Impaired Release of Neutrophil Extracellular Traps and Anemia-Associated T Cell Deficiency in Hereditary Hemorrhagic Telangiectasia

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Experimental Section
2.1. Selection of Participants and Clinical Characteristics
2.2. Laboratory and Immunological Parameters of the Whole Blood
2.3. Plasma
2.4. Isolation of Human Neutrophils
2.5. Bacteria
2.6. NET Release
2.7. Reactive Oxygen Species
2.8. F-Actin Detection with Phalloidin
2.9. Microscopy
2.10. Migration to Lipopolysaccharide (LPS)
2.11. ELISA
2.12. Statistical Analysis
2.13. Study Approval
3. Results
3.1. Clinical Characteristics of the HHT Patients
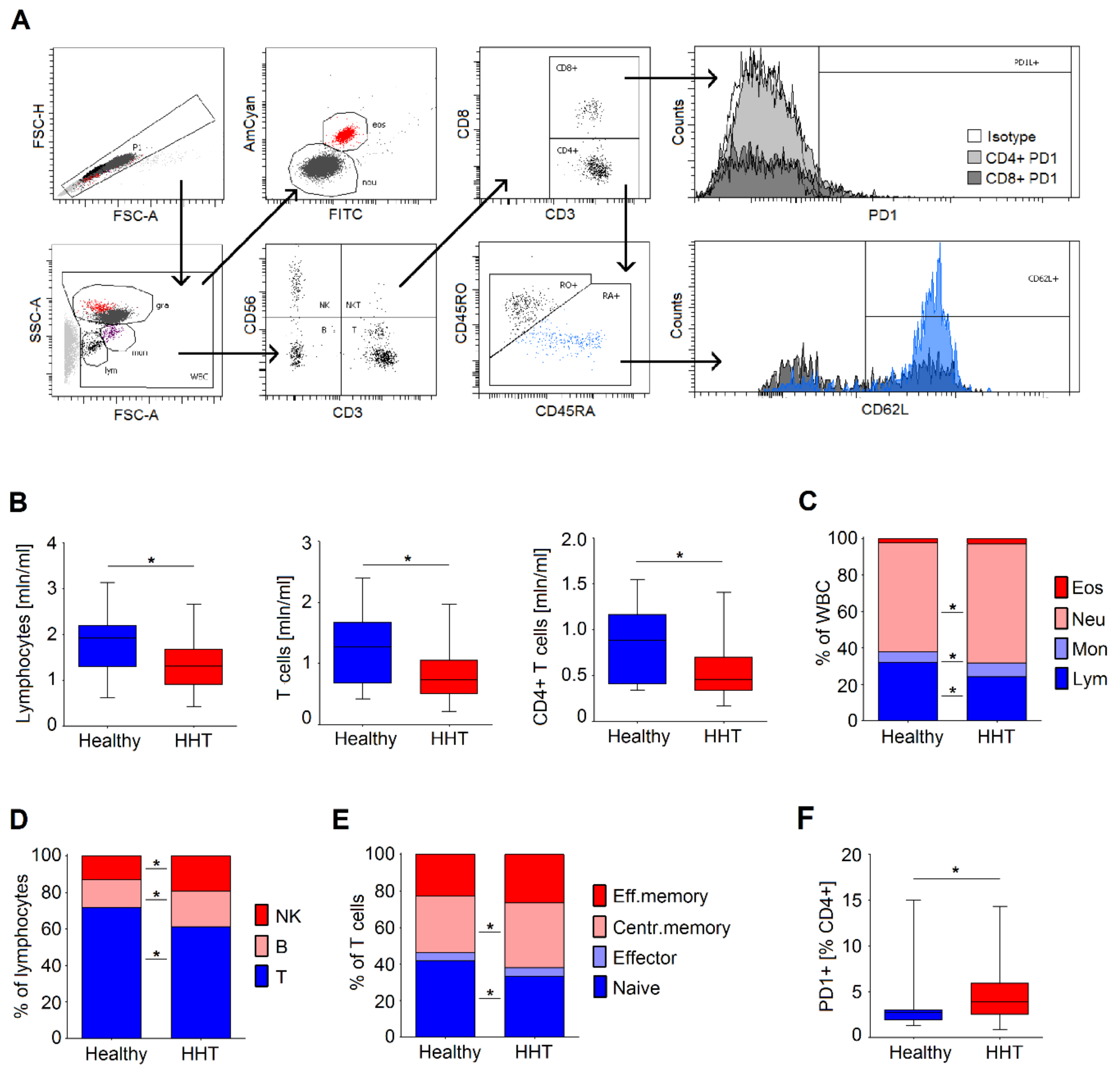
3.2. Immune Status of Healthy and HHT Individuals of Comparable Age
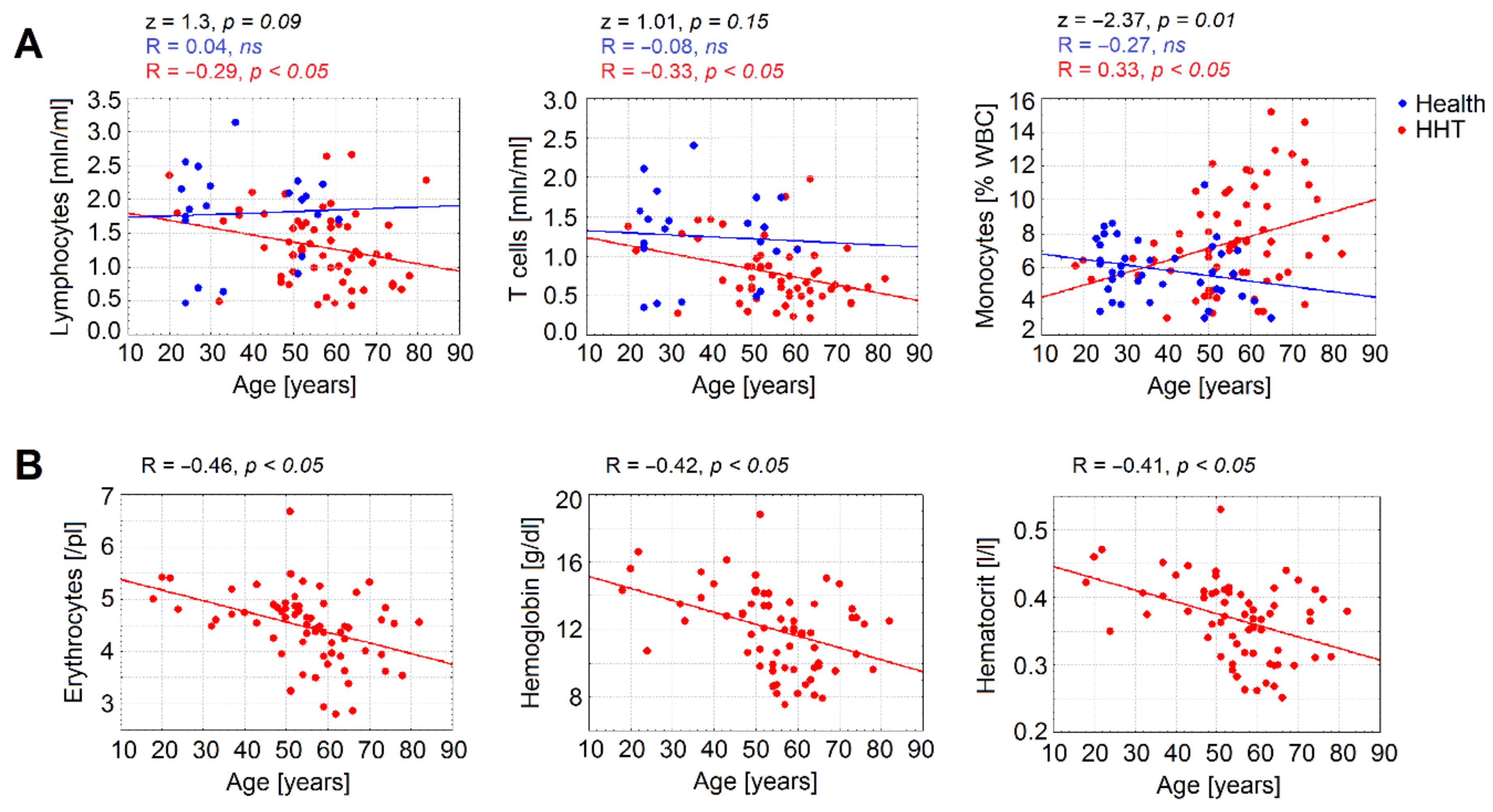
3.3. Age Is a Significant Factor Contributing to Declined Immune Parameters in HHT
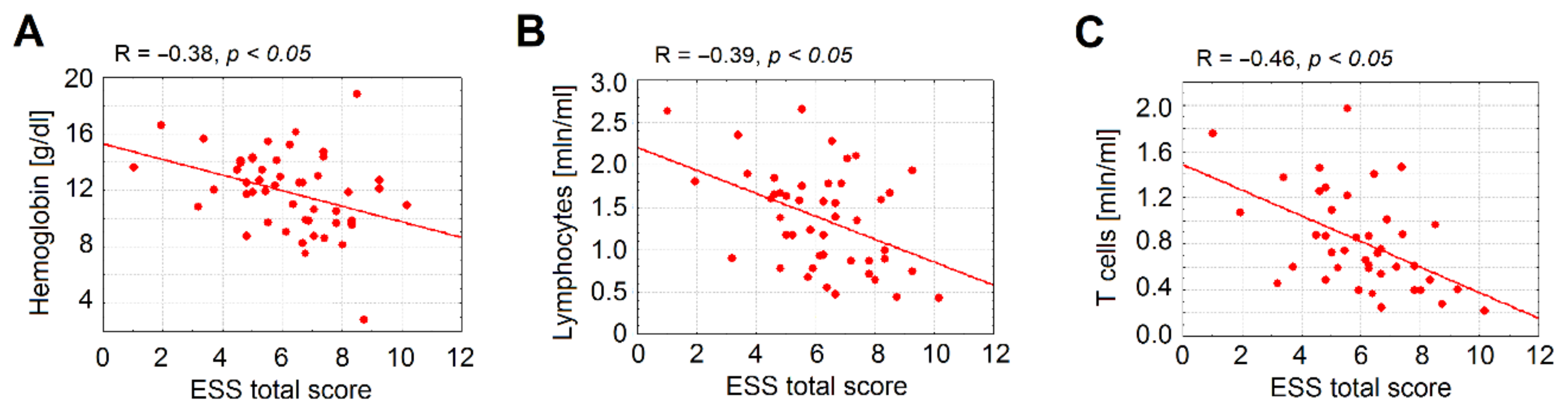
3.4. Anemia Is Associated with a Significant Decrease in Lymphocytes Counts in HHT
