Potential Therapeutic Targets of Quercetin and Its Derivatives: Its Role in the Therapy of Cognitive Impairment

 ,
,  ,
, 

Abstract
:1. Introduction
2. Overview of Quercetin
3. Potential Therapeutic Targets of Quercetin
3.1. Voltage-Gated Ion Channels
3.2. Neuroreceptors
3.2.1. Dopamine Receptors
3.2.2. Glutaminergic Receptors
3.2.3. Acetylcholine Receptors
3.2.4. Serotonergic Receptors
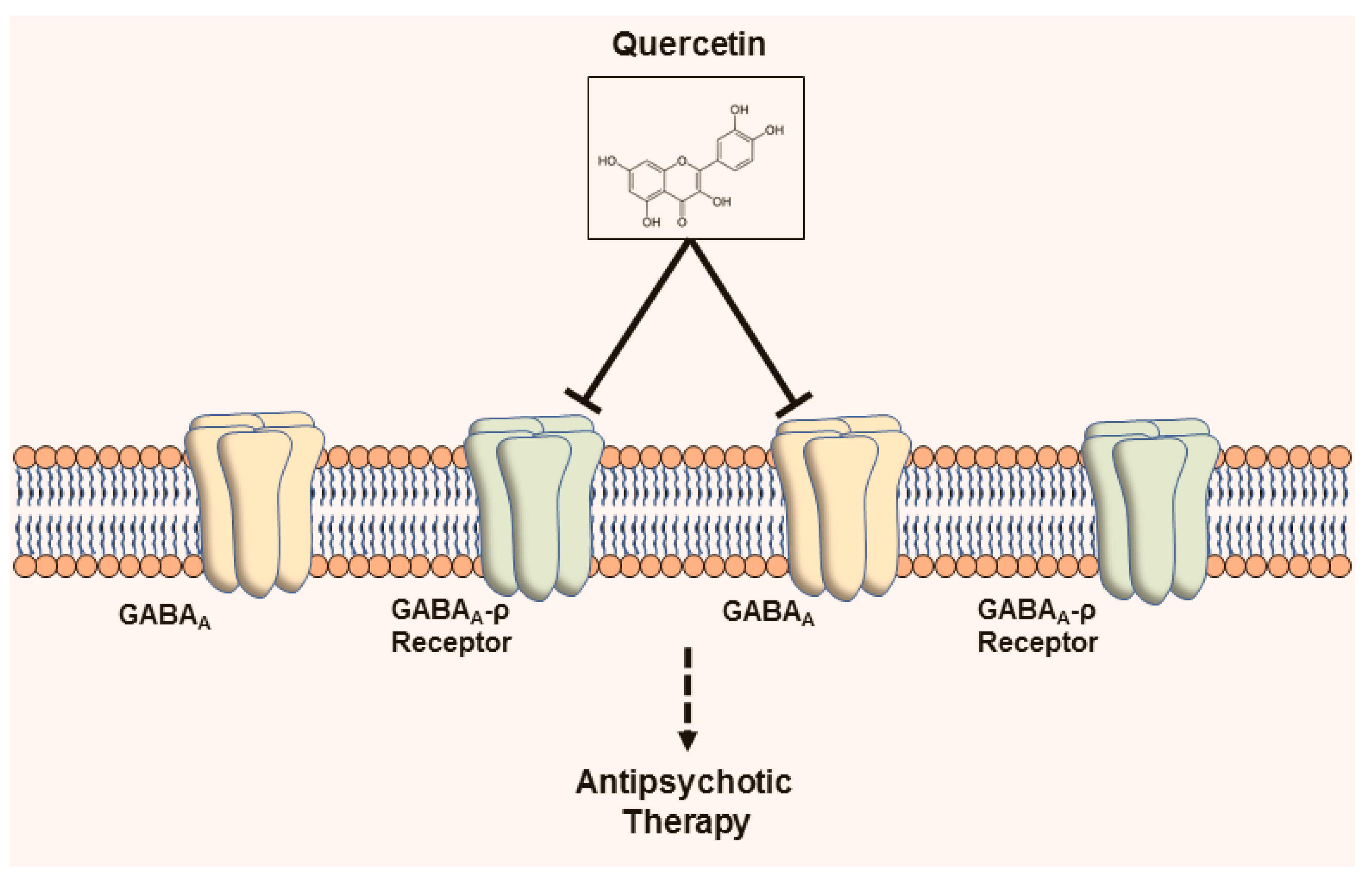
3.2.5. Gamma-aminobutyric acid-ergic Receptors
3.2.6. Glycinergic Receptors
3.3. Miscellaneous Targets
3.3.1. Toll-Like Receptors and Cytokine Receptors
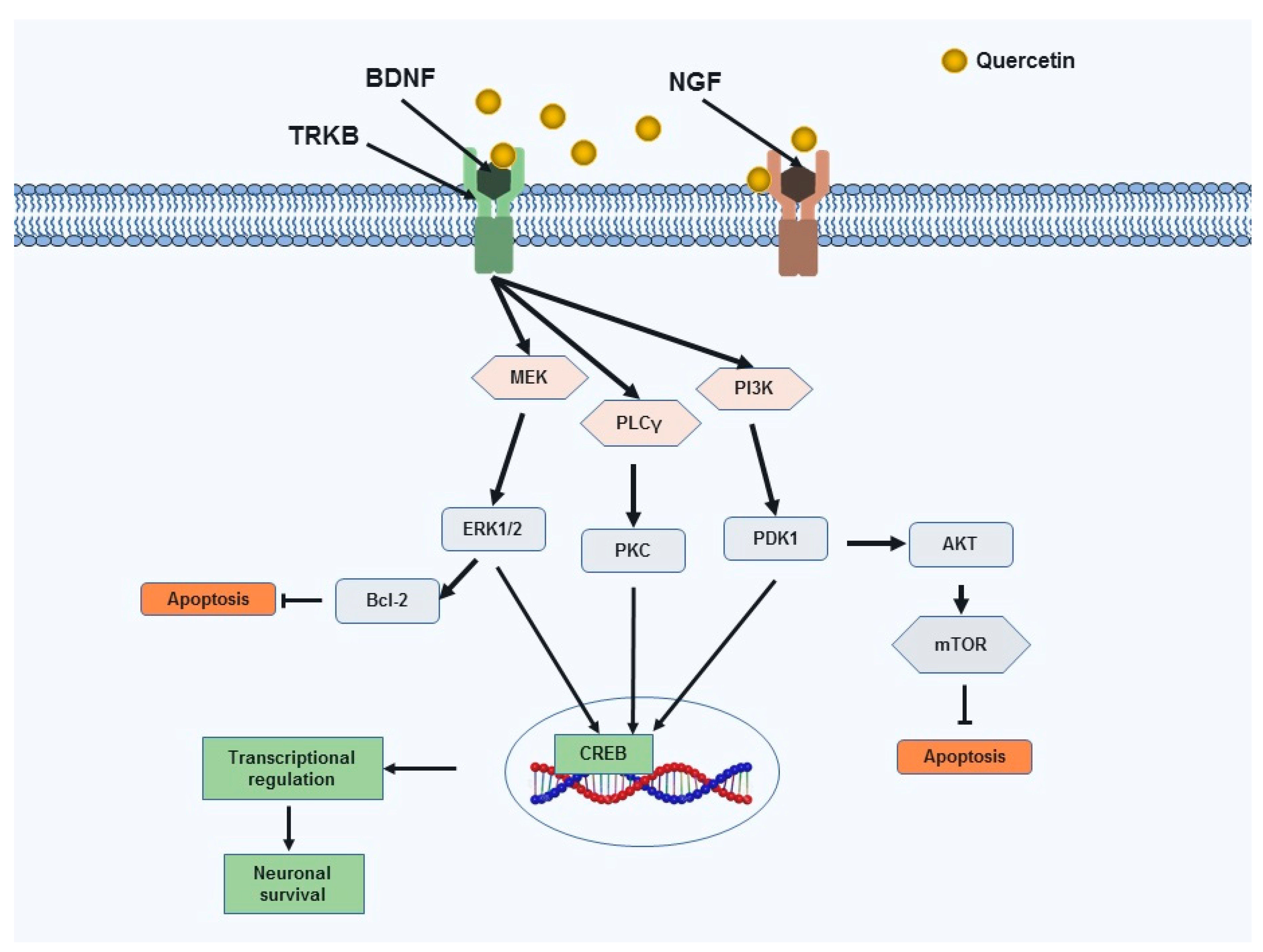
3.3.2. Neurotrophic Factors
3.3.3. Apolipoprotein E
3.3.4. Nuclear Factor Erythroid 2-Related Factor 2-Antioxidant Responsive Element
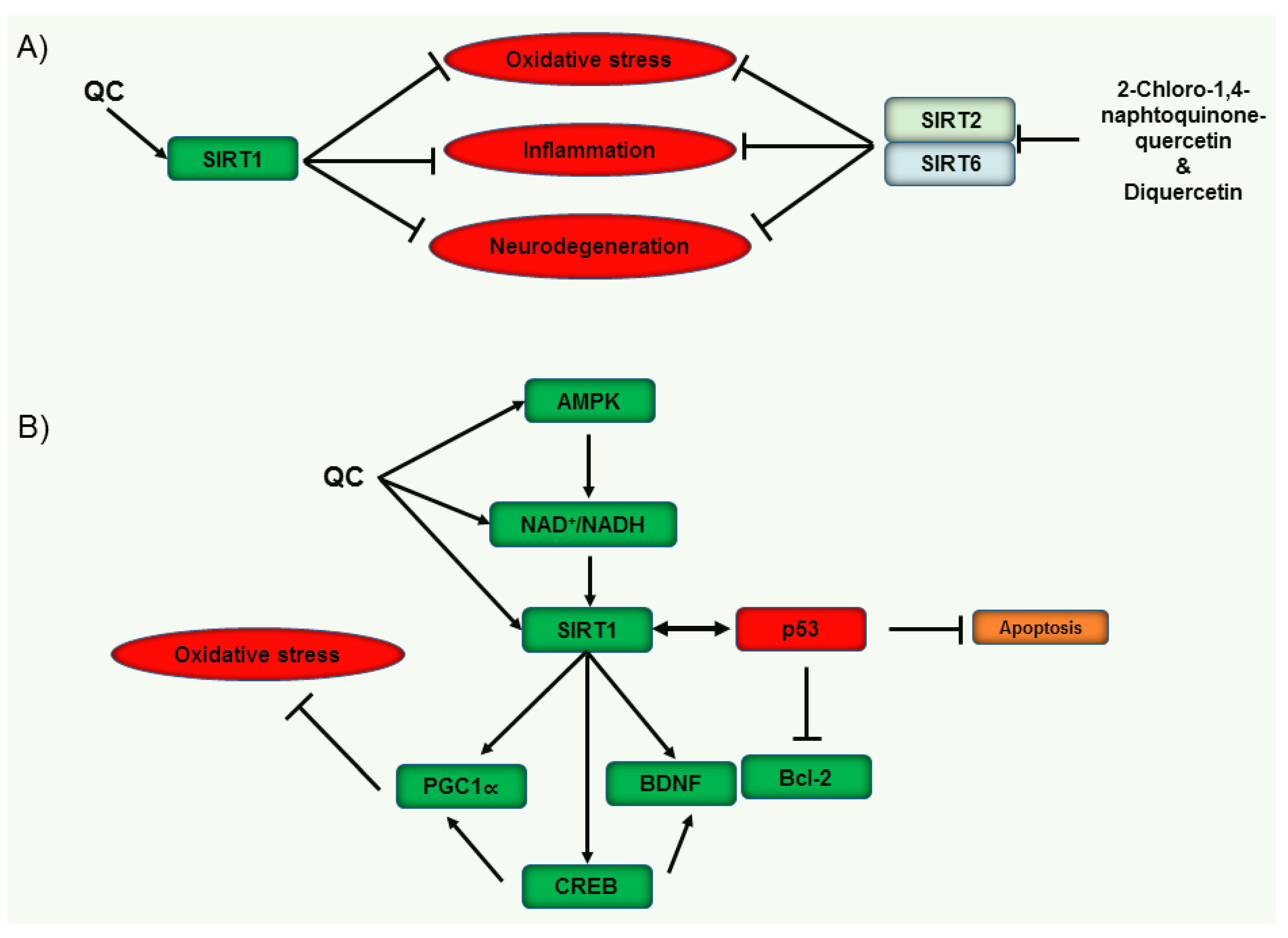
3.3.5. Sirtuins
4. Closing Remarks
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 6-OHDA | 6-hydroxydopamine |
| AD | Alzheimer’s disease |
| Akt | Protein Kinase B |
| AMPA | α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid |
| APOE | Apolipoprotein E |
| Aβ | Amyloid beta |
| BBB | Blood-brain barrier |
| Bcl-2 | B-cell lymphoma 2 |
| BDNF | Brain-derived neurotrophic factor |
| CD | Cluster of differentiation |
| CREB | Cyclic AMP response element binding protein |
| ERK | Extracellular signal-regulated kinase 1/2 |
| GABA | Gamma-aminobutyric acid |
| IACh | Inward currents |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| Keap1 | Kelch-like ECH-associated protein 1 |
| MAPK | Mitogen-activated protein kinase |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| mTOR | Mammalian target of rapamycin |
| NDDs | Neurodegenerative diseases |
| NF-κB | Nuclear factor-kappa B |
| NGF | Nerve growth factor |
| NKCC1 | Na-K-Cl cotransporter |
| NMDA | N-methyl-D-aspartate |
| NO | Nitric oxide |
| NQO1 | NAD(P)H quinone oxidoreducase-1 |
| Nrf2-ARE | Nuclear factor erythroid 2-related factor 2-antioxidant responsive element |
| PD | Parkinson’s disease |
| PDK1 | 3-phosphoinositide-dependent protein kinase 1 |
| PI3K | Phosphatidylinositol 3-kinase |
| PKC | Protein kinase C |
| PLCγ | Phospholipase C gamma |
| QC | Quercetin |
| SIRT | Sirtuin |
| TLR | Toll-like receptor |
| TrkB | Tropomyosin receptor kinase B |
| α7 nAChR | α7 nicotinic acetylcholine receptor |
| HO-1 | Heme-oxygenase-1 |
References
- Jakaria, M.; Haque, M.E.; Kim, J.; Cho, D.-Y.; Kim, I.-S.; Choi, D.-K. Active ginseng components in cognitive impairment: Therapeutic potential and prospects for delivery and clinical study. Oncotarget 2018, 9, 33601–33620. [Google Scholar] [CrossRef] [PubMed]
- Jakaria, M.; Kim, J.; Karthivashan, G.; Park, S.-Y.; Ganesan, P.; Choi, D.-K. Emerging signals modulating potential of ginseng and its active compounds focusing on neurodegenerative diseases. J. Ginseng Res. 2019, 43, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Jakaria, M.; Park, S.-Y.; Haque, M.E.; Karthivashan, G.; Kim, I.-S.; Ganesan, P.; Choi, D.-K. Neurotoxic Agent-Induced Injury in Neurodegenerative Disease Model: Focus on Involvement of Glutamate Receptors. Front. Mol. Neurosci. 2018, 11, 307. [Google Scholar] [CrossRef] [PubMed]
- Amieva, H.; Meillon, C.; Helmer, C.; Barberger-Gateau, P.; Dartigues, J.F. Ginkgo biloba extract and long-term cognitive decline: A 20-year follow-up population-based study. PLoS ONE 2013, 8, e52755. [Google Scholar] [CrossRef]
- Jakaria, M.; Cho, D.Y.; EzazulHaque, M.; Karthivashan, G.; Kim, I.S.; Ganesan, P.; Choi, D.K. Neuropharmacological Potential and Delivery Prospects of Thymoquinone for Neurological Disorders. Oxid. Med. Cell. Longev. 2018, 2018, 1209801. [Google Scholar] [CrossRef]
- Uddin, M.S.; Al Mamun, A.; Kabir, M.T.; Jakaria, M.; Mathew, B.; Barreto, G.E.; Ashraf, G.M. Nootropic and Anti-Alzheimer’s Actions of Medicinal Plants: Molecular Insight into Therapeutic Potential to Alleviate Alzheimer’s Neuropathology. Mol. Neurobiol. 2018, 56, 4925–4944. [Google Scholar] [CrossRef]
- Jakaria, M.; Haque, M.E.; Cho, D.Y.; Azam, S.; Kim, I.S.; Choi, D.K. Molecular Insights into NR4A2(Nurr1): An Emerging Target for Neuroprotective Therapy Against Neuroinflammation and Neuronal Cell Death. Mol. Neurobiol. 2019, 56, 5799–5814. [Google Scholar] [CrossRef]
- Ay, M.; Luo, J.; Langley, M.; Jin, H.; Anantharam, V.; Kanthasamy, A.; Kanthasamy, A.G. Molecular mechanisms underlying protective effects of quercetin against mitochondrial dysfunction and progressive dopaminergic neurodegeneration in cell culture and MitoPark transgenic mouse models of Parkinson’s Disease. J. Neurochem. 2017, 141, 766–782. [Google Scholar] [CrossRef]
