The Rise of Mitochondria in Peripheral Arterial Disease Physiopathology: Experimental and Clinical Data

 , ,
, ,
Abstract
:1. Introduction
2. Mitochondrial Function under Normal and Pathological Conditions
2.1. Normal Condition
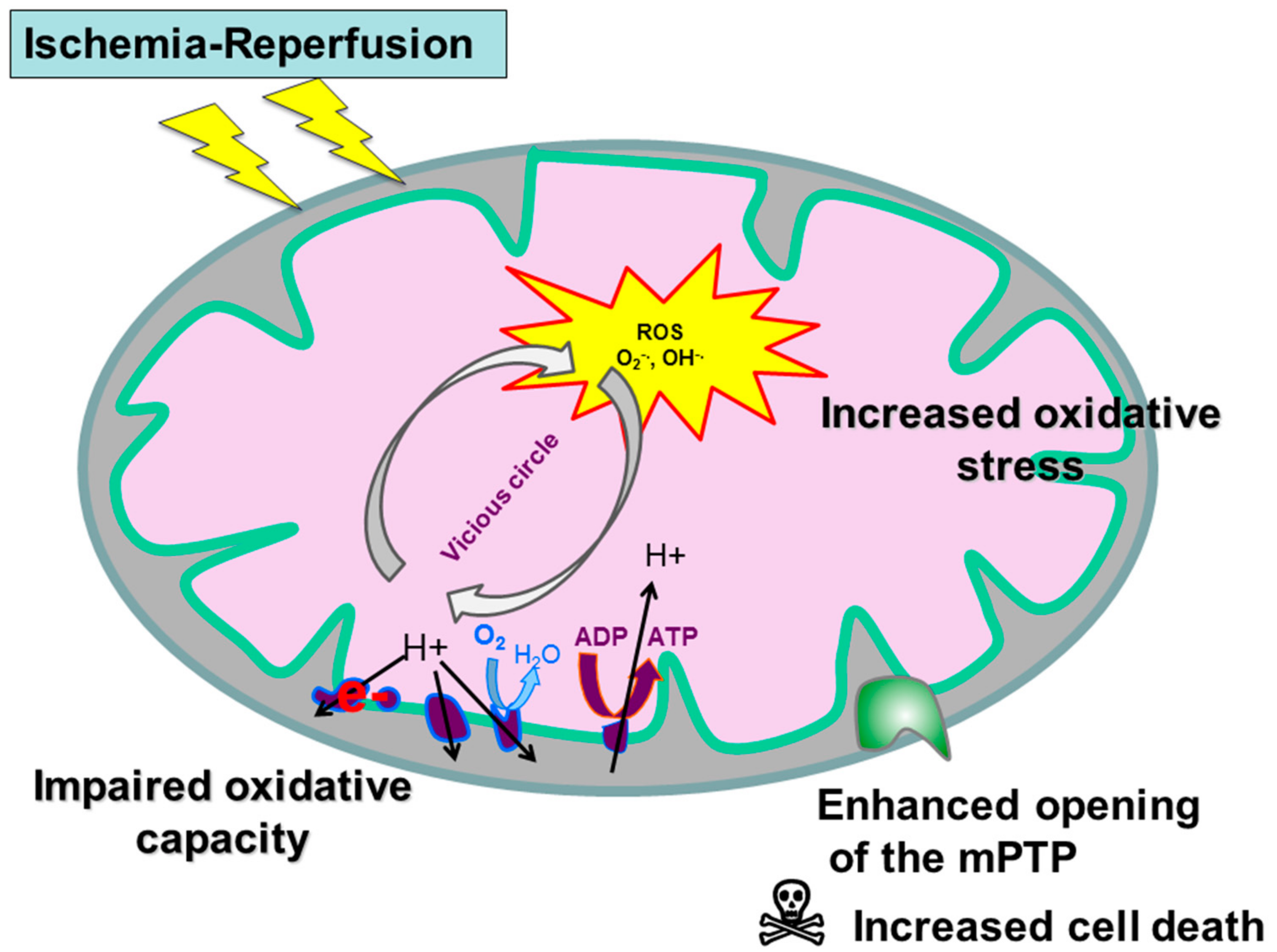
2.2. Ischemic Condition
3. Mitochondrial Oxidative Capacities in PAD
3.1. Experimental Data
3.2. Clinical Data
4. Reactive Oxygen Species Production, Proteins, Lipids and DNA Alterations and Impaired Antioxidant Defense, in PAD
4.1. Experimental Data
4.2. Clinical Data
5. Mitochondrial Implication in Apoptosis during PAD
5.1. Experimental Data
5.2. Clinical Data
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Criqui, M.H.; Aboyans, V. Epidemiology of peripheral artery disease. Circ. Res. 2015, 116, 1509–1526. [Google Scholar] [CrossRef]
- Duff, S.; Mafilios, M.S.; Bhounsule, P.; Hasegawa, J.T. The burden of critical limb ischemia: A review of recent literature. Vasc. Health. Risk. Manag. 2019, 15, 187–208. [Google Scholar] [CrossRef] [PubMed]
- Dua, A.; Lee, C.J. Epidemiology of Peripheral Arterial Disease and Critical Limb Ischemia. Tech. Vasc. Interv. Radiol. 2016, 19, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Leardini-Tristao, M.; Charles, A.L.; Lejay, A.; Pizzimenti, M.; Meyer, A.; Estato, V.; Tibiriçá, E.; Andres, E.; Geny, B. Beneficial Effect of Exercise on Cognitive Function during Peripheral Arterial Disease: Potential Involvement of Myokines and Microglial Anti-Inflammatory Phenotype Enhancement. J. Clin. Med. 2019, 8, 653. [Google Scholar] [CrossRef] [PubMed]
- Aboyans, V.; Ricco, J.B.; Bartelink, M.L.E.L.; Björck, M.; Brodmann, M.; Cohnert, T.; Collet, J.P.; Czerny, M.; De Carlo, M.; Debus, S.; et al. Editor’s Choice—2017 ESC Guidelines on the Diagnosis and Treatment of Peripheral Arterial Diseases, in collaboration with the European Society for Vascular Surgery (ESVS). Eur. J. Vasc. Endovasc. Surg. 2018, 55, 305–368. [Google Scholar] [CrossRef]
- Conte, M.S.; Bradbury, A.W.; Kolh, P.; White, J.V.; Dick, F.; Fitridge, R.; Mills, J.L.; Ricco, J.B.; Suresh, K.R.; Murad, M.H.; et al. Global Vascular Guidelines on the Management of Chronic Limb-Threatening Ischemia. Eur. J. Vasc. Endovasc. Surg. 2019, 58, S1–S109.e33. [Google Scholar] [CrossRef]
- Paradis, S.; Charles, A.L.; Meyer, A.; Lejay, A.; Scholey, J.W.; Chakfé, N.; Zoll, J.; Geny, B. Chronology of mitochondrial and cellular events during skeletal muscle ischemia-reperfusion. Am. J. Physiol.-Cell Physiol. 2016, 310, C968–C982. [Google Scholar] [CrossRef]
- Koutakis, P.; Ismaeel, A.; Farmer, P.; Purcell, S.; Smith, R.S.; Eidson, J.L.; Bohannon, W.T. Oxidative stress and antioxidant treatment in patients with peripheral artery disease. Physiol. Rep. 2018, 6, e13650. [Google Scholar] [CrossRef]
- Walker, J.E. ATP Synthesis by Rotary Catalysis (Nobel lecture). Angew. Chem. Int. Edit. 1998, 37, 2308–2319. [Google Scholar] [CrossRef]
- Walker, J.E.; Dickson, V.K. The peripheral stalk of the mitochondrial ATP synthase. Biochim. Biophys. Acta (BBA)—Bioenerg. 2006, 1757, 286–296. [Google Scholar] [CrossRef]
- Walker, J.E. The ATP synthase: The understood, the uncertain and the unknown. Biochem. Soc. Trans. 2013, 41, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lejay, A.; Meyer, A.; Schlagowski, A.I.; Charles, A.L.; Singh, F.; Bouitbir, J.; Pottecher, J.; Chakfé, N.; Zoll, J.; Geny, B. Mitochondria: Mitochondrial participation in ischemia-reperfusion injury in skeletal muscle. Int. J. Biochem. Cell Biol. 2014, 50, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Makris, K.I.; Nella, A.A.; Zhu, Z.; Swanson, S.A.; Casale, G.P.; Gutti, T.L.; Judge, A.R.; Pipinos, I.I. Mitochondriopathy of peripheral arterial disease. Vascular 2007, 15, 336–343. [Google Scholar] [CrossRef]
- Bernardi, P. The mitochondrial permeability transition pore: A mystery solved? Front. Physiol. 2013, 4, 95. [Google Scholar] [CrossRef] [PubMed]
- Chouchani, E.T.; Pell, V.R.; James, A.M.; Work, L.M.; Saeb-Parsy, K.; Frezza, C.; Krieg, T.; Murphy, M.P. A Unifying Mechanism for Mitochondrial Superoxide Production during Ischemia-Reperfusion Injury. Cell Metab. 2016, 23, 254–263. [Google Scholar] [CrossRef]
- Chouchani, E.T.; Pell, V.R.; Gaude, E.; Aksentijević, D.; Sundier, S.Y.; Robb, E.L.; Logan, A.; Nadtochiy, S.M.; Ord, E.N.J.; Smith, A.C.; et al. Ischaemic accumulation of succinate controls reperfusion injury through mitochondrial ROS. Nature 2014, 515, 431–435. [Google Scholar] [CrossRef]
