Effects of the Extracts from Fruit and Stem of Camellia japonica on Induced Pluripotency and Wound Healing

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Preparation of Plant Extracts
2.2. Cell Culture
2.3. Mice
2.4. High-Throughput Screening
2.5. Plant-Extract-Induced hiPSC Generation
2.6. Immunocytochemistry
2.7. In Vitro Differentiation
2.8. Teratoma Formation and Histological Analysis
2.9. Wound Healing Analysis
2.10. Histopathological Analysis
2.11. RNA Isolation and Quantitative Real-Time Reverse Transcriptase PCR
2.12. Western Blot Analysis
3. Results
3.1. Experimental Results
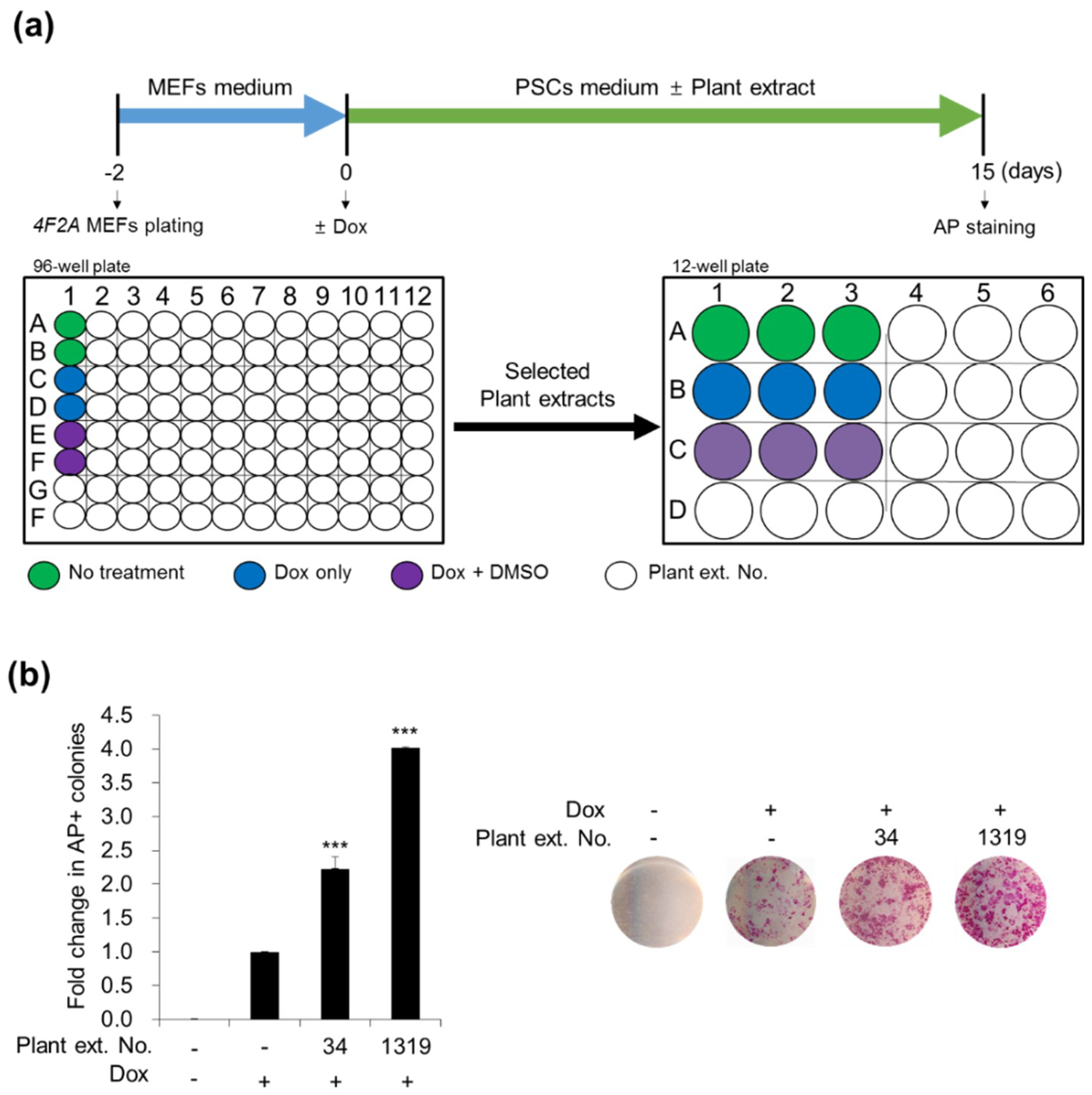
3.1.1. Screening for Plant Extracts That Enhance Cellular Reprogramming
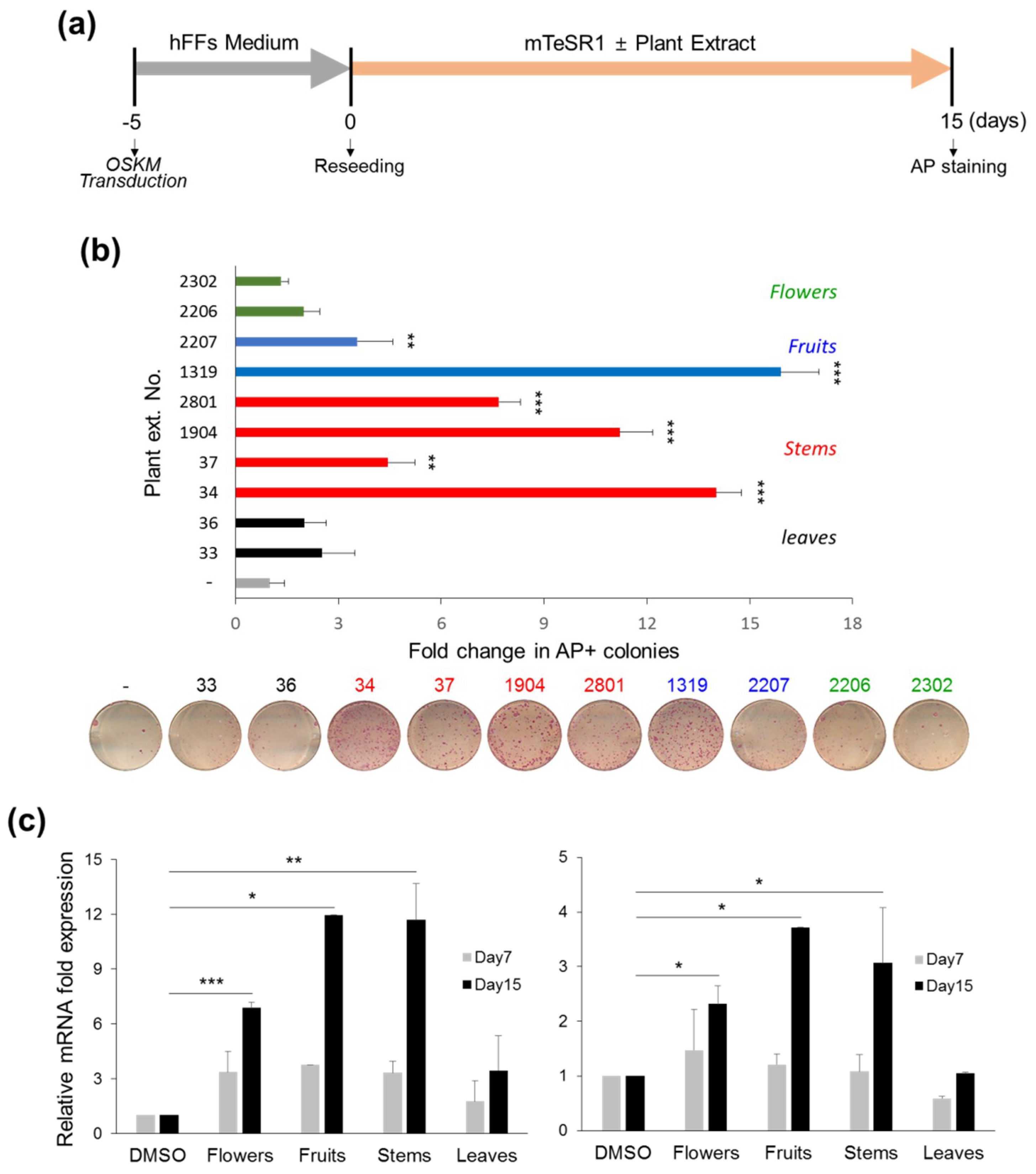
3.1.2. Stimulatory Effects of CJ Extracts on Human cell Reprogramming
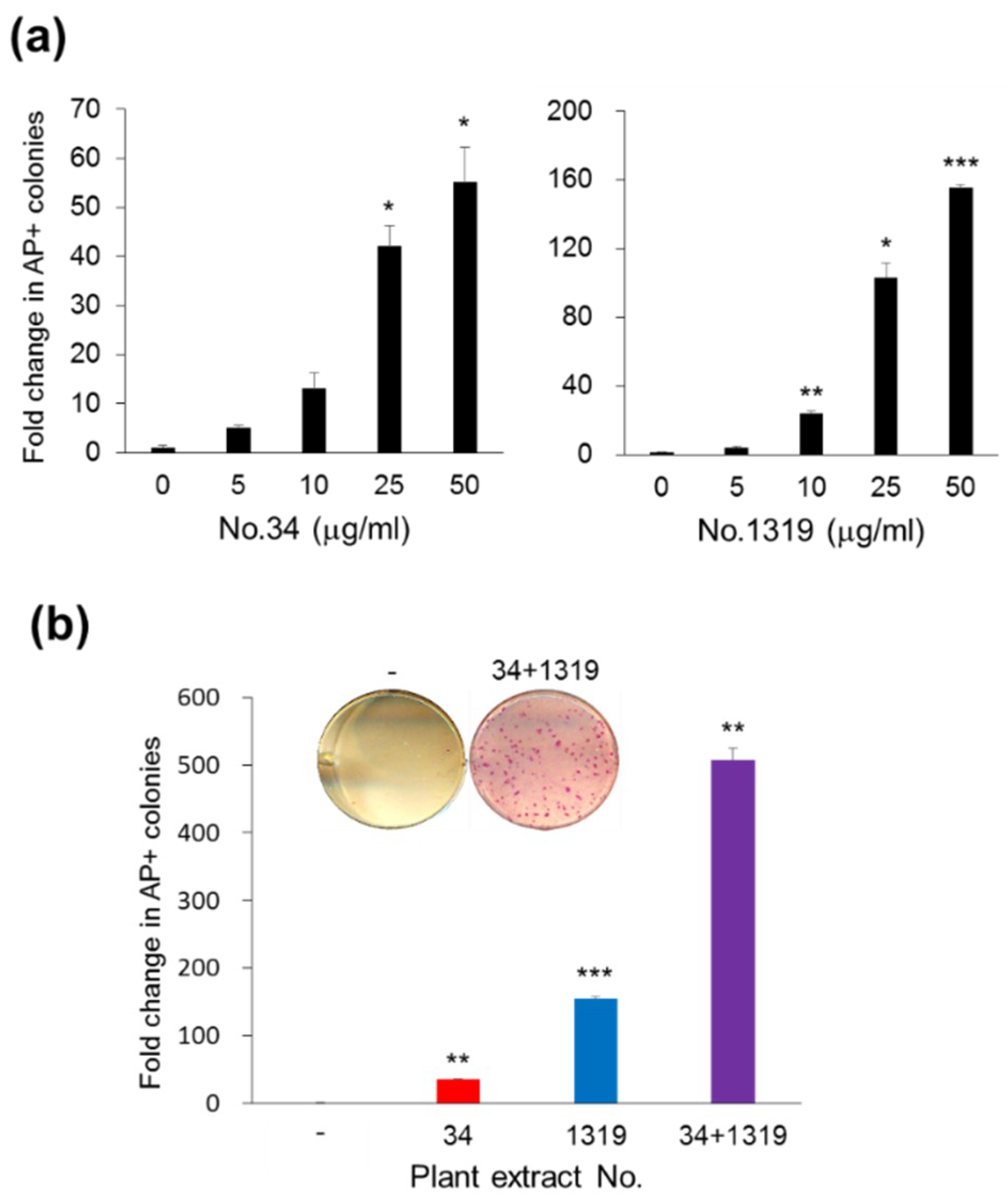
3.1.3. Synergistic Effects of CJ Extracts from Fruit and Stem on Human Cell Reprogramming
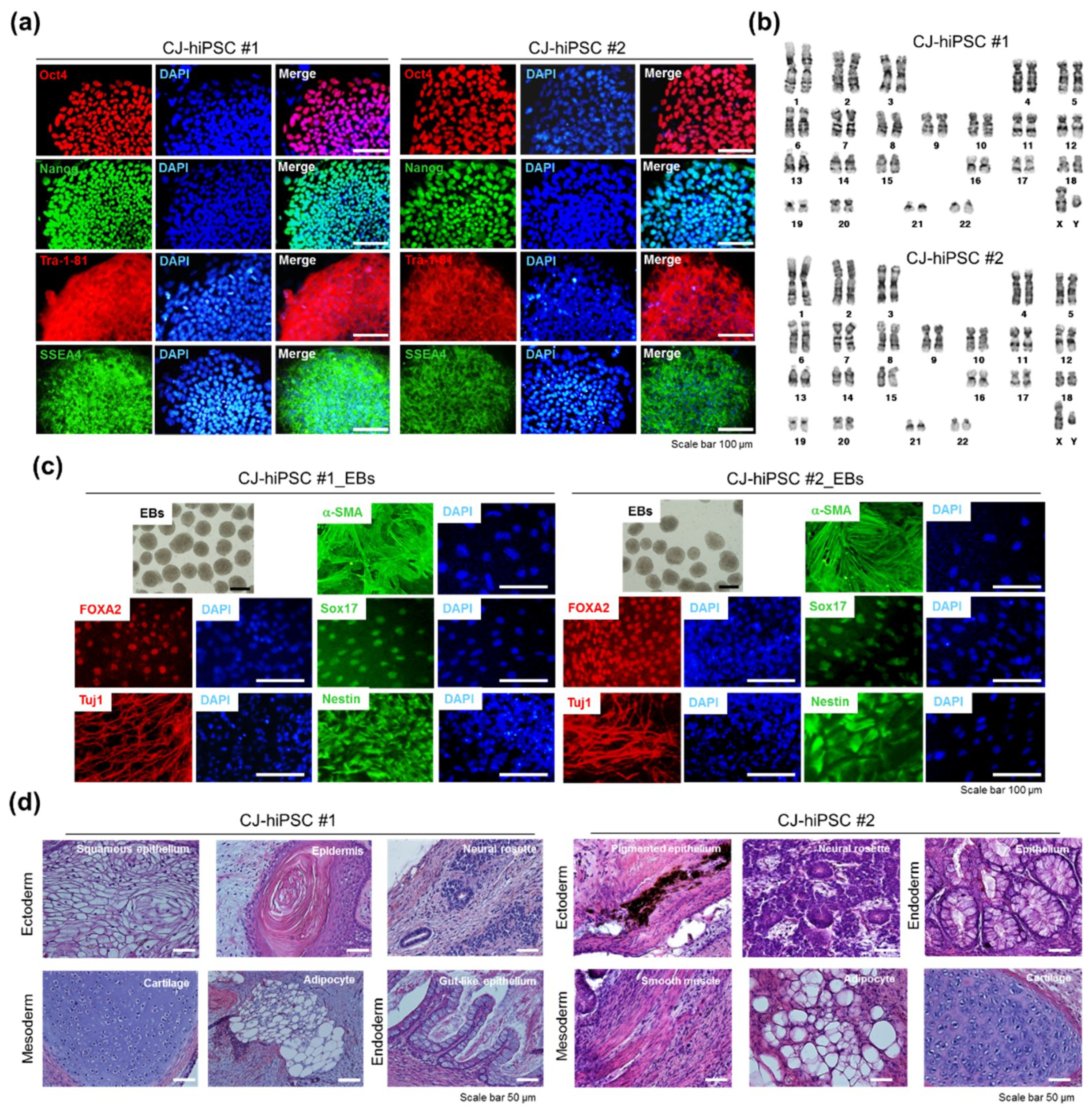
3.1.4. Characterization of the CJ-Extract-Induced hiPSC Pluripotency Ability
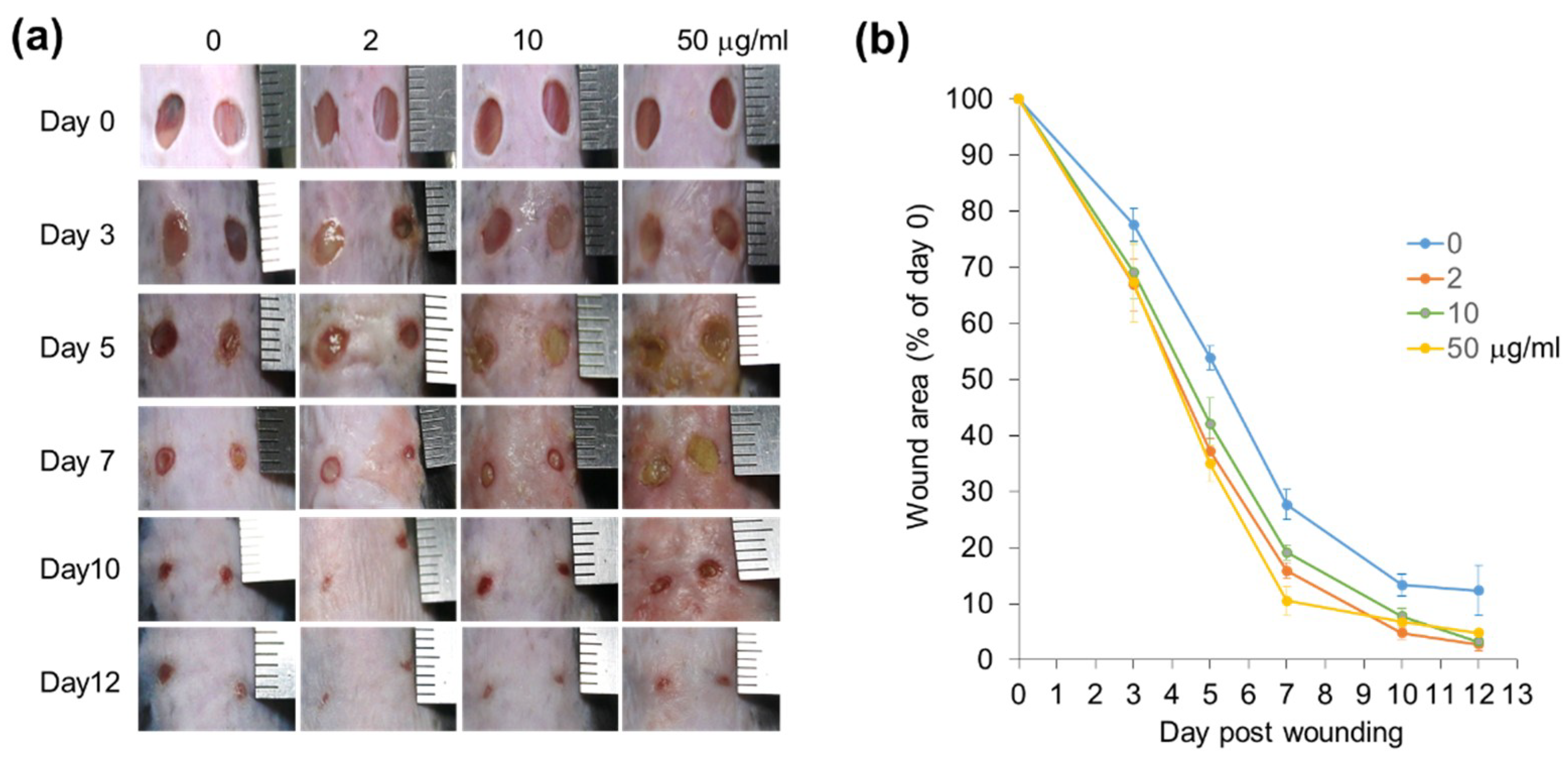
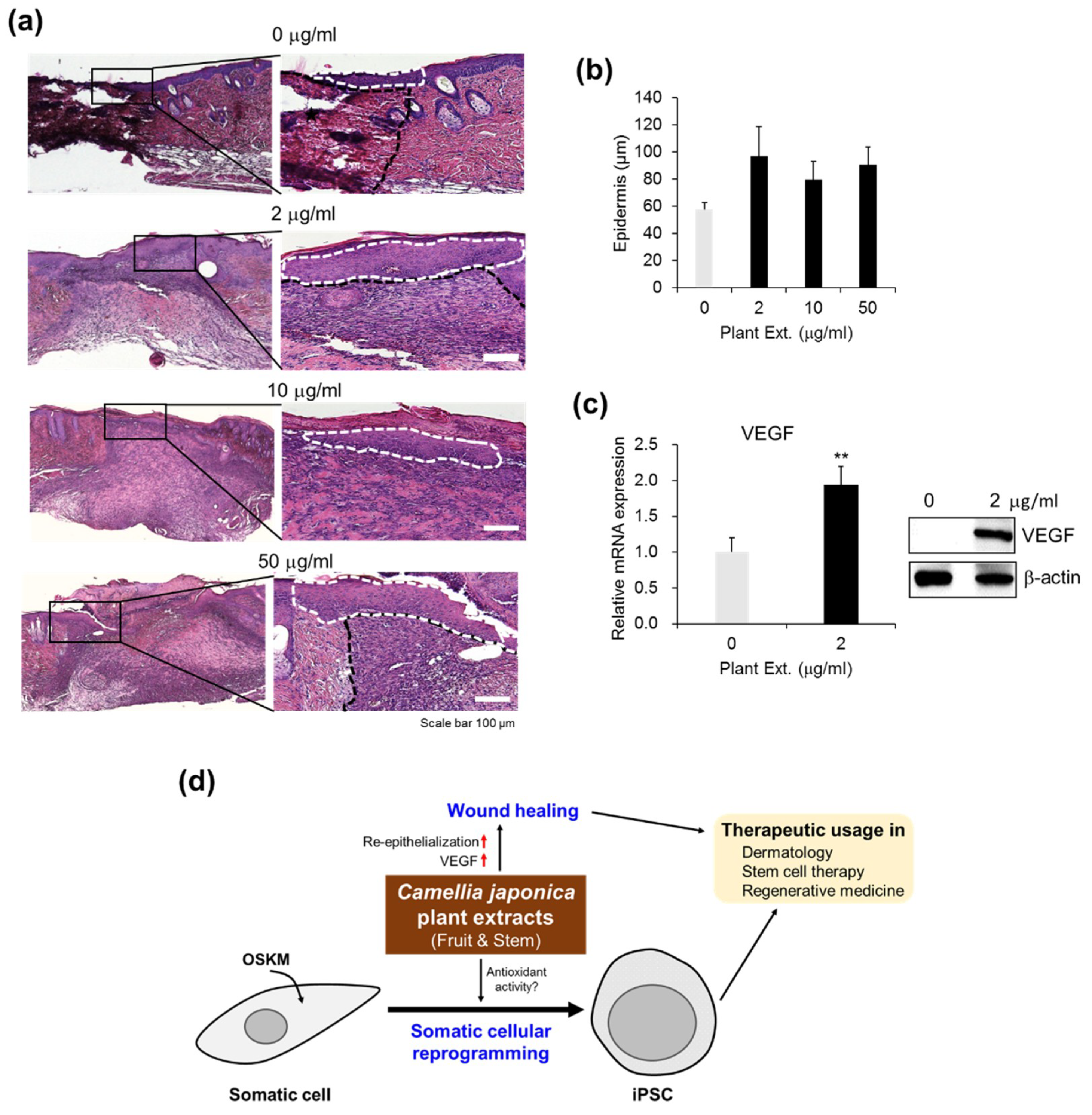
3.1.5. Positive Effects of CJ Extracts on Re-Epithelialization in Wound Healing
3.1.6. Upregulation of VEGF Expression by CJ Extracts Promotes Wound Healing
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Lup, S.D.; Tian, X.; Xu, J.; Perez-Perez, J.M. Wound signaling of regenerative cell reprogramming. Plant Sci. Int. J. Exp. Plant Biol. 2016, 250, 178–187. [Google Scholar] [CrossRef] [PubMed]
- Katsuyama, T.; Paro, R. Epigenetic reprogramming during tissue regeneration. FEBS Lett. 2011, 585, 1617–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosteiro, L.; Pantoja, C.; Alcazar, N.; Marion, R.M.; Chondronasiou, D.; Rovira, M.; Fernandez-Marcos, P.J.; Munoz-Martin, M.; Blanco-Aparicio, C.; Pastor, J.; et al. Tissue damage and senescence provide critical signals for cellular reprogramming in vivo. Science 2016, 354, aaf4445. [Google Scholar] [CrossRef] [PubMed]