- Dajas, F. Life or death: Neuroprotective and anticancer effects of quercetin. J. Ethnopharmacol. 2012, 143, 383–396. [Google Scholar] [CrossRef]
- Suganthy, N.; Devi, K.P.; Nabavi, S.F.; Braidy, N.; Nabavi, S.M. Bioactive effects of quercetin in the central nervous system: Focusing on the mechanisms of actions. Biomed. Pharmacother. = Biomed. Pharmacother. 2016, 84, 892–908. [Google Scholar] [CrossRef]
- Tinay, I.; Sener, T.E.; Cevik, O.; Cadirci, S.; Toklu, H.; Cetinel, S.; Sener, G.; Tarcan, T. Antioxidant Agent Quercetin Prevents Impairment of Bladder Tissue Contractility and Apoptosis in a Rat Model of Ischemia/Reperfusion Injury. Lower Urin. Tract Symptoms 2017, 9, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Bondonno, N.; Bondonno, C.; Hodgson, J.; Ward, N.C.; Croft, K.D. The Efficacy of Quercetin in Cardiovascular Health. Curr. Nutr. Rep. 2015, 4, 290–303. [Google Scholar] [CrossRef]
- Kosari-Nasab, M.; Shokouhi, G.; Ghorbanihaghjo, A.; Mesgari-Abbasi, M.; Salari, A.A. Quercetin mitigates anxiety-like behavior and normalizes hypothalamus–pituitary–adrenal axis function in a mouse model of mild traumatic brain injury. Behav. Pharmacol. 2019, 30, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G.; Garrick, J.M.; Roquè, P.J.; Pellacani, C. Mechanisms of Neuroprotection by Quercetin: Counteracting Oxidative Stress and More. Oxid. Med. Cell. Longev. 2016, 2016, 2986796. [Google Scholar] [CrossRef]
- Trippier, P.C.; Jansen Labby, K.; Hawker, D.D.; Mataka, J.J.; Silverman, R.B. Target- and Mechanism-Based Therapeutics for Neurodegenerative Diseases: Strength in Numbers. J. Med. Chem. 2013, 56, 3121–3147. [Google Scholar] [CrossRef] [Green Version]
- RusznyÁK, S.T.; Szent-GyÖRgyi, A. Vitamin P: Flavonols as Vitamins. Nature 1936, 138, 27. [Google Scholar] [CrossRef]
- Sampson, L.; Rimm, E.; Hollman, P.C.; de Vries, J.H.; Katan, M.B. Flavonol and flavone intakes in US health professionals. J. Am. Diet. Assoc. 2002, 102, 1414–1420. [Google Scholar] [CrossRef]
- Nishimuro, H.; Ohnishi, H.; Sato, M.; Ohnishi-Kameyama, M.; Matsunaga, I.; Naito, S.; Ippoushi, K.; Oike, H.; Nagata, T.; Akasaka, H.; et al. Estimated daily intake and seasonal food sources of quercetin in Japan. Nutrients 2015, 7, 2345–2358. [Google Scholar] [CrossRef]
- Jakaria, M.; Azam, S.; Cho, D.Y.; Haque, M.E.; Kim, I.S.; Choi, D.K. The Methanol Extract of Allium cepa L. Protects Inflammatory Markers in LPS-Induced BV-2 Microglial Cells and Upregulates the Antiapoptotic Gene and Antioxidant Enzymes in N27-A Cells. Antioxidants 2019, 8, 348. https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/31480531. [CrossRef]
- Harborne, J.B. Flavonoids in the environment: Structure-activity relationships. Prog. Clin. Biol. Res. 1988, 280, 17–27. [Google Scholar]
- Williams, C.A.; Grayer, R.J. Anthocyanins and other flavonoids. Nat. Prod. Rep. 2004, 21, 539–573. [Google Scholar] [CrossRef] [PubMed]
- Wiczkowski, W.A.; Piskuła, M. Food flavonoids. Pol. J. Food Nutr. Sci. 2004, 13, 101–114. [Google Scholar]
- Chang, Q.; Wong, Y.-S. Identification of Flavonoids in Hakmeitau Beans (Vigna sinensis) by High-Performance Liquid Chromatography− Electrospray Mass Spectrometry (LC-ESI/MS). J. Agric. Food Chem. 2004, 52, 6694–6699. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Foo, L.Y. Polyphenolics of Salvia—A review. Phytochemistry 2002, 59, 117–140. [Google Scholar] [CrossRef]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Hollman, P.; Katan, M. Absorption, metabolism and health effects of dietary flavonoids in man. Biomed. Pharmacother. 1997, 51, 305–310. [Google Scholar] [CrossRef]
- Mariani, C.; Braca, A.; Vitalini, S.; De Tommasi, N.; Visioli, F.; Fico, G. Flavonoid characterization and in vitro antioxidant activity of Aconitum anthoraL. (Ranunculaceae). Phytochemistry 2008, 69, 1220–1226. [Google Scholar] [CrossRef]
- Materska, M.; Perucka, I. Antioxidant activity of the main phenolic compounds isolated from hot pepper fruit (Capsicum annuum L.). J. Agric. Food Chem. 2005, 53, 1750–1756. [Google Scholar] [CrossRef]
- Janisch, K.M.; Williamson, G.; Needs, P.; Plumb, G.W. Properties of quercetin conjugates: Modulation of LDL oxidation and binding to human serum albumin. Free Radic. Res. 2004, 38, 877–884. [Google Scholar] [CrossRef]
- Murota, K.; Terao, J. Antioxidative flavonoid quercetin: Implication of its intestinal absorption and metabolism. Arch. Biochem. Biophys. 2003, 417, 12–17. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed]
- Wittig, J.; Herderich, M.; Graefe, E.U.; Veit, M. Identification of quercetin glucuronides in human plasma by high-performance liquid chromatography-tandem mass spectrometry. J. Chromatography. B Biomed. Sci. Appl. 2001, 753, 237–243. [Google Scholar] [CrossRef]