- Chouchani, E.T.; Methner, C.; Nadtochiy, S.M.; Logan, A.; Pell, V.R.; Ding, S.; James, A.M.; Cochemé, H.M.; Reinhold, J.; Lilley, K.S.; et al. Cardioprotection by S-nitrosation of a cysteine switch on mitochondrial complex I. Nat. Med. 2013, 19, 753–759. [Google Scholar] [CrossRef]
- Pottecher, J.; Kindo, M.; Chamaraux-Tran, T.N.; Charles, A.L.; Lejay, A.; Kemmel, V.; Vogel, T.; Chakfe, N.; Zoll, J.; Diemunsch, P.; et al. Skeletal muscle ischemia-reperfusion injury and cyclosporine A in the aging rat. Fundam. Clin. Pharmacol. 2016, 30, 216–225. [Google Scholar] [CrossRef]
- Thaveau, F.; Zoll, J.; Bouitbir, J.; N’guessan, B.; Plobner, P.; Chakfe, N.; Kretz, J.G.; Richard, R.; Piquard, F.; Geny, B. Effect of chronic pre-treatment with angiotensin converting enzyme inhibition on skeletal muscle mitochondrial recovery after ischemia/reperfusion. Fundam. Clin. Pharmacol. 2010, 24, 333–340. [Google Scholar] [CrossRef]
- Brandão, M.L.; Roselino, J.E.S.; Piccinato, C.E.; Cherri, J. Mitochondrial alterations in skeletal muscle submitted to total ischemia. J. Surg. Res. 2003, 110, 235–240. [Google Scholar] [CrossRef]
- Mansour, Z.; Bouitbir, J.; Charles, A.L.; Talha, S.; Kindo, M.; Pottecher, J.; Zoll, J.; Geny, B. Remote and local ischemic preconditioning equivalently protects rat skeletal muscle mitochondrial function during experimental aortic cross-clamping. J. Vasc. Surg. 2012, 55, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Pipinos, I.I.; Swanson, S.A.; Zhu, Z.; Nella, A.A.; Weiss, D.J.; Gutti, T.L.; McComb, R.D.; Baxter, B.T.; Lynch, T.G.; Casale, G.P. Chronically ischemic mouse skeletal muscle exhibits myopathy in association with mitochondrial dysfunction and oxidative damage. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R290–R296. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.A.; Ryan, T.E.; Lin, C.T.; Inigo, M.M.R.; Green, T.D.; Brault, J.J.; Spangenburg, E.E.; McClung, J.M. Diminished force production and mitochondrial respiratory deficits are strain-dependent myopathies of subacute limb ischemia. J. Vasc. Surg. 2017, 65, 1504–1514.e11. [Google Scholar] [CrossRef] [PubMed]
- Charles, A.L.; Guilbert, A.S.; Guillot, M.; Talha, S.; Lejay, A.; Meyer, A.; Kindo, M.; Wolff, V.; Bouitbir, J.; Zoll, J.; et al. Muscles Susceptibility to Ischemia-Reperfusion Injuries Depends on Fiber Type Specific Antioxidant Level. Front. Physiol. 2017, 8, 52. [Google Scholar] [CrossRef]
- Paradis, S.; Charles, A.L.; Georg, I.; Goupilleau, F.; Meyer, A.; Kindo, M.; Laverny, G.; Metzger, D.; Geny, B. Aging Exacerbates Ischemia-Reperfusion-Induced Mitochondrial Respiration Impairment in Skeletal Muscle. Antioxidants (Basel) 2019, 8, 168. [Google Scholar] [CrossRef]
- Pottecher, J.; Adamopoulos, C.; Lejay, A.; Bouitbir, J.; Charles, A.L.; Meyer, A.; Singer, M.; Wolff, V.; Diemunsch, P.; Laverny, G.; et al. Diabetes Worsens Skeletal Muscle Mitochondrial Function, Oxidative Stress, and Apoptosis After Lower-Limb Ischemia-Reperfusion: Implication of the RISK and SAFE Pathways? Front. Physiol. 2018, 9, 579. [Google Scholar] [CrossRef]
- Koutakis, P.; Miserlis, D.; Myers, S.A.; Kim, J.K.S.; Zhu, Z.; Papoutsi, E.; Swanson, S.A.; Haynatzki, G.; Ha, D.M.; Carpenter, L.A.; et al. Abnormal accumulation of desmin in gastrocnemius myofibers of patients with peripheral artery disease: Associations with altered myofiber morphology and density, mitochondrial dysfunction and impaired limb function. J. Histochem. Cytochem. 2015, 63, 256–269. [Google Scholar] [CrossRef]
- Pipinos, I.I.; Judge, A.R.; Zhu, Z.; Selsby, J.T.; Swanson, S.A.; Johanning, J.M.; Baxter, B.T.; Lynch, T.G.; Dodd, S.L. Mitochondrial defects and oxidative damage in patients with peripheral arterial disease. Free Radic. Biol. Med. 2006, 41, 262–269. [Google Scholar] [CrossRef]
- Pipinos, I.I.; Sharov, V.G.; Shepard, A.D.; Anagnostopoulos, P.V.; Katsamouris, A.; Todor, A.; Filis, K.A.; Sabbah, H.N. Abnormal mitochondrial respiration in skeletal muscle in patients with peripheral arterial disease. J. Vasc. Surg. 2003, 38, 827–832. [Google Scholar] [CrossRef]
- Brass, E.P.; Hiatt, W.R.; Gardner, A.W.; Hoppel, C.L. Decreased NADH dehydrogenase and ubiquinol-cytochrome c oxidoreductase in peripheral arterial disease. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H603–H609. [Google Scholar] [CrossRef]
- Hart, C.R.; Layec, G.; Trinity, J.D.; Le Fur, Y.; Gifford, J.R.; Clifton, H.L.; Richardson, R.S. Oxygen availability and skeletal muscle oxidative capacity in patients with peripheral artery disease: Implications from in vivo and in vitro assessments. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H897–H909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hart, C.R.; Layec, G.; Trinity, J.D.; Kwon, O.S.; Zhao, J.; Reese, V.R.; Gifford, J.R.; Richardson, R.S. Increased skeletal muscle mitochondrial free radical production in peripheral arterial disease despite preserved mitochondrial respiratory capacity. Exp. Physiol. 2018, 103, 838–850. [Google Scholar] [CrossRef]