3.5. Immune Parameters Correlate with Type of Vascular Malformation in HHT
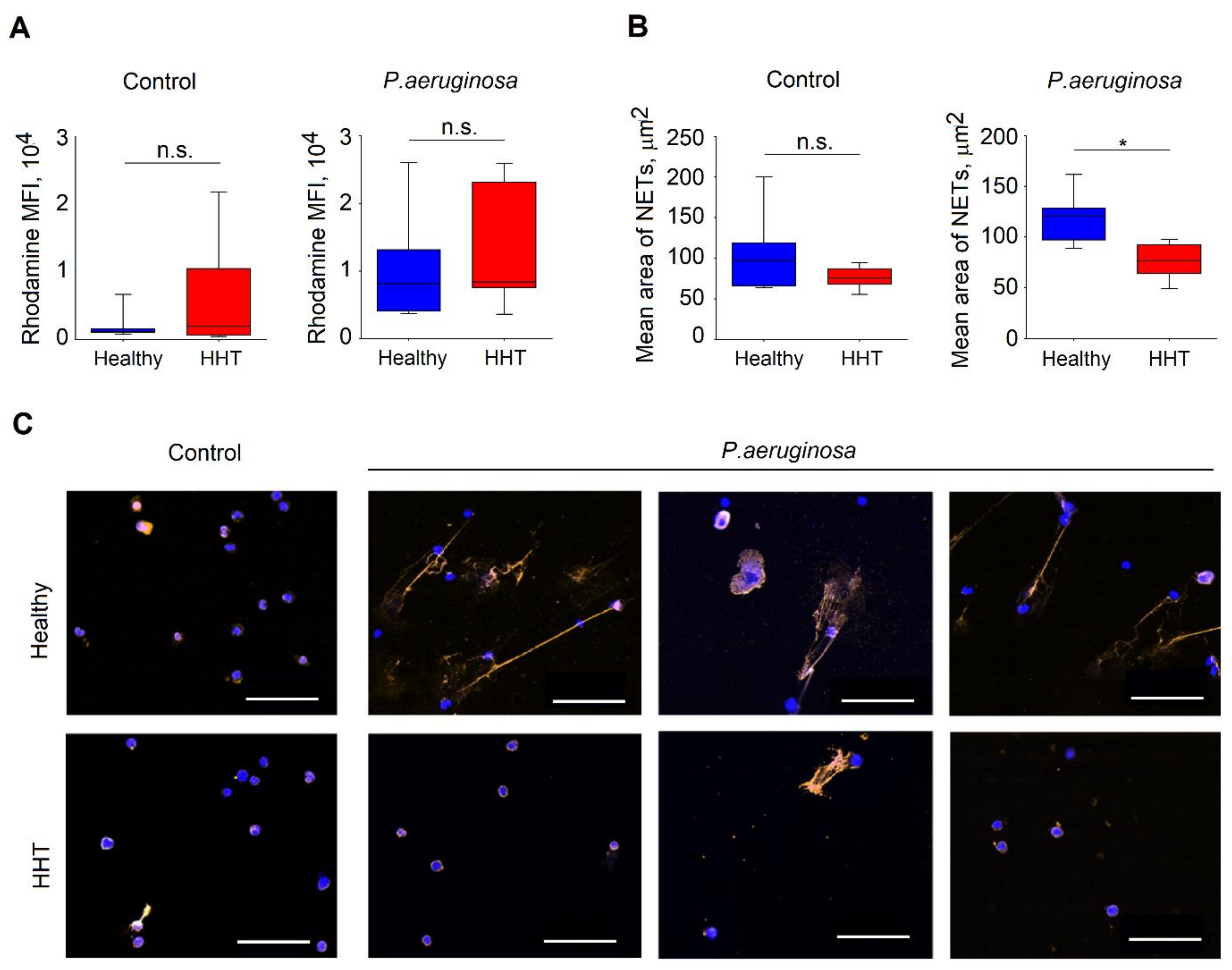
3.6. Reduced Functionality of Neutrophils Isolated From Patients with HHT
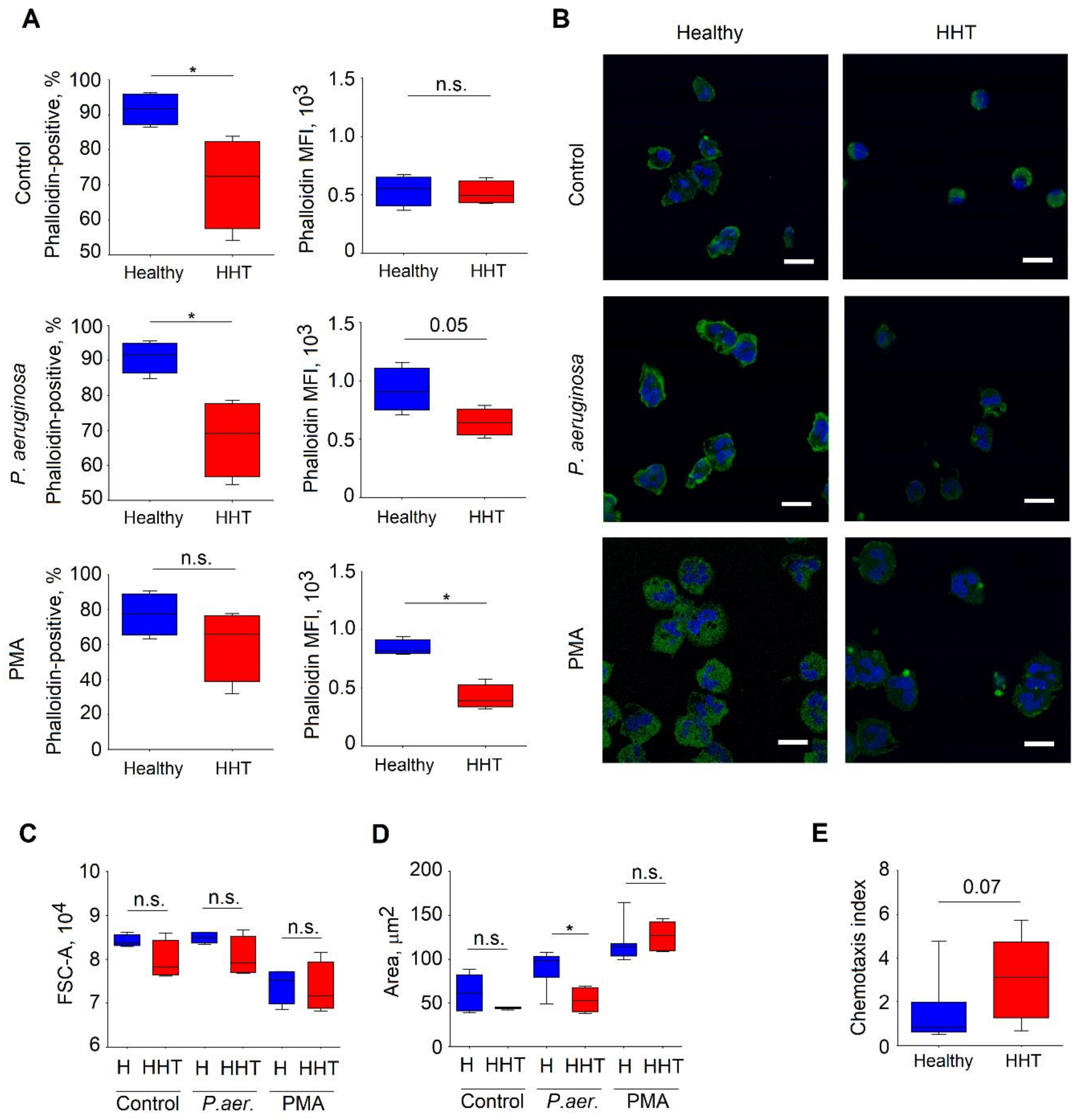
3.7. Altered Cytoskeleton in Neutrophils Isolated From HHT Patients
4. Discussion
Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McAllister, K.A.; Grogg, K.M.; Johnson, D.W.; Gallione, C.J.; Baldwin, M.A.; Jackson, C.E.; Helmbold, E.A.; Markel, D.S.; McKinnon, W.C.; Murrell, J.; et al. Endoglin, a TGF-beta binding protein of endothelial cells, is the gene for hereditary haemorrhagic telangiectasia type 1. Nat. Genet. 1994, 8, 345–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shovlin, C.L.; Hughes, J.M.; Tuddenham, E.G.; Temperley, I.; Perembelon, Y.F.; Scott, J.; Seidman, C.E.; Seidman, J.G. A gene for hereditary haemorrhagic telangiectasia maps to chromosome 9q3. Nat. Genet. 1994, 6, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.W.; Berg, J.N.; Gallione, C.J.; McAllister, K.A.; Warner, J.P.; Helmbold, E.A.; Markel, D.S.; Jackson, C.E.; Porteous, M.E.; Marchuk, D.A. A second locus for hereditary hemorrhagic telangiectasia maps to chromosome 12. Genome Res. 1995, 5, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kjeldsen, A.D.; Vase, P.; Green, A. Hereditary haemorrhagic telangiectasia: A population-based study of prevalence and mortality in Danish patients. J. Intern. Med. 1999, 245, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Gallione, C.J.; Repetto, G.M.; Legius, E.; Rustgi, A.