEW | Zebrafish, strain not mentioned | Heart rate, p waves, QRS complexes, and T waves |
| Zhu et al., 2018 [20] | Dissecting stereomicroscope (Olympus corporation, Shinjuku, Tokyo Japan) | Zebralab Blood Flow System | Zebrafish, AB strain | Heart rate |
| Martin et al., 2019 [21] | Andor Zyla 4.2 sCMOS (Andor Technologies, Belfast, NI) camera mounted to a Nikon Ti microscope (Nikon Corporation, Minato, Tokyo, Japan) | FishRateZ (LabVIEW) | Zebrafish | Heart rate (automatic detection) |
| Schutera et al., 2019 [22] | Not mentioned | Deep learning (PyCharm) | Zebrafish and Medaka | Heart rate, systolic/diastolic volume, ejection fraction, fractional shortening, and stroke volume |
| Nasrat et al., 2016 [23] | Not mentioned | Microsoft Visual Studio Community 2015 | Zebrafish | Fractional shortening |
| Gaur et al., 2018 [24] | Inverted microscope Olympus IX73 Series (Olympus corporation, Shinjuku, Tokyo, Japan) and 10-megapixel camera ProCAM HS-10 MP (Radical Scientific Equipments Private Limited, Ambala, Haryana, India) | ZebraPace (ImageJ based) | Zebrafish | Heart rate |
| Parameters | Control | Astemizole (20 µM) | p |
|---|---|---|---|
| End diastolic volume (µm3) a,b | 490,153.6 ± 182,235.3 | 773,517.1 ± 324,727.3 | ≤0.01 |
| Long axis (µm) a | 140.81 ± 20.18 | 147.1 ± 18.75 | NS |
| Short axis (µm) b | 80.96 ± 16.67 | 98.42 ± 17.85 | ≤0.01 |
| End systolic volume (µm3) a,b | 180,197.1 ± 76,735.82 | 524,531.6 ± 338,855.3 | ≤0.0001 |
| Long axis (µm) a | 106.77 ± 17.16 | 122.34 ± 30.64 | ≤0.01 |
| Short axis (µm) b | 56.38 ± 14.85 | 85.31 ± 22.18 | ≤0.0001 |
| Shortening fraction (%) b | 30.72 ± 13.87 | 13.87 ± 12.10 | ≤0.0001 |
| Ejection fraction (%) b | 62.93 ± 9.97 | 35.70 ± 22.65 | ≤0.001 |
| Stroke volume (pL/beat) a,b | 309.96 ± 134.42 | 248.99 ± 151.00 | NS |
| Heart rate (bpm) b | 133.42 ± 7.84 | 104.00 ± 24.61 | ≤0.0001 |
| Cardiac output (pL/min) a,b | 40,359.96 ± 18,286.35 | 25,351 ± 16,238.36 | ≤0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurnia, K.A.; Saputra, F.; Roldan, M.J.M.; Castillo, A.L.; Huang, J.-C.; Chen, K.H.-C.; Lai, H.-T.; Hsiao, C.-D. Measurement of Multiple Cardiac Performance Endpoints in Daphnia and Zebrafish by Kymograph. Inventions 2021, 6, 8. https://0-doi-org.brum.beds.ac.uk/10.3390/inventions6010008
Kurnia KA, Saputra F, Roldan MJM, Castillo AL, Huang J-C, Chen KH-C, Lai H-T, Hsiao C-D. Measurement of Multiple Cardiac Performance Endpoints in Daphnia and Zebrafish by Kymograph. Inventions. 2021; 6(1):8. https://0-doi-org.brum.beds.ac.uk/10.3390/inventions6010008
Chicago/Turabian StyleKurnia, Kevin Adi, Ferry Saputra, Marri Jmelou M. Roldan, Agnes L. Castillo, Jong-Chin Huang, Kelvin H.-C. Chen, Hong-Thih Lai, and Chung-Der Hsiao. 2021. "Measurement of Multiple Cardiac Performance Endpoints in Daphnia and Zebrafish by Kymograph" Inventions 6, no. 1: 8. https://0-doi-org.brum.beds.ac.uk/10.3390/inventions6010008