- Morisaki, K.; Furuyama, T.; Matsubara, Y.; Inoue, K.; Kurose, S.; Yoshino, S.; Nakayama, K.; Yamashita, S.; Yoshiya, K.; Yoshiga, R.; et al. External validation of CLI Frailty Index and assessment of predictive value of modified CLI Frailty Index for patients with critical limb ischemia undergoing infrainguinal revascularization. Vascular 2019, 1708538119836005. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, R.; Deguchi, J.; Hashimoto, T.; Sato, O. Sarcopenia as a Possible Negative Predictor of Limb Salvage in Patients with Chronic Limb-Threatening Ischemia. Ann. Vasc. Dis. 2019, 12, 194–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsubara, Y.; Matsumoto, T.; Aoyagi, Y.; Tanaka, S.; Okadome, J.; Morisaki, K.; Shirabe, K.; Maehara, Y. Sarcopenia is a prognostic factor for overall survival in patients with critical limb ischemia. J. Vasc. Surg. 2015, 61, 945–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AlGhatrif, M.; Zane, A.; Oberdier, M.; Canepa, M.; Studenski, S.; Simonsick, E.; Spencer, R.G.; Fishbein, K.; Reiter, D.; Lakatta, E.G.; et al. Lower Mitochondrial Energy Production of the Thigh Muscles in Patients with Low-Normal Ankle-Brachial Index. J. Am. Heart Assoc 2017, 6. [Google Scholar] [CrossRef] [PubMed]
- Pipinos, I.I.; Shepard, A.D.; Anagnostopoulos, P.V.; Katsamouris, A.; Boska, M.D. Phosphorus 31 nuclear magnetic resonance spectroscopy suggests a mitochondrial defect in claudicating skeletal muscle. J. Vasc. Surg. 2000, 31, 944–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.Y.; Green, S.; Askew, C.D.; Barker, G.; Green, A.; Walker, P.J. Skeletal muscle mitochondrial ATP production rate and walking performance in peripheral arterial disease. Clin. Physiol. Funct. Imaging 2002, 22, 226–232. [Google Scholar] [CrossRef]
- Lindegaard, B.P.; Bækgaard, N.; Quistorff, B. Mitochondrial dysfunction in calf muscles of patients with combined peripheral arterial disease and diabetes type 2. Int. Angiol. 2017, 36, 482–495. [Google Scholar]
- Tecilazich, F.; Dinh, T.; Lyons, T.E.; Guest, J.; Villafuerte, R.A.; Sampanis, C.; Gnardellis, C.; Zuo, C.S.; Veves, A. Postexercise phosphocreatine recovery, an index of mitochondrial oxidative phosphorylation, is reduced in diabetic patients with lower extremity complications. J. Vasc. Surg. 2013, 57, 997–1005. [Google Scholar] [CrossRef] [Green Version]
- Zorov, D.B.; Juhaszova, M.; Sollott, S.J. Mitochondrial ROS-induced ROS release: An update and review. Biochim. Biophys. Acta. 2006, 1757, 509–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lejay, A.; Choquet, P.; Thaveau, F.; Singh, F.; Schlagowski, A.; Charles, A.L.; Laverny, G.; Metzger, D.; Zoll, J.; Chakfe, N.; et al. A new murine model of sustainable and durable chronic critical limb ischemia fairly mimicking human pathology. Eur. J. Vasc. Endovasc. Surg. 2015, 49, 205–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, B.; Kang, C.; Kim, J.; Yoo, D.; Cho, B.R.; Kang, P.M.; Lee, D. H2O2-responsive antioxidant polymeric nanoparticles as therapeutic agents for peripheral arterial disease. Int. J. Pharm. 2016, 511, 1022–1032. [Google Scholar] [CrossRef] [PubMed]
- Lejay, A.; Charles, A.L.; Georg, I.; Goupilleau, F.; Delay, C.; Talha, S.; Thaveau, F.; Chakfe, N.; Geny, B. Critical limb ischemia exacerbates mitochondrial dysfunction in ApoE-/- mice compared to ApoE+/+ mice, but N-acetyl cysteine still confers protection. Eur. J. Vasc. Endovasc. Surg. 2019, 58, 576–582. [Google Scholar] [CrossRef]
- Tran, T.P.; Tu, H.; Pipinos, I.I.; Muelleman, R.L.; Albadawi, H.; Li, Y.L. Tourniquet-induced acute ischemia-reperfusion injury in mouse skeletal muscles: Involvement of superoxide. Eur. J. Pharmacol. 2011, 650, 328–334. [Google Scholar] [CrossRef] [Green Version]
- Miura, S.; Saitoh, S.; Kokubun, T.; Owada, T.; Yamauchi, H.; Machii, H.; Takeishi, Y. Mitochondrial-Targeted Antioxidant Maintains Blood Flow, Mitochondrial Function, and Redox Balance in Old Mice Following Prolonged Limb Ischemia. Int. J. Mol. Sci. 2017, 18, 1897. [Google Scholar] [CrossRef] [Green Version]
- Lejay, A.; Laverny, G.; Paradis, S.; Schlagowski, A.I.; Charles, A.L.; Singh, F.; Zoll, J.; Thaveau, F.; Lonsdorfer, E.; Dufour, S.; et al. Moderate Exercise Allows for shorter Recovery Time in Critical Limb Ischemia. Front. Physiol. 2017, 8, 523. [Google Scholar] [CrossRef] [Green Version]