- Jessen, K.R.; Mirsky, R.; Arthur-Farraj, P. The role of cell plasticity in tissue repair: Adaptive cellular reprogramming. Dev. Cell 2015, 34, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Pesaresi, M.; Bonilla-Pons, S.A.; Cosma, M.P. In vivo somatic cell reprogramming for tissue regeneration: The emerging role of the local microenvironment. Curr. Opin. Cell Biol. 2018, 55, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, antioxidants, and the degenerative diseases of aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef] [PubMed]
- Rohani, L.; Johnson, A.A.; Arnold, A.; Stolzing, A. The aging signature: A hallmark of induced pluripotent stem cells? Aging Cell 2014, 13, 2–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soria-Valles, C.; Lopez-Otin, C. Ipscs: On the road to reprogramming aging. Trend Mol. Med. 2016, 22, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Sharma, V.; Qi, S.; Guarch, M.E.; Zhao, P.; Luo, Z.; Fan, W.; Wang, Y.; Mbabaali, F.; Neculai, D. Antioxidant supplementation reduces genomic aberrations in human induced pluripotent stem cells. Stem Cell Rep. 2014, 2, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutta, D.; Heo, I.; Clevers, H. Disease modeling in stem cell-derived 3d organoid systems. Trend Mol. Med. 2017, 23, 393–410. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Kong, L.; Zhu, S. Reprogramming cell fates by small molecules. Protein Cell. 2017, 5, 328–348. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Schultz, P.G. Small molecules and future regenerative medicine. Curr. Top. Med. Chem. 2005, 5, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, W.; Laurent, T.; Ding, S. Small molecules, big roles—The chemical manipulation of stem cell fate and somatic cell reprogramming. J. Cell Sci. 2012, 125, 5609–5620. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.M.; Heo, D.R.; Lee, J.Y.; Seo, C.S.; Chung, S.K. High-efficiency generation of induced pluripotent stem cells from human foreskin fibroblast cells using the sagunja-tang herbal formula. BMC Complement. Altern. Med. 2017, 17, 529. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, J.; Kurpisz, M. Techniques of human embryonic stem cell and induced pluripotent stem cell derivation. Arch. Immunol. Ther. Exp. 2016, 64, 349–370. [Google Scholar] [CrossRef] [PubMed]