- Azuma, K.; Ippoushi, K.; Ito, H.; Higashio, H.; Terao, J. Combination of lipids and emulsifiers enhances the absorption of orally administered quercetin in rats. J. Agric. Food Chem. 2002, 50, 1706–1712. [Google Scholar] [CrossRef] [PubMed]
- Mullen, W.; Edwards, C.A.; Crozier, A. Absorption, excretion and metabolite profiling of methyl-, glucuronyl-, glucosyl- and sulpho-conjugates of quercetin in human plasma and urine after ingestion of onions. Br. J. Nutr. 2006, 96, 107–116. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef]
- DuPont, M.S.; Mondin, Z.; Williamson, G.; Price, K.R. Effect of variety, processing, and storage on the flavonoid glycoside content and composition of lettuce and endive. J. Agric. Food Chem. 2000, 48, 3957–3964. [Google Scholar] [CrossRef]
- Harwood, M.; Danielewska-Nikiel, B.; Borzelleca, J.F.; Flamm, G.W.; Williams, G.M.; Lines, T.C. A critical review of the data related to the safety of quercetin and lack of evidence of in vivo toxicity, including lack of genotoxic/carcinogenic properties. Food Chem. Toxicol. 2007, 45, 2179–2205. https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pubmed/17698276. [CrossRef]
- Knab, A.M.; Shanely, R.A.; Henson, D.A.; Jin, F.; Heinz, S.A.; Austin, M.D.; Nieman, D.C. Influence of quercetin supplementation on disease risk factors in community-dwelling adults. J. Am. Diet. Assoc. 2011, 111, 542–549. [Google Scholar] [CrossRef]
- Choi, J.S.; Piao, Y.J.; Kang, K.W. Effects of quercetin on the bioavailability of doxorubicin in rats: Role of CYP3A4 and P-gp inhibition by quercetin. Arch. Pharm. Res. 2011, 34, 607–613. [Google Scholar] [CrossRef]
- Turnbull, F. Effects of different blood-pressure-lowering regimens on major cardiovascular events: Results of prospectively-designed overviews of randomised trials. Lancet (London, England) 2003, 362, 1527–1535. [Google Scholar]
- Kawabata, K.; Mukai, R.; Ishisaka, A. Quercetin and related polyphenols: New insights and implications for their bioactivity and bioavailability. Food Funct. 2015, 6, 1399–1417. [Google Scholar] [CrossRef] [PubMed]
- Larson, A.J.; Symons, J.D.; Jalili, T. Quercetin: A Treatment for Hypertension?-A Review of Efficacy and Mechanisms. Pharmaceuticals 2010, 3, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Mrvová, N.; Škandík, M.; Kuniaková, M.; Račková, L. Modulation of BV-2 microglia functions by novel quercetin pivaloyl ester. Neurochem. Int. 2015, 90, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Ishisaka, A.; Ichikawa, S.; Sakakibara, H.; Piskula, M.K.; Nakamura, T.; Kato, Y.; Ito, M.; Miyamoto, K.-I.; Tsuji, A.; Kawai, Y.; et al. Accumulation of orally administered quercetin in brain tissue and its antioxidative effects in rats. Free Radic. Biol. Med. 2011, 51, 1329–1336. [Google Scholar] [CrossRef]
- Berardini, N.; Fezer, R.; Conrad, J.; Beifuss, U.; Carle, R.; Schieber, A.J.J.O.A.; Chemistry, F. Screening of mango (Mangiferaindica L.) cultivars for their contents of flavonol O-and xanthone C-glycosides, anthocyanins, and pectin. Agric. Food Chem. 2005, 53, 1563–1570. [Google Scholar] [CrossRef]
- Nemeth, K.; Piskula, M.J.C.R.I.F.S. Food content, processing, absorption and metabolism of onion flavonoids. Nutrition 2007, 47, 397–409. [Google Scholar] [CrossRef]
- Keddy, P.G.W.; Dunlop, K.; Warford, J.; Samson, M.L.; Jones, Q.R.D.; Rupasinghe, H.P.V.; Robertson, G.S. Neuroprotective and anti-inflammatory effects of the flavonoid-enriched fraction AF4 in a mouse model of hypoxic-ischemic brain injury. PLoS ONE 2012, 7, 51324. [Google Scholar] [CrossRef]
- Slimestad, R.; Verheul, M.J.J.J.O.A.; Chemistry, F. Seasonal variations in the level of plant constituents in greenhouse production of cherry tomatoes.). Agric. Food Chem. 2005, 53, 3114–3119. [Google Scholar] [CrossRef]
- Uthra, C.; Shrivastava, S.; Jaswal, A.; Sinha, N.; Reshi, M.S.; Shukla, S. Therapeutic potential of quercetin against acrylamide induced toxicity in rats. Biomed. Pharmacother. 2017, 86, 705–714. [Google Scholar] [CrossRef]
- Ferri, P.; Angelino, D.; Gennari, L.; Benedetti, S.; Ambrogini, P.; Del Grande, P.; Ninfali, P. Enhancement of flavonoid ability to cross the blood-brain barrier of rats by co-administration with alpha-tocopherol. Food Funct. 2015, 6, 394–400. [Google Scholar] [CrossRef]
- Flamini, G.; Antognoli, E.; Morelli, I.J.P. Two flavonoids and other compounds from the aerial parts of Centaurea bracteata from Italy. Phytochemistry 2001, 57, 559–564. [Google Scholar] [CrossRef]