- Guillot, M.; Charles, A.L.; Chamaraux-Tran, T.N.; Bouitbir, J.; Meyer, A.; Zoll, J.; Schneider, F.; Geny, B. Oxidative stress precedes skeletal muscle mitochondrial dysfunction during experimental aortic cross-clamping but is not associated with early lung, heart, brain, liver, or kidney mitochondrial impairment. J. Vasc. Surg. 2014, 60, 1043–1051.e5. [Google Scholar] [CrossRef] [Green Version]
- McDermott, M.M.; Peterson, C.A.; Sufit, R.; Ferrucci, L.; Guralnik, J.M.; Kibbe, M.R.; Polonsky, T.S.; Tian, L.; Criqui, M.H.; Zhao, L.; et al. Peripheral artery disease, calf skeletal muscle mitochondrial DNA copy number, and functional performance. Vasc. Med. 2018, 23, 340–348. [Google Scholar] [CrossRef] [Green Version]
- Koutakis, P.; Weiss, D.J.; Miserlis, D.; Shostrom, V.K.; Papoutsi, E.; Ha, D.M.; Carpenter, L.A.; McComb, R.D.; Casale, G.P.; Pipinos, I.I. Oxidative damage in the gastrocnemius of patients with peripheral artery disease is myofiber type selective. Redox. Biol. 2014, 2, 921–928. [Google Scholar] [CrossRef] [Green Version]
- Weiss, D.J.; Casale, G.P.; Koutakis, P.; Nella, A.A.; Swanson, S.A.; Zhu, Z.; Miserlis, D.; Johanning, J.M.; Pipinos, I.I. Oxidative damage and myofiber degeneration in the gastrocnemius of patients with peripheral arterial disease. J. Transl. Med. 2013, 11, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brass, E.P.; Wang, H.; Hiatt, W.R. Multiple skeletal muscle mitochondrial DNA deletions in patients with unilateral peripheral arterial disease. Vasc. Med. 2000, 5, 225–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, H.K.; Hiatt, W.R.; Hoppel, C.L.; Brass, E.P. Skeletal muscle mitochondrial DNA injury in patients with unilateral peripheral arterial disease. Circulation 1999, 99, 807–812. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.L.; Yin, T.C.; Shao, P.L.; Chen, K.H.; Wu, R.W.; Chen, C.C.; Lin, P.Y.; Chung, S.Y.; Sheu, J.J.; Sung, P.H.; et al. Hyperbaric oxygen facilitates the effect of endothelial progenitor cell therapy on improving outcome of rat critical limb ischemia. Am. J. Transl. Res. 2019, 11, 1948–1964. [Google Scholar]
- Sheu, J.J.; Lee, F.Y.; Wallace, C.G.; Tsai, T.H.; Leu, S.; Chen, Y.L.; Chai, H.T.; Lu, H.I.; Sun, C.K.; Yip, H.K. Administered circulating microparticles derived from lung cancer patients markedly improved angiogenesis, blood flow and ischemic recovery in rat critical limb ischemia. J. Transl. Med. 2015, 13. [Google Scholar] [CrossRef] [Green Version]
- Tetsi, L.; Charles, A.L.; Georg, I.; Goupilleau, F.; Lejay, A.; Talha, S.; Maumy-Bertrand, M.; Lugnier, C.; Geny, B. Effect of the Phosphodiesterase 5 Inhibitor Sildenafil on Ischemia-Reperfusion-Induced Muscle Mitochondrial Dysfunction and Oxidative Stress. Antioxidants (Basel) 2019, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Lejay, A.; Paradis, S.; Lambert, A.; Charles, A.L.; Talha, S.; Enache, I.; Thaveau, F.; Chakfe, N.; Geny, B. N-Acetyl Cysteine Restores Limb Function, Improves Mitochondrial Respiration, and Reduces Oxidative Stress in a Murine Model of Critical Limb Ischaemia. Eur. J. Vasc. Endovasc. Surg. 2018, 56, 730–738. [Google Scholar] [CrossRef] [Green Version]
- Masud, R.; Shameer, K.; Dhar, A.; Ding, K.; Kullo, I.J. Gene expression profiling of peripheral blood mononuclear cells in the setting of peripheral arterial disease. J. Clin. Bioinforma. 2012, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, R.G.; Duscha, B.D.; Robbins, J.L.; Redfern, S.I.; Chung, J.; Bensimhon, D.R.; Kraus, W.E.; Hiatt, W.R.; Regensteiner, J.G.; Annex, B.H. Increased levels of apoptosis in gastrocnemius skeletal muscle in patients with peripheral arterial disease. Vasc. Med. 2007, 12, 285–290. [Google Scholar] [CrossRef]
- Gardner, A.W.; Parker, D.E.; Montgomery, P.S.; Sosnowska, D.; Casanegra, A.I.; Ungvari, Z.; Csiszar, A.; Sonntag, W.E. Greater Endothelial Apoptosis and Oxidative Stress in Patients with Peripheral Artery Disease. Int. J. Vasc. Med. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Skórkowska-Telichowska, K.; Adamiec, R.; Tuchendler, D.; Gasiorowski, K. Susceptibility to apoptosis of lymphocytes from patients with peripheral arterial disease. Clin. Investig. Med. 2009, 32, E345–E351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, A.W.; Parker, D.E.; Montgomery, P.S.; Sosnowska, D.; Casanegra, A.I.; Esponda, O.L.; Ungvari, Z.; Csiszar, A.; Sonntag, W.E. Impaired vascular endothelial growth factor A and inflammation in patients with peripheral artery disease. Angiology 2014, 65, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Alves-Cabratosa, L.; Garcia-Gil, M.; Comas-Cufí, M.; Blanch, J.; Ponjoan, A.; Martí-Lluch, R.; Elosua-Bayes, M.; Parramon, D.; Camós, L.; Ramos, R. Role of Low Ankle-Brachial Index in Cardiovascular and Mortality Risk Compared with Major Risk Conditions. J. Clin. Med. 2019, 8, 870. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Animals | Study Design Ischemia-Reperfusion Duration | Outcomes Measured | Main Results | Reference |