- Sasidharan, S.; Chen, Y.; Saravanan, D.; Sundram, K.; Latha, L.Y. Extraction, isolation and characterization of bioactive compounds from plants’ extracts. Afr. J. Tradit. Complement. Altern. Med. 2011, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cos, P.; Vlietinck, A.J.; Berghe, D.V.; Maes, L. Anti-infective potential of natural products: How to develop a stronger in vitro ‘proof-of-concept’. J. Ethnopharmacol. 2006, 106, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Duraipandiyan, V.; Ayyanar, M.; Ignacimuthu, S. Antimicrobial activity of some ethnomedicinal plants used by paliyar tribe from tamil nadu, india. BMC Complement. Altern. Med. 2006, 6, 35. [Google Scholar] [CrossRef] [PubMed]
- Farnsworth, N.R.; Akerele, O.; Bingel, A.S.; Soejarto, D.D.; Guo, Z. Medicinal plants in therapy. Bull. World Health Organ. 1985, 63, 965–981. [Google Scholar] [CrossRef]
- Efferth, T. Cancer therapy with natural products and medicinal plants. Planta Med. 2010, 76, 1035–1036. [Google Scholar] [CrossRef] [PubMed]
- Abu-Darwish, M.S.; Efferth, T. Medicinal plants from near east for cancer therapy. Front. Pharmacol. 2018, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Alviano, D.S.; Alviano, C.S. Plant extracts: Search for new alternatives to treat microbial diseases. Curr. Pharm. Biotechnol. 2009, 10, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Woo, Y.; Lee, H.; Jeong, Y.S.; Shin, G.Y.; Oh, J.G.; Kim, J.S.; Oh, J. Antioxidant potential of selected korean edible plant extracts. Biomed. Res. Int. 2017. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.; Jang, H.W.; Shibamoto, T. Antioxidant activities of extracts from teas prepared from medicinal plants, morus alba l., camellia sinensis l., and cudrania tricuspidata, and their volatile components. J. Agric. Food Chem. 2012, 60, 9097–9105. [Google Scholar] [CrossRef] [PubMed]