- Li, J.; Mottamal, M.; Li, H.; Liu, K.; Zhu, F.; Cho, Y.-Y.; Sosa, C.P.; Zhou, K.; Bowden, G.T.; Bode, A.M.; et al. Quercetin-3-methyl ether suppresses proliferation of mouse epidermal JB6 P+ cells by targeting ERKs. Carcinogenesis 2011, 33, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Bhattacharya, P.; Paul, S.; Patnaik, R. Rhamnetin Attenuates Oxidative Stress and Matrix Metalloproteinase in Animal Model of Ischemia/Reperfusion: A Possible Antioxidant Therapy in Stroke. Am. J. Neuroprot. Neuroregener. 2013, 5, 1–7. [Google Scholar] [CrossRef]
- Yao, L.; Datta, N.; Tomás-Barberán, F.A.; Ferreres, F.; Martos, I.; Singanusong, R.J.F.C. Flavonoids, phenolic acids and abscisic acid in Australian and New Zealand Leptospermum honeys. Food Chem. 2003, 81, 159–168. [Google Scholar] [CrossRef]
- Rangel-Ordonez, L.; Noldner, M.; Schubert-Zsilavecz, M.; Wurglics, M. Plasma levels and distribution of flavonoids in rat brain after single and repeated doses of standardized Ginkgo biloba extract EGb 761(R). Planta Med. 2010, 76, 1683–1690. [Google Scholar] [CrossRef]
- Park, H.J.; Lee, S.J.; Cho, J.; Gharbi, A.; Han, H.D.; Kang, T.H.; Kim, Y.; Lee, Y.; Park, W.S.; Jung, I.D.; et al. Tamarixetin Exhibits Anti-inflammatory Activity and Prevents Bacterial Sepsis by Increasing IL-10 Production. J. Nat. Prod. 2018, 81, 1435–1443. [Google Scholar] [CrossRef]
- Son, K.H.; Kwon, S.J.; Chang, H.W.; Kim, H.P.; Kang, S.S. Papyriflavonol A, a new prenylated flavonol from Broussonetiapapyrifera. Fitoterapia 2001, 72, 456–458. [Google Scholar] [CrossRef]
- Ho, L.; Ferruzzi, M.G.; Janle, E.M.; Wang, J.; Gong, B.; Chen, T.-Y.; Lobo, J.; Cooper, B.; Wu, Q.L.; Talcott, S.T.; et al. Identification of brain-targeted bioactive dietary quercetin-3-O-glucuronide as a novel intervention for Alzheimer’s disease. FASEB J. 2013, 27, 769–781. [Google Scholar] [CrossRef]
- Ishisaka, A.; Mukai, R.; Terao, J.; Shibata, N.; Kawai, Y. Specific localization of quercetin-3-O-glucuronide in human brain. Arch. Biochem. Biophys. 2014, 557, 11–17. [Google Scholar] [CrossRef]
- Roubalová, L.; Biedermann, D.; Papoušková, B.; Vacek, J.; Kuzma, M.; Křen, V.; Ulrichová, J.; Dinkova-Kostova, A.T.; Vrba, J. Semisynthetic flavonoid 7-O-galloylquercetin activates Nrf2 and induces Nrf2-dependent gene expression in RAW264.7 and Hepa1c1c7 cells. Chem. Biol. Interact. 2016, 260, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Wallace, C.H.R.; Baczkó, I.; Jones, L.; Fercho, M.; Light, P.E. Inhibition of cardiac voltage-gated sodium channels by grape polyphenols. Br. J. Pharmacol. 2006, 149, 657–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, Y.; Han, D.D.; Zhang, T.; Yang, Z. Quercetin improves cognitive deficits in rats with chronic cerebral ischemia and inhibits voltage-dependent sodium channels in hippocampal CA1 pyramidal neurons. Phytother. Res. 2010, 24, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.W.; Lin, T.Y.; Wang, S.J. Quercetin inhibits depolarization-evoked glutamate release in nerve terminals from rat cerebral cortex. Neurotoxicology 2013, 39, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Wu, H.; Tang, C.; Ke, J.; Wang, Y. Protective effect of quercetin on bupivacaine-induced neurotoxicity via T-type calcium channel inhibition. Trop. J. Pharm. Res. 2017, 16, 1827–1833. [Google Scholar] [CrossRef]
- Haleagrahara, N.; Siew, C.J.; Mitra, N.K.; Kumari, M. Neuroprotective effect of bioflavonoid quercetin in 6-hydroxydopamine-induced oxidative stress biomarkers in the rat striatum. Neurosci. Lett. 2011, 500, 139–143. [Google Scholar] [CrossRef]
- Sriraksa, N.; Wattanathorn, J.; Muchimapura, S.; Tiamkao, S.; Brown, K.; Chaisiwamongkol, K. Cognitive-enhancing effect of quercetin in a rat model of Parkinson’s disease induced by 6-hydroxydopamine. Evid. Based Complement. Altern. Med. 2012, 2012, 823206. [Google Scholar] [CrossRef]
- Lv, C.; Hong, T.; Yang, Z.; Zhang, Y.; Wang, L.; Dong, M.; Zhao, J.; Mu, J.; Meng, Y. Effect of Quercetin in the 1-Methyl-4-phenyl-1, 2, 3, 6-tetrahydropyridine-Induced Mouse Model of Parkinson’s Disease. Evid. Based Complement Altern. Med. 2012, 2012, 928643. [Google Scholar] [CrossRef]
- Naidu, P.S.; Singh, A.; Kulkarni, S.K. D2-dopamine receptor and alpha2-adrenoreceptor-mediated analgesic response of quercetin. Indian J. Exp. Biol. 2003, 41, 1400–1404. [Google Scholar]
- Selvakumar, K.; Bavithra, S.; Krishnamoorthy, G.; Ganesh, A.; Venkataraman, P.; Arunakaran, J. Impact of quercetin on PCBs (Aroclor-1254)-induced impairment of dopaminergic receptors expression in hippocampus of adult male Wistar rats. Biomed. Prev. Nutr. 2013, 3, 42–52. [Google Scholar] [CrossRef]