|---|---|---|---|---|
| Mice, n = 25 young (23 ± 1 weeks) and old (84 ± 1 weeks) | Unilateral tourniquet I: 2 h/R: 2 h | Skeletal muscle mitochondrial capacity (by oxygraphy) | Impaired mitochondrial respiration in young PAD mice compared with sham (VADP 33.0 ± 2.4 for the contralateral limb versus 18.4 ± 3.8 for the ischemic limb, p < 0.01). Enhanced impairment in old PAD mice (VADP 5.9 ± 2.7 pmol/s/mg wet weight, p < 0.001). | Paradis et al., 2019, Antioxidants [25]. |
| Rats, n = 36 diabetic and non-diabetic | Aortic banding I: 3 h/R: 2 h | Skeletal muscle mitochondrial capacity (by oxygraphy) | Significant decline in mitochondrial respiration after ischemia-reperfusion injury in diabetic rats compared to non-diabetic (p < 0.05). | Pottecher et al., 2018, Front Physiol [26]. |
| Mice, n = 69 BALB/c (ischemia susceptible) and C57BL/6 (ischemia protected) | Aortic banding I and R: duration not specified | Skeletal muscle mitochondrial capacity (by western blot and oxygraphy) | Skeletal muscle mitochondrial impairments in BALB/c limb muscle but not in C57BL/6 (p < 0.01). | Schmidt et al., 2017, J Vasc Surg [23]. |
| Mice, n = 22 | Aortic banding I: 3 h/R: 2 h | Skeletal muscle mitochondrial capacity (by oxygraphy) | Decreased mitochondrial respiration in glycolytic versus oxidative muscles. | Charles et al., 2017, Front Physiol [24]. |
| Rats, n = 12 Old 71-73 weeks | Unilateral tourniquet I: 3 h/R: 2 h | Skeletal muscle mitochondrial capacity (by oxygraphy) | Reduced mitochondrial complexes I, II and IV activities in PAD muscles compared with contralateral ones (VMAX 7.34 ± 1.5 versus 2.87 ± 1.22 µmol O2/min/g dry weight for PAD muscles, p < 0.05). | Pottecher et al., 2016, Fundam Clin Pharmacol [18]. |
| Rats n = 28 | Aortic banding I: 3 h/R: 2 h | Skeletal muscle mitochondrial capacity (by oxygraphy) | IR reduced V(max) (−21.2%, 6.6 ± 1 versus 5.2 ± 1 μmol O2/min/g dry weight, p = 0.001), V(succ) (−22.2%, p = 0.032), and V(TMPD) (−22.4%, p = 0.033). | Mansour et al., 2012, J Vasc Surg [21]. |
| Rats, n = 22 | Unilateral tourniquet I: 5 h R: 5 min | Skeletal muscle mitochondrial capacity (by oxygraphy) | Reduced mitochondrial complexes I, II and IV activities in PAD rats compared with sham (VMAX 4.4 ± 0.4 versus 8.7 ± 0.5 µmol O2/min/g dry weight, p < 0.001). | Thaveau et al., 2010, Fundam Clin Pharmacol [19]. |
| Mice, n = 48 | CLTI Sequential left femoral and iliac ligations R: week 12 | Skeletal muscle mitochondrial capacity (by oxygraphy) | Reduced activity of complexes I (by 34%), III (by 45%) and IV (by 42%) in ischemic muscles compared with controls (p < 0.05). | Pipinos et al., 2008, Am J Physiol Regul Integr Comp Physiol [22]. |
| Rats, n = 20 | Unilateral tourniquet I: 5 h | Respiration of isolated mitochondria (by polarographic analysis) | Inhibition of the mitochondrial respiratory chain | Brandão et al., J Surg Res. 2003 [20]. |
| Population | Number Studied (Symptomatic/Controls) | Outcomes Measured | Main Results | Reference |
|---|---|---|---|---|
| Early stage PAD | 10/11 | O2 delivery, tissue oxygenation and Vmax (by high-resolution respirometry). Skeletal muscle mitochondrial capacity (by oxygraphy) | PAD patients exhibited significantly lower O2 delivery (p < 0.05), tissue-reoxygenation (58 ± 3 % for controls versus 44 ± 3 % for PAD patients, p < 0.05) and Vmax (p < 0.05) during exercise, compared with healthy controls. No differences were found in the mitochondrial respiration rate. | Hart et al., 2018, Am J Physiol Heart Circ Physiol [31]. |
| Claudicant PAD | 10/12 | Skeletal muscle mitochondrial capacity (by oxygraphy) | No differences were found in the mitochondrial respiration rate between PAD patients and healthy controls. | Hart et al., 2018, Exp Physiol [32]. |
| Claudicant PAD; claudicant PAD + DT2 | 15 (PAD)/15 (PAD + DT2)/10 (controls) | Skeletal muscle mitochondrial capacity (by oxygraphy) | Significant reduction in oxygen consumption in the PAD+DT2 group compared with the PAD group or the control group (p < 0.05). No differences were found in the mitochondrial respiration rate between PAD patients and healthy controls. | Lindegaard et al., 2017, Int Angiol [39]. |