- Ding, A.J.; Zheng, S.Q.; Huang, X.B.; Xing, T.K.; Wu, G.S.; Sun, H.Y.; Qi, S.H.; Luo, H.R. Current perspective in the discovery of anti-aging agents from natural products. Nat. Prod. Bioprospect. 2017, 7, 335–404. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.Y.; Jiang, J.G.; Yang, L.; Wang, D.W.; Zhu, W. Anti-ageing active ingredients from herbs and nutraceuticals used in traditional Chinese medicine: Pharmacological mechanisms and implications for drug discovery. Br. J. Pharmacol. 2017, 174, 1395–1425. [Google Scholar] [CrossRef] [PubMed]
- Calixto, J.B.; Scheidt, C.; Otuki, M.; Santos, A.R. Biological activity of plant extracts: Novel analgesic drugs. Expert Opin. Emerg. Drug 2001, 6, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Budovsky, A.; Yarmolinsky, L.; Ben-Shabat, S. Effect of medicinal plants on wound healing. Wound Repair Regen. 2015, 23, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Thangapazham, R.L.; Sharad, S.; Maheshwari, R.K. Phytochemicals in wound healing. Adv. Wound Care 2016, 5, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, K.; Gordon, S.P.; Meyerowitz, E.M. Regeneration in plants and animals: Dedifferentiation, transdifferentiation, or just differentiation? Trend Cell Biol. 2011, 21, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Choi, J.-H.; Cui, L.; Li, Y.; Yang, J.M.; Yun, J.-J.; Jung, J.E.; Choi, W.; Yoon, K.C. Anti-inflammatory and antioxidative effects of Camellia japonica on human corneal epithelial cells and experimental dry eye: In vivo and in vitro study. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1196–1207. [Google Scholar] [CrossRef] [PubMed]
- Piao, M.J.; Yoo, E.S.; Koh, Y.S.; Kang, H.K.; Kim, J.; Kim, Y.J.; Kang, H.H.; Hyun, J.W. Antioxidant effects of the ethanol extract from flower of Camellia japonica via scavenging of reactive oxygen species and induction of antioxidant enzymes. Int. J. Mol. Sci. 2011, 12, 2618–2630. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Jung, E.; Shin, S.; Kim, M.; Kim, Y.S.; Lee, J.; Park, D. Anti-inflammatory activity of Camellia japonica oil. BMB Rep. 2012, 45, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Miura, D.; Kida, Y.; Nojima, H. Camellia oil and its distillate fractions effectively inhibit the spontaneous metastasis of mouse melanoma bl6 cells. FEBS Lett. 2007, 581, 2541–2548. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, J.W.; Ko, N.Y.; Mun, S.H.; Kim, D.K.; Kim, J.D.; Kim, H.S.; Lee, K.R.; Kim, Y.K.; Radinger, M.; et al. Camellia japonica suppresses immunoglobulin e-mediated allergic response by the inhibition of syk kinase activation in mast cells. Clin. Exp. Allergy 2008, 38, 794–804. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Davidson, P.M.; Chung, H.J. Antibacterial activity in extracts of Camellia japonica l. Petals and its application to a model food system. J. Food Prot. 2001, 64, 1255–1260. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Shim, B.; Yoon, J.; Lee, H.; Lee, H.; Yoo, S.; Wi, A.; Park, W.; Kim, H.; Kim, D. Vascular protective effect of an ethanol extract of Camellia japonica fruit: Endothelium-dependent relaxation of coronary artery and reduction of smooth muscle cell migration. Oxid. Med. Cell. Longev. 2015. [Google Scholar] [CrossRef]
- Lee, H.-H.; Paudel, K.R.; Jeong, J.; Wi, A.-J.; Park, W.-S.; Kim, D.-W.; Oak, M.-H. Antiatherogenic effect of Camellia japonica fruit extract in high fat diet-fed rats. Evid. Based Complement. Altern. Med. 2016. [Google Scholar] [CrossRef]
- Jung, E.; Lee, J.; Baek, J.; Jung, K.; Lee, J.; Huh, S.; Kim, S.; Koh, J.; Park, D. Effect of Camellia japonica oil on human type I procollagen production and skin barrier function. J. Ethnopharmacol. 2007, 112, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Akanda, M.R.; Park, B.-Y. Involvement of MAPK/NF-κB signal transduction pathways: Camellia japonica mitigates inflammation and gastric ulcer. Biomed. Pharmacother. 2017, 95, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Ahn, K. The worldwide trend of using botanical drugs and strategies for developing global drugs. BMB Rep. 2017, 50, 111–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.A.; Cheong, D.E.; Lim, H.D.; Kim, W.H.; Ham, M.H.; Oh, M.H.; Wu, Y.; Shin, H.J.; Kim, G.J. Antimicrobial and anti-biofilm activities of the methanol extracts of medicinal plants against dental pathogens streptococcus mutans and candida albicans. J. Microbiol. Biotechnol. 2017, 27, 1242–1248. [Google Scholar] [CrossRef] [PubMed]