- Gupta, R.; Shukla, R.K.; Pandey, A.; Sharma, T.; Dhuriya, Y.K.; Srivastava, P.; Singh, M.P.; Siddiqi, M.I.; Pant, A.B.; Khanna, V.K. Involvement of PKA/DARPP-32/PP1α and β- arrestin/Akt/GSK-3β Signaling in Cadmium-Induced DA-D2 Receptor-Mediated Motor Dysfunctions: Protective Role of Quercetin. Sci. Rep. 2018, 8, 2528. [Google Scholar] [CrossRef]
- Tongjaroenbuangam, W.; Ruksee, N.; Chantiratikul, P.; Pakdeenarong, N.; Kongbuntad, W.; Govitrapong, P. Neuroprotective effects of quercetin, rutin and okra (Abelmoschusesculentus Linn.) in dexamethasone-treated mice. Neurochem. Int. 2011, 59, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Holzmann, I.; da Silva, L.M.; Correa da Silva, J.A.; Steimbach, V.M.; de Souza, M.M. Antidepressant-like effect of quercetin in bulbectomized mice and involvement of the antioxidant defenses, and the glutamatergic and oxidonitrergic pathways. Pharmacol. Biochem. Behav. 2015, 136, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, P.; Anuradha, M.; Kar, A. Pharmacophore Screening and Docking studies of AMPA Receptor Implicated in Alzheimerâ   s disease with Some CNS Acting Phytocompounds from Selected Ayurvedic Medicinal Plants. Neuropsychiatry 2018, 8, 1101–1114. [Google Scholar] [CrossRef]
- Moghbelinejad, S.; Mohammadi, G.; Khodabandehloo, F.; Najafipour, R.; NaserpourFarivar, T.; Rashvand, Z.; Nassiri-Asl, M. The Role of Quercetin in Gene Expression of GluR1 Subunit of AMPA Receptors, and NR2A and NR2B Subunits of NMDA Receptors in Kainic Acid Model of Seizure in Mice. Iran. Red Crescent Med. J. 2016, 19, e42415. [Google Scholar] [CrossRef]
- Lee, B.-H.; Choi, S.-H.; Shin, T.-J.; Pyo, M.K.; Hwang, S.-H.; Lee, S.-M.; Paik, H.-D.; Kim, H.-C.; Nah, S.-Y. Effects of quercetin on α9α10 nicotinic acetylcholine receptor-mediated ion currents. Eur. J. Pharmacol. 2011, 650, 79–85. [Google Scholar] [CrossRef]
- Lee, B.-H.; Shin, T.-J.; Hwang, S.-H.; Choi, S.-H.; Kang, J.; Kim, H.-J.; Park, C.-W.; Lee, S.-H.; Nah, S.-Y. Inhibitory Effects of Quercetin on Muscle-type of Nicotinic Acetylcholine Receptor-Mediated Ion Currents Expressed in Xenopus Oocytes. Korean J. Physiol. Pharmacol. Off. J. Korean Physiol. Soc. Korean Soc. Pharmacol. 2011, 15, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.-H.; Hwang, S.-H.; Choi, S.-H.; Shin, T.-J.; Kang, J.; Lee, S.-M.; Nah, S.-Y. Quercetin Inhibits α3β4 Nicotinic Acetylcholine Receptor-Mediated Ion Currents Expressed in Xenopus Oocytes. Korean J. Physiol. Pharmacol. Off. J. Korean Physiol. Soc. Korean Soc. Pharmacol. 2011, 15, 17–22. [Google Scholar] [CrossRef]
- Lee, B.H.; Choi, S.H.; Shin, T.J.; Pyo, M.K.; Hwang, S.H.; Kim, B.R.; Lee, S.M.; Lee, J.H.; Kim, H.C.; Park, H.Y.; et al. Quercetin enhances human alpha7 nicotinic acetylcholine receptor-mediated ion current through interactions with Ca(2+) binding sites. Mol. Cell. 2010, 30, 245–253. [Google Scholar] [CrossRef]
- Lee, B.-H.; Choi, S.-H.; Kim, H.-J.; Jung, S.-W.; Hwang, S.-H.; Pyo, M.-K.; Rhim, H.; Kim, H.-C.; Kim, H.-K.; Lee, S.-M.; et al. Differential Effects of Quercetin and Quercetin Glycosides on Human α7 Nicotinic Acetylcholine Receptor-Mediated Ion Currents. Biomol. Ther. 2016, 24, 410–417. [Google Scholar] [CrossRef]
- Swaminathan, M.; Chee, F.C.; Chin, P.S.; Buckle, J.M.; Rahman, A.N.; Doughty, W.S.; Chung, Y.L. Flavonoids with M1 Muscarinic Acetylcholine Receptor Binding Activity. Molecules 2014, 19, 8933–8948. [Google Scholar] [CrossRef] [Green Version]
- Bandaruk, Y.; Mukai, R.; Terao, J. Cellular uptake of quercetin and luteolin and their effects on monoamine oxidase-A in human neuroblastoma SH-SY5Y cells. Toxicol. Rep. 2014, 1, 639–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Cano, D.; Menendez, C.; Moreno, E.; Moral-Sanz, J.; Barreira, B.; Galindo, P.; Pandolfi, R.; Jimenez, R.; Moreno, L.; Cogolludo, A.; et al. The Flavonoid Quercetin Reverses Pulmonary Hypertension in Rats. PLoS ONE 2014, 9, e114492. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Jeong, S.M.; Lee, J.H.; Kim, J.H.; Yoon, I.S.; Lee, J.H.; Choi, S.H.; Lee, S.M.; Chang, C.G.; Kim, H.C.; et al. Quercetin inhibits the 5-hydroxytryptamine type 3 receptor-mediated ion current by interacting with pre-transmembrane domain I. Mol. Cell. 2005, 20, 69–73. [Google Scholar]
- Jung, J.W.; Lee, S. Anxiolytic effects of quercetin: Involvement of GABAergic system. J. Life Sci. 2014, 24, 290–296. [Google Scholar] [CrossRef]
- Calero, C.I.; González, A.N.B.; Gasulla, J.; Alvarez, S.; Evelson, P.; Calvo, D.J. Quercetin antagonism of GABAAρ1 receptors is prevented by ascorbic acid through a redox-independent mechanism. Eur. J. Pharmacol. 2013, 714, 274–280. [Google Scholar] [CrossRef]