| Patients with low ABI | 82 (ABI of 0.90 to 1.10)/281 (ABI of 1.11 to 1.40) | Phosphocreatine recovery (by phosphorus-31 magnetic resonance spectroscopy) | Significantly lower muscle mitochondrial energy production in patients with lower ABI, compared with those with higher ABI (20.8 ms−1 for higher ABI versus 19.3 ms−1 for lower ABI, p = 0.015). | AlGhatrif et al., 2017, J Am Heart Assoc [36]. |
| PAD (no stage specified) | 30/30 | Skeletal muscle mitochondrial capacity (by oxygraphy) | PAD subjects presented significantly lower respiratory activity compared with controls (p < 0.05). | Koutakis et al., 2015, J Histochem Cytochem [27]. |
| Claudicant PAD + neuropathy + DT2 | 7/14 | Phosphocreatine recovery (by phosphorus-31 magnetic resonance spectroscopy) | Reduced mitochondrial oxidative phosphorylation in DT2 patients with lower extremity complications (neuropathy and PAD) (p < 0.05). | Tecilazich et al., 2013, J Vasc Surg [40]. |
| Claudicant PAD; CLI | 25/16 | Skeletal muscle mitochondrial capacity (by spectrophotometry) | Decreased activity of complexes I, III and IV in PAD muscle compared to control (p < 0.05). | Pipinos et al., 2006, Free Radic Biol Med [28]. |
| Claudicant PAD; CLI | 9/9 | Skeletal muscle mitochondrial capacity (by oxygraphy) | Significantly lower respiratory rates, and lower acceptor control ratio (2.90 ± 0.20 for controls versus 1.41 ± 0.10 for PAD) in patients with PAD compared with controls (p < 0.05). | Pipinos et al., 2003, J Vasc Surg [29]. |
| Claudicant PAD | 7/11 | ATP synthesis (by luminometer) | Similar mitochondrial ATP production rate were in PAD patients and healthy controls. | Hou et al., 2002, Clin Physiol Funct Imaging [38]. |
| Claudicant PAD | 17/9 | Skeletal muscle mitochondrial capacity (by spectrophotometry) | Significant reduction in NADH dehydrogenase and ubiquinol-cytochrome c oxidoreductase activity by 27% and 38%, respectively, in PAD compared with controls (p < 0.05). | Brass et al., 2001, Am J Physiol Heart Circ Physiol [30]. |
| Claudicant PAD | 12/14 | Phosphocreatine and ADP recovery (by phosphorus-31 magnetic resonance spectroscopy) | Defective phosphocreatine (44 ± 3 s for controls versus 137 ± 41 s for PAD) and ADP recovery (29 ± 2 s versus 60 ± 10 s for PAD) in PAD compared with controls (p < 0.05). | Pipinos et al., 2000, J Vasc Surg [37]. |
| Animals | Study Design Ischemia-Reperfusion Duration | Outcomes Measured | Main Results | Reference |
|---|---|---|---|---|
| Mice, n = 7 ApoE-/- versus ApoE+/+ | CLTI Sequential right femoral and iliac ligations. R: day 40 | Free radical measurement (by electron paramagnetic resonance spectroscopy) | Enhanced ROS production in muscles of ApoE-/- (+63.6%) and ApoE+/+ (+41.4%) mice compared with contralateral muscles. | Lejay et al., 2019, Eur J Vasc Endovasc Surg [44]. |
| Rats, n = 36 diabetic versus non-diabetic | Aortic banding I: 3 h/R: 2 h | DHE measures of ROS (by epifluorescence microscopy) | Increase in normalized DHE fluorescence in diabetic PAD compared to diabetic controls (p < 0.001). | Pottecher et al., 2018, Front Physiol [26]. |
| Mice, n = 20 | CLTI Left femoral ligation. R: day 21 | mtDNA damage quantification (by quantitative PCR) | Increase in mtDNA damages in ischemic muscles of PAD mice compared with sham (p < 0.05). | Miura et al., 2017, Int J Mol Sci [46]. |
| Mice, n = 20 | CLTI Sequential right femoral and iliac ligations. R: day 30 | Antioxidant quantification (by quantitative PCR) | Significant decrease in mRNA expression of the antioxidant enzymes SOD1 (0.39 ± 0.10 for sham limb versus 0.10 ± 0.06 for ischemic limb), SOD2 (0.32 ± 0.16 versus 0.11 ± 0.07) and catalase (0.38 ± 0.04 versus 0.22 ± 0.11) in ischemic muscles compared with control ones (p < 0.05). | Lejay et al., 2017, Front Physiol [47]. |
| Mice, n = 22 | Aortic banding I: 3 h/R: 2 h | Free radical measurement (by electron paramagnetic resonance spectroscopy) | Ischemia-reperfusion injury increased ROS production in ischemic muscles compared to no ischemic contralateral (+79.15 ± 28.72%, p = 0.04). | Charles et al., 2017, Front Physiol [24]. |
| Mice, n = 6 | CLTI Right femoral ligation. R: day 10 | H2O2 measurement (by Amplex Red assay) | Significant increase in H2O2 level in ischemic muscles compared with sham ones (p < 0.05). | Kwon et al., 2016, Int J Pharm [43]. |