- Cowan, C.A.; Klimanskaya, I.; McMahon, J.; Atienza, J.; Witmyer, J.; Zucker, J.P.; Wang, S.; Morton, C.C.; McMahon, A.P.; Powers, D. Derivation of embryonic stem-cell lines from human blastocysts. N. Engl. J. Med. 2004, 350, 1353–1356. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, H.K.; Rho, J.Y.; Han, Y.M.; Kim, J. The human OCT-4 isoforms differ in their ability to confer self-renewal. J. Biol. Chem. 2006, 281, 33554. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Xia, Y.; Son, M.Y.; Jin, G.; Seol, B.; Kim, M.J.; Son, M.J.; Do, M.; Lee, M.; Kim, D. A novel small molecule facilitates the reprogramming of human somatic cells into a pluripotent state and supports the maintenance of an undifferentiated state of human pluripotent stem cells. Angew. Chem. 2012, 124, 12677–12681. [Google Scholar] [CrossRef]
- Carey, B.; Markoulaki, S.; Beard, C.; Hanna, J.; Jaenisch, R. Single-gene transgenic mouse strains for reprogramming adult somatic cells. Nat. Method 2010, 7, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Lyssiotis, C.A.; Foreman, R.K.; Staerk, J.; Garcia, M.; Mathur, D.; Markoulaki, S.; Hanna, J.; Lairson, L.L.; Charette, B.D.; Bouchez, L.C.; et al. Reprogramming of murine fibroblasts to induced pluripotent stem cells with chemical complementation of klf4. Proc. Natl. Acad. Sci. USA 2009, 106, 8912–8917. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Ueno, M.; Kamiya, D.; Nishiyama, A.; Matsumura, M.; Wataya, T.; Takahashi, J.B.; Nishikawa, S.; Nishikawa, S.; Muguruma, K.; et al. A rock inhibitor permits survival of dissociated human embryonic stem cells. Nat. Biotechnol. 2007, 25, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Goldblum, O.M.; Alvarez, O.M.; Mertz, P.M.; Eaglstein, W.H. Dimethyl sulfoxide (DMSO) does not affect epidermal wound healing. Proc. Soc. Exp. Biol. Med. 1983, 172, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Martin, P. Wound healing—Aiming for perfect skin regeneration. Science 1997, 276, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Hoeben, A.; Landuyt, B.; Highley, M.S.; Wildiers, H.; Van Oosterom, A.T.; De Bruijn, E.A. Vascular endothelial growth factor and angiogenesis. Pharmacol. Rev. 2004, 56, 549–580. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.E.; Wilgus, T.A. Vascular endothelial growth factor and angiogenesis in the regulation of cutaneous wound repair. Adv. Wound Care 2014, 3, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Bao, P.; Kodra, A.; Tomic-Canic, M.; Golinko, M.S.; Ehrlich, H.P.; Brem, H. The role of vascular endothelial growth factor in wound healing. J. Surg. Res. 2009, 153, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Banito, A.; Rashid, S.T.; Acosta, J.C.; Li, S.; Pereira, C.F.; Geti, I.; Pinho, S.; Silva, J.C.; Azuara, V.; Walsh, M. Senescence impairs successful reprogramming to pluripotent stem cells. Genes Dev. 2009, 23, 2134–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteban, M.A.; Wang, T.; Qin, B.; Yang, J.; Qin, D.; Cai, J.; Li, W.; Weng, Z.; Chen, J.; Ni, S. Vitamin C enhances the generation of mouse and human induced pluripotent stem cells. Cell Stem Cell 2010, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Bigarella, C.L.; Liang, R.; Ghaffari, S. Stem cells and the impact of ROS signaling. Development 2014, 141, 4206–4218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohyeldin, A.; Garzón-Muvdi, T.; Quiñones-Hinojosa, A. Oxygen in stem cell biology: A critical component of the stem cell niche. Cell Stem Cell 2010, 7, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Mah, N.; Wang, Y.; Liao, M.-C.; Prigione, A.; Jozefczuk, J.; Lichtner, B.; Wolfrum, K.; Haltmeier, M.; Flöttmann, M.; Schaefer, M. Molecular insights into reprogramming-initiation events mediated by the OSKM gene regulatory network. PLoS ONE 2011, 6, e24351. [Google Scholar] [CrossRef] [PubMed]
- Chiou, S.-S.; Wang, S.S.-W.; Wu, D.-C.; Lin, Y.-C.; Kao, L.-P.; Kuo, K.-K.; Wu, C.-C.; Chai, C.-Y.; Lin, C.-L.S.; Lee, C.-Y. Control of oxidative stress and generation of induced pluripotent stem cell-like cells by Jun Dimerization protein 2. Cancers 2013, 5, 959–984. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, L.; Neel, B.G.; Aifantis, I. Vitamin c in stem cell reprogramming and cancer. Trend Cell Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, C.; Vissers, M.C. Ascorbate as a co-factor for fe- and 2-oxoglutarate dependent dioxygenases: Physiological activity in tumor growth and progression. Front. Oncol. 2014, 4, 359. [Google Scholar] [CrossRef] [PubMed]