- Moghbelinejad, S.; Alizadeh, S.; Mohammadi, G.; Khodabandehloo, F.; Rashvand, Z.; Najafipour, R.; Nassiri-Asl, M. The effects of quercetin on the gene expression of the GABA A receptor α5 subunit gene in a mouse model of kainic acid-induced seizure. J. Physiol. Sci. 2017, 67, 339–343. [Google Scholar] [CrossRef]
- Fan, H.-R.; Du, W.-F.; Zhu, T.; Wu, Y.-J.; Liu, Y.-M.; Wang, Q.; Wang, Q.; Gu, X.; Shan, X.; Deng, S. Quercetin reduces cortical GABAergic transmission and alleviates MK-801-induced hyperactivity. E Bio. Med. 2018, 34, 201–213. [Google Scholar] [CrossRef]
- Lee, B.-H.; Lee, J.-H.; Yoon, I.-S.; Lee, J.-H.; Choi, S.-H.; Pyo, M.K.; Jeong, S.M.; Choi, W.-S.; Shin, T.-J.; Lee, S.-M. Human glycine α1 receptor inhibition by quercetin is abolished or inversed by α267 mutations in transmembrane domain 2. Brain Res. 2007, 1161, 1–10. [Google Scholar] [CrossRef]
- Sun, H.; Cheng, X.P.; You-Ye, Z.; Jiang, P.; Zhou, J.N. Quercetin subunit specifically reduces GlyR-mediated current in rat hippocampal neurons. Neuroscience 2007, 148, 548–559. [Google Scholar] [CrossRef]
- Kaneko, M.; Takimoto, H.; Sugiyama, T.; Seki, Y.; Kawaguchi, K.; Kumazawa, Y. Suppressive effects of the flavonoids quercetin and luteolin on the accumulation of lipid rafts after signal transduction via receptors. Immunopharmacol. Immunotoxicol. 2008, 30, 867–882. [Google Scholar] [CrossRef]
- Byun, E.B.; Yang, M.S.; Choi, H.G.; Sung, N.Y.; Song, D.S.; Sin, S.J.; Byun, E.H. Quercetin negatively regulates TLR4 signaling induced by lipopolysaccharide through Tollip expression. Biochem. Biophys. Res. Commun. 2013, 431, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Bhaskar, S.; Helen, A. Quercetin modulates toll-like receptor-mediated protein kinase signaling pathways in oxLDL-challenged human PBMCs and regulates TLR-activated atherosclerotic inflammation in hypercholesterolemic rats. Mol. Cell. Biochem. 2016, 423, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.H.; Choi, Y.H.; Moon, S.K.; Kim, W.J.; Kim, G.Y. Quercetin inhibits lipopolysaccharide-induced nitric oxide production in BV2 microglial cells by suppressing the NF-kappaB pathway and activating the Nrf2-dependent HO-1 pathway. Int. Immunopharmacol. 2013, 17, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Y.; Chen, Z.; Jasmer, K.J.; Chuang, D.Y.; Gu, Z.; Hannink, M.; Simonyi, A. Quercetin Attenuates Inflammatory Responses in BV-2 Microglial Cells: Role of MAPKs on the Nrf2 Pathway and Induction of Heme Oxygenase-1. PLoS ONE 2015, 10, 0141509. [Google Scholar] [CrossRef] [PubMed]
- Rahvar, M.; Owji, A.A.; Mashayekhi, F.J. Effect of quercetin on the brain-derived neurotrophic factor gene expression in the rat brain. Bratisl. Lek. Listy 2018, 119, 28–31. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Aboukhatwa, M.A.; Lei, D.L.; Manaye, K.; Khan, I.; Luo, Y. Anti-depressant natural flavonols modulate BDNF and beta amyloid in neurons and hippocampus of double TgAD mice. Neuropharmacology 2010, 58, 911–920. [Google Scholar] [CrossRef] [Green Version]
- Yao, R.-Q.; Qi, D.-S.; Yu, H.-L.; Liu, J.; Yang, L.-H.; Wu, X.-X. Quercetin Attenuates Cell Apoptosis in Focal Cerebral Ischemia Rat Brain Via Activation of BDNF–TrkB–PI3K/Akt Signaling Pathway. Neurochem. Res. 2012, 37, 2777–2786. [Google Scholar] [CrossRef]
- Nakajima, K.; Niisato, N.; Marunaka, Y. Quercetin stimulates NGF-induced neurite outgrowth in PC12 cells via activation of Na(+)/K(+)/2Cl(-) cotransporter. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2011, 28, 147–156. [Google Scholar] [CrossRef]
- Chan, G.K.L.; Hu, W.W.H.; Zheng, Z.X.; Huang, M.; Lin, Y.X.Y.; Wang, C.Y.; Gong, A.G.W.; Yang, X.Y.; Tsim, K.W.K.; Dong, T.T.X. Quercetin Potentiates the NGF-Induced Effects in Cultured PC 12 Cells: Identification by HerboChips Showing a Binding with NGF. Evid.-Based Complement. Altern. Med. 2018, 2018, 1502457. [Google Scholar] [CrossRef]
- Ola, M.S.; Ahmed, M.M.; Shams, S.; Al-Rejaie, S.S. Neuroprotective effects of quercetin in diabetic rat retina. Saudi J. Biol. Sci. 2017, 24, 1186–1194. [Google Scholar] [CrossRef]
- Testa, G.; Gamba, P.; Badilli, U.; Gargiulo, S.; Maina, M.; Guina, T.; Calfapietra, S.; Biasi, F.; Cavalli, R.; Poli, G.; et al. Loading into nanoparticles improves quercetin’s efficacy in preventing neuroinflammation induced by oxysterols. PLoS ONE 2014, 9, e96795. [Google Scholar] [CrossRef] [PubMed]
- Ioannis, B.; Afrodite, D.; Vasilios, P.; Despina, P. Phytochemicals and cognitive health: Are flavonoids doing the trick? Biomed. Pharmacother. 2019, 109, 1488–1497. [Google Scholar] [CrossRef]