| Mice, n = 28 | CLTI Sequential right femoral and iliac ligations. R: day 30. | Free radical measurement (by electron paramagnetic resonance spectroscopy) DHE measures of ROS (by epifluorescence microscopy) | CLI induced a significant increase in ROS production in ischemic muscles compared with controls. DHE staining was higher in ischemic muscles (p < 0.01). | Lejay et al., 2015, Eur J Vasc Endovasc Surg [42]. |
| Rats, n = 35 | Aortic banding I: 2 h/R: 10 min and 2 h | DHE staining (by epifluorescence microscopy) | ROS increased significantly after ischemia alone (+324 ± 66%, p = 0.038), normalized after 10 min of reperfusion, and increased again at 2 h of reperfusion (+349.2 ± 67%, p = 0.024). Oxidative stress preceded skeletal muscle mitochondrial dysfunction. | Guillot et al., 2014, J Vasc Surg [48]. |
| Mice, n = 18 | Unilateral tourniquet I: 3 h/R: 4 h | Superoxide anion production measurement (by luminometer); Quantification of MnSOD (by Western blot) | Increased superoxide production and decreased activity of the mitochondria-targeted SOD isoform) in the ischemia-reperfusion group. | Tran et al., 2011, Eur J Pharmacol [45]. |
| Mice, n = 48 | CLTI Sequential left femoral and iliac ligations R: week 12 | Protein carbonyls, HNE adducts and MnSOD expression quantification (by reverse phase protein lysate microarray) | Significantly higher expression of protein carbonyls, HNE adducts and MnSOD in ischemic muscles compared with controls (p < 0.05). | Pipinos et al., 2008, Am J Physiol Regul Integr Comp Physiol [22]. |
| Population | Number Studied (Symptomatic/Controls) | Outcomes Measured | Main Results | Reference |
|---|---|---|---|---|
| Claudicant PAD | 10/34 | Mitochondrial DNA copy number (by quantitative PCR) | Significant association between disease severity and increased mitochondrial DNA copy number (p < 0.05). | McDermott et al., 2018, Vasc Med [49]. |
| Claudicant PAD | 10/12 | Free radical measurement (by electron paramagnetic resonance spectroscopy) | Significant increase in mitochondria-derived ROS production in PAD (1.0 ± 0.36 AU/mg tissue for controls versus 4.3 ± 1.0 AU/mg tissue for PAD, p < 0.05). | Hart et al., 2018, Exp Physiol [32]. |
| Claudicant PAD; CLTI | 28 claudicants/25 CLTI/25 controls | Carbonyl groups quantification (by quantitative fluorescence microscopy) | Observation of a 25% increase in carbonyl groups (markers of oxidative damage) in myofibers of all PAD patients compared with controls (p < 0.05). | Koutakis et al., 2014, Redox Biol [50]. |
| Claudicant PAD; CLTI | 34/21 | Carbonyl groups and HNE adducts quantification (by quantitative fluorescence microscopy) | Significant increase in carbonyl groups (30%, p < 0.0001) and HNE adducts (40%, p < 0.0001) in PAD myofibers compared to controls. | Weiss et al., 2013, J Transl Med [51]. |
| Claudicant PAD; CLTI | 16/10 | Lipid hydroperoxides measurement (by ferrous oxidation/xylenol orange technique); Protein carbonyls measurments (using an Enzyme Immuno-Assay kit); HNE detection (by western blot); Antioxidant activity (by spectrophotometry) | Higher levels of lipid hydroperoxides (12.45 ± 0.74 mmol/g wet weight for controls versus 20.32 ± 1.02 for PAD), protein carbonyls (0.22 ± 0.02 nmol/mg for controls versus 0.35 ± 0.04) and HNE (191.2 ± 7.17 total binding versus 226.4 ± 10.4) was found in PAD patients compared to controls (p < 0.05). Significant decrease in SOD activity, and increase in catalase and glutathione peroxidase activities. | Pipinos et al., 2006, Free Radic Biol Med [28]. |
| Claudicant PAD | 9 claudicants | Quantification of mitochondrial DNA injury (by PCR) | Substantial injury to mitochondrial DNA in PAD patients occurring bilaterally in patients with unilateral PAD. | Brass et al., 2000, Vasc Med [52]. |
| Claudicant PAD | 8/10 | Quantification of mitochondrial DNA injury (by PCR) | Accumulation of 4977-bp mitochondrial deletion frequency in patients with PAD compared with controls (0.05 ± 0.01 % for controls versus 0.43 ± 0.28 % for the less-affected limb versus 0.88 ± 0.53 % for the worse-affected limb, p < 0.05). | Bhat et al., 1999, Circulation [53]. |
| Animals | Study Design Ischemia-Reperfusion Duration | Outcomes Measured | Main Results | Reference |
|---|---|---|---|---|
| Mice, n = 7 ApoE-/- versus ApoE+/+ | CLTI Sequential right femoral and iliac ligations. | Calcium retention capacity (by spectrofluometry) | Impairment in calcium retention capacity in ischemic muscles of ApoE-/- and ApoE+/+ mice compared with contralateral muscles (p = 0.001). | Lejay et al., 2019, Eur J Vasc Endovasc Surg [44]. |
| Mice, n = 25 young (23 weeks) versus aged (84 weeks) C57Bl6J | Unilateral tourniquet I: 2 h/R: 2 h | Calcium retention capacity (by spectrofluometry) | Significant reduction in calcium retention capacity in young (-60.9 ± 7.3%) and aged (-60.9 ± 4.6%) mice compared with sham (p < 0.001). | Paradis et al., 2019, Antioxidants [25]. |