- Fronza, M.; Heinzmann, B.; Hamburger, M.; Laufer, S.; Merfort, I. Determination of the wound healing effect of calendula extracts using the scratch assay with 3T3 fibroblasts. J. Ethnopharmacol. 2009, 126, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.N.; Sharma, G.; Yang, J.-L.; Choi, H.S.; Lim, S.-I.; Kang, K.W.; Oh, W.K. Oleanane triterpenes as protein tyrosine phosphatase 1B (PTP1B) inhibitors from Camellia japonica. Phytochemistry 2014, 103, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Thao, N.T.P.; Hung, T.M.; Lee, M.K.; Kim, J.C.; Min, B.S.; Bae, K. Triterpenoids from Camellia japonica and their cytotoxic activity. Chem. Pharm. Bull. 2010, 58, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Thao, N.T.; Hung, T.M.; Cuong, T.D.; Kim, J.C.; Kim, E.H.; Jin, S.E.; Na, M.; Lee, Y.M.; Kim, Y.H.; Choi, J.S.; et al. 28-nor-oleanane-type triterpene saponins from Camellia japonica and their inhibitory activity on LPS-induced no production in macrophage raw264.7 cells. Bioorg. Med. Chem. Lett. 2010, 20, 7435–7439. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeon, H.; Kim, J.Y.; Choi, J.; Han, E.; Song, C.-L.; Lee, J.; Cho, Y.S. Effects of the Extracts from Fruit and Stem of Camellia japonica on Induced Pluripotency and Wound Healing. J. Clin. Med. 2018, 7, 449. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm7110449
Jeon H, Kim JY, Choi J, Han E, Song C-L, Lee J, Cho YS. Effects of the Extracts from Fruit and Stem of Camellia japonica on Induced Pluripotency and Wound Healing. Journal of Clinical Medicine. 2018; 7(11):449. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm7110449
Chicago/Turabian StyleJeon, Hyejin, Jae Yun Kim, Jung‐Kyun Choi, Enna Han, Cho-Lok Song, Jungwoon Lee, and Yee Sook Cho. 2018. "Effects of the Extracts from Fruit and Stem of Camellia japonica on Induced Pluripotency and Wound Healing" Journal of Clinical Medicine 7, no. 11: 449. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm7110449