- Verghese, P.B.; Castellano, J.M.; Holtzman, D.M. Apolipoprotein E in Alzheimer’s disease and other neurological disorders. Lancet Neurol. 2011, 10, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Boesch-Saadatmandi, C.; Niering, J.; Minihane, A.M.; Wiswedel, I.; Gardeman, A.; Wolffram, S.; Rimbach, G. Impact of apolipoprotein E genotype and dietary quercetin on paraoxonase 1 status in apoE3 and apoE4 transgenic mice. Atherosclerosis 2010, 211, 110–113. [Google Scholar] [CrossRef] [PubMed]
- Egert, S.; Boesch-Saadatmandi, C.; Wolffram, S.; Rimbach, G.; Muller, M.J. Serum lipid and blood pressure responses to quercetin vary in overweight patients by apolipoprotein E genotype. J. Nutr. 2010, 140, 278–284. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hu, J.; Zhong, L.; Wang, N.; Yang, L.; Liu, C.C.; Li, H.; Wang, X.; Zhou, Y.; Zhang, Y.; et al. Quercetin stabilizes apolipoprotein E and reduces brain Abeta levels in amyloid model mice. Neuropharmacology 2016, 108, 179–192. [Google Scholar] [CrossRef]
- Kulkarni, P.; Benzeroual, K. Neuroprotective effect of flavonoids, via up-regulating Nrf2-ARE pathway, in MPP+-induced PC12 cells, as a model of Parkinson’s disease. FASEB J. 2015, 29, 621–623. [Google Scholar]
- Liu, Y.-W.; Liu, X.-L.; Kong, L.; Zhang, M.-Y.; Chen, Y.-J.; Zhu, X.; Hao, Y.-C. Neuroprotection of quercetin on central neurons against chronic high glucose through enhancement of Nrf2/ARE/glyoxalase-1 pathway mediated by phosphorylation regulation. Biomed. Pharmacother. 2019, 109, 2145–2154. [Google Scholar] [CrossRef]
- Bahar, E.; Kim, J.-Y.; Yoon, H. Quercetin Attenuates Manganese-Induced Neuroinflammation by Alleviating Oxidative Stress through Regulation of Apoptosis, iNOS/NF-κB and HO-1/Nrf2 Pathways. Int. J. Mol. Sci. 2017, 18, 1989. [Google Scholar] [CrossRef]
- Wang, D.; Zhao, J.; Li, S.; Shen, G.; Hu, S. Quercetin attenuates domoic acid-induced cognitive deficits in mice. Nutr. Neurosci. 2018, 21, 123–131. [Google Scholar] [CrossRef]
- Dong, F.; Wang, S.; Wang, Y.; Yang, X.; Jiang, J.; Wu, D.; Qu, X.; Fan, H.; Yao, R. Quercetin ameliorates learning and memory via the Nrf2-ARE signaling pathway in d-galactose-induced neurotoxicity in mice. Biochem. Biophys. Res. Commun. 2017, 491, 636–641. [Google Scholar] [CrossRef] [PubMed]
- Dang, W. The controversial world of sirtuins. Drug Discov. Today Technol. 2014, 12, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Heger, V.; Tyni, J.; Hunyadi, A.; Horáková, L.; Lahtela-Kakkonen, M.; Rahnasto-Rilla, M. Quercetin based derivatives as sirtuin inhibitors. Biomed. Pharmacother. 2019, 111, 1326–1333. [Google Scholar] [CrossRef] [PubMed]
- Herskovits, A.Z.; Guarente, L. Sirtuin deacetylases in neurodegenerative diseases of aging. Cell Res. 2013, 23, 746. [Google Scholar] [CrossRef]
- Kim, D.; Nguyen, M.D.; Dobbin, M.M.; Fischer, A.; Sananbenesi, F.; Rodgers, J.T.; Delalle, I.; Baur, J.A.; Sui, G.; Armour, S.M.; et al. SIRT1 deacetylase protects against neurodegeneration in models for Alzheimer’s disease and amyotrophic lateral sclerosis. EMBO J. 2007, 26, 3169–3179. [Google Scholar] [CrossRef]
- Khan, R.I.; Nirzhor, S.S.R.; Akter, R. A Review of the Recent Advances Made with SIRT6 and its Implications on Aging Related Processes, Major Human Diseases, and Possible Therapeutic Targets. Biomolecules 2018, 8, 44. [Google Scholar] [CrossRef]
- Peng, J.; Li, Q.; Li, K.; Zhu, L.; Lin, X.; Lin, X.; Shen, Q.; Li, G.; Xie, X. Quercetin Improves Glucose and Lipid Metabolism of Diabetic Rats: Involvement of Akt Signaling and SIRT1. J. Diabetes Res. 2017, 2017, 3417306. [Google Scholar] [CrossRef]
- Hung, C.H.; Chan, S.H.; Chu, P.M.; Tsai, K.L. Quercetin is a potent anti-atherosclerotic compound by activation of SIRT1 signaling under oxLDL stimulation. Mol. Nutr. Food Res. 2015, 59, 1905–1917. [Google Scholar] [CrossRef]
- Dong, J.; Zhang, X.; Zhang, L.; Bian, H.X.; Xu, N.; Bao, B.; Liu, J. Quercetin reduces obesity-associated ATM infiltration and inflammation in mice: A mechanism including AMPKalpha1/SIRT1. J. Lipid Res. 2014, 55, 363–374. [Google Scholar] [CrossRef]
- Lazo-Gomez, R.; Tapia, R. Quercetin prevents spinal motor neuron degeneration induced by chronic excitotoxic stimulus by a sirtuin 1-dependent mechanism. Transl. Neurodegener. 2017, 6, 31. [Google Scholar] [CrossRef]