| Rats, n = 12 | CLTI Left femoral ligation. R: day 14 | Protein expression of indicators of apoptosis (by Western blot) | Higher expression of proteins cleaved-caspase 3, cleaved-PARP and mitochondrial Bax in CLTI muscles compared with sham ones (p < 0.05). | Hsu et al., 2019, Am J Transl Res [54]. |
| Mice, n = 16 | Unilateral tourniquet I: 2 h/R: 2 h | Calcium retention capacity (by spectrofluometry) | Decrease in calcium retention capacity in ischemic limbs compared with contralateral ones (-61.1 ± 6.8%, p < 0.01). | Tetsi et al., 2019. Antioxidants [56]. |
| Mice, n = 20 | CLTI Sequential right femoral and iliac ligations. R: day 30. | Calcium retention capacity (by spectrofluometry) | Significant reduction of calcium retention capacity in ischemic limbs compared with contralateral ones (p < 0.001). | Lejay et al., 2018, Eur J Vasc Endovasc Surg [57]. |
| Mice, n = 20 | CLTI Sequential right femoral and iliac ligations. | Calcium retention capacity (by spectrofluometry) | Lower calcium retention capacity in ischemic limbs compared with controls (p < 0.001). | Lejay et al., 2017, Front Physiol [47]. |
| Rats, n = 16 | CLTI Right femoral ligation. R: day 14 | Protein expression of indicators of apoptosis and anti-apoptotic factor (by Western blot) | Higher expression of the proteins cleaved-caspase 3, cleaved-PARP and cytosolic Bax in CLTI muscles compared with sham ones (p < 0.001). Lower expression of the anti-apoptotic marker Bcl-2 in CLTI muscles compared with sham ones (p < 0.001). | Sheu et al., 2015, J Transl Med [55]. |
| Rats n = 28 | Aortic banding I: 3 h/R: 2 h | Quantification of gene expression (by quantitative PCR) | IR increased Bax (63.4%, p = 0.020) and Bax/Bcl-2 ratio (+84.6%, p = 0.029). SODs and GPx messenger RNA were not modified, but glutathione tended to be decreased after IR. | Mansour et al., 2012, J Vasc Surg [21]. |
| Population | Number Studied (Symptomatic/Controls) | Outcomes Measured | Main Results | Reference |
|---|---|---|---|---|
| Claudicant PAD | 130/36 | Caspase activity measurement (by caspase Assay) | No difference observed in apoptosis between the PAD and the control group (p = 0.463). | Gardner et al., 2014, Angiology [62]. |
| Claudicant PAD | 156/16 | Caspase activity measurement (by caspase Assay) | Higher percentage of endothelial cell apoptosis in the PAD group compared with the control group (+164%, p < 0.001). | Gardner et al., 2014, Int J Vasc Med [60]. |
| Claudicant PAD | 19/18 | Quantification of gene expression (by quantitative PCR) | Upregulation in genes mediating apoptosis: BCL-2, G0S2, KLF6, PTP4A1 and CFLAR. | Masud et al., 2012, J Clin Bioinforma [58]. |
| Claudicant PAD | 10/10 | Detection and quantification of apoptosis (by fluorescence microscopy) | Higher percentage of late apoptotic lymphocytes (by 33%) in the PAD patients compared with healthy controls. | Skórkowska-Telichowska et al., 2009, Clin Invest Med [61]. |
| Claudicant PAD | 26/28 | Apoptosis detection (by TUNEL Assay); Caspase activity measurement (by caspase Assay) | The fraction of TUNEL-positive nuclei was greater in PAD patients compared with controls (1.53% ± 0.96 for controls versus 3.83% ± 2.6 for PAD, p < 0.001). Caspase-3 activity was increased in PAD group compared with control group (0.22 ± 0.05 units mg−1 soluble protein versus 0.39 ± 0.09 for PAD, p < 0.001). | Mitchell et al., 2007, Vasc Med [59]. |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pizzimenti, M.; Riou, M.; Charles, A.-L.; Talha, S.; Meyer, A.; Andres, E.; Chakfé, N.; Lejay, A.; Geny, B. The Rise of Mitochondria in Peripheral Arterial Disease Physiopathology: Experimental and Clinical Data. J. Clin. Med. 2019, 8, 2125. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8122125
Pizzimenti M, Riou M, Charles A-L, Talha S, Meyer A, Andres E, Chakfé N, Lejay A, Geny B. The Rise of Mitochondria in Peripheral Arterial Disease Physiopathology: Experimental and Clinical Data. Journal of Clinical Medicine. 2019; 8(12):2125. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8122125
Chicago/Turabian StylePizzimenti, Mégane, Marianne Riou, Anne-Laure Charles, Samy Talha, Alain Meyer, Emmanuel Andres, Nabil Chakfé, Anne Lejay, and Bernard Geny. 2019. "The Rise of Mitochondria in Peripheral Arterial Disease Physiopathology: Experimental and Clinical Data" Journal of Clinical Medicine 8, no. 12: 2125. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm8122125