- Fusi, J.; Bianchi, S.; Daniele, S.; Pellegrini, S.; Martini, C.; Galetta, F.; Giovannini, L.; Franzoni, F. An in vitro comparative study of the antioxidant activity and SIRT1 modulation of natural compounds. Biomed. Pharmacother. 2018, 101, 805–819. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Sarkar, S.; Mandal, A.K.; Das, N. Neuroprotective role of nanoencapsulated quercetin in combating ischemia-reperfusion induced neuronal damage in young and aged rats. PLoS ONE 2013, 8, e57735. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Ahmad, R.; Naqvi, A.A.; Alam, M.A.; Abdur Rub, R.; Ahmad, F.J. Enhancement of Quercetin Oral Bioavailability by Self-Nanoemulsifying Drug Delivery System and their Quantification Through Ultra High Performance Liquid Chromatography and Mass Spectrometry in Cerebral Ischemia. Drug Res. 2017, 67, 564–575. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl | Common Name | Substituents | Sources | Targets in Neuroprotection | Ability to crossBBB | References | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| R1 | R2 | R3 | R4 | R5 | R6 | R7 | ||||||
| 1 | Quercetin 3′-O-(3-chloropivaloyl) (quercetin pivaloyl ester) | OH | OH | H | OH | H | OH |  | Synthetically modified quercetin | Toll-like receptor (TLR)-4 | + | [43] |
| 2 | 3, 5, 7, 3’, 4’-pentahydroxyflavon (quercetin) | OH | OH | H | OH | OH | OH | H | Elderberries, cranberries, coriander leaves, canned capers | Nuclear factor erythroid 2-related factor 2-antioxidant responsive element (Nrf2-ARE) and antioxidant/anti-inflammatory enzyme paraoxonase 2 (PON2) enzyme | + | [13,44] |
| 3 | Quercetin 3-O-glucoside (isoquercetin) | O-Glc | OH | H | OH | OH | OH | H | Mango fruits, beans, plums, onions | TLR-2 and 4 | + | [44,45,46,47] |
| 4 | Quercetin 3-O-rhamnoside (quercitrin) | O-Rha | OH | H | OH | OH | OH | H | Mango–fruits, pepper–fruits, cranberry, lingonberry | TLR-2 and 4 | − | [28,45,47] |
| 5 | Quercetin 3-O-rhamnosyl-(1–6)-glucoside (rutin) | O-X | OH | H | OH | OH | OH | H | Plums, cherries, tomatoes, buckwheat leaves, buckwheat seeds, chokeberry | − | + | [48,49,50] |
| 6 | Quercetin 7-O-glucoside | OH | OH | H | O-Glc | OH | OH | H | Beans | − | − | [23] |
| 7 | Quercetin 3-O-rhamnoside-7-O-glucoside | O-Y | OH | H | Glc | OH | OH | H | Pepper fruits | − | − | [28] |
| 8 | Quercetin 3-sulfate-7-O-arabinoside | O-Sul | OH | H | O-Ara | OH | OH | H | Salt bush | − | − | [21] |
| 9 | Quercetin 3-O-glucoside-3’-sulfate | O-Glc | OH | H | OH | O-Sul | OH | H | Corn flower | TLR-2 and 4 | − | [47,51] |
| 10 | Quercetin 5-methyl ether (azaleatin) | OH | O-M | H | OH | OH | OH | H | Flowers of Rhododendron mucronatum, Plumbago capensis, Ceratostigmawillmottiana, Carya pecan, leaves of Eucryphia | Extracellular signal-regulated kinase 1/2 (ERK)-pathway | − | [13,52] |
| 11 | Quercetin 7-methyl ether (rhamnetin) | OH | OH | H | O-M | OH | OH | H | Cloves, berries from buckthorn family, such as Rhamnusinfectorius, R. cathartica | − | + | [13,53] |
| 12 | Quercetin 3’-methyl ether (isohramnetin) | OH | OH | H | OH | O-M | OH | H | Honey | TLR-2 and 4 | + | [47,54,55] |
| 13 | Quercetin 4’-methyl ether (tamarixetin) | OH | OH | H | OH | OH | O-M | H | Artemisia annua | TLR-4 and myeloid differentiation primary response 88 | + | [13,55,56] |
| 14 | Quercetin 7-methoxy-3-O-glucoside | O-Glc | OH | H | O-M | OH | OH | H | Honey | − | − | [54] |
| 15 | Quercetin 3’-methoxy-3-O-galactoside | O-Gal | OH | H | OH | O-M | OH | H | Sage | – | − | [24] |
| 16 | 6, 5’-Di-C-prenylquercetin | OH | OH | Z | OH | OH | OH | Z | Paper mullberry | − | − | [57] |
| 17 | Quercetin-3-O-glucuronide |  | OH | H | OH | H | OH | OH | Red wine | Cyclic AMP response element binding protein (CREB) phosphorylation and ↓amyloid beta (Aβ)1-40 | + | [58,59] |
| 18 | 7-O-galloylquercetin | OH | OH | H |  | H | OH | OH | Semisynthetic flavonoid | Activate Nrf2/ARE and ↑antioxidant enzyme NAD(P)H quinone oxidoreducase-1 (NQO1) | − | [60] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakaria, M.; Azam, S.; Jo, S.-H.; Kim, I.-S.; Dash, R.; Choi, D.-K. Potential Therapeutic Targets of Quercetin and Its Derivatives: Its Role in the Therapy of Cognitive Impairment. J. Clin. Med. 2019, 8, 1789. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8111789
Jakaria M, Azam S, Jo S-H, Kim I-S, Dash R, Choi D-K. Potential Therapeutic Targets of Quercetin and Its Derivatives: Its Role in the Therapy of Cognitive Impairment. Journal of Clinical Medicine. 2019; 8(11):1789. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8111789
Chicago/Turabian StyleJakaria, Md., Shofiul Azam, Song-Hee Jo, In-Su Kim, Raju Dash, and Dong-Kug Choi. 2019. "Potential Therapeutic Targets of Quercetin and Its Derivatives: Its Role in the Therapy of Cognitive Impairment" Journal of Clinical Medicine 8, no. 11: 1789. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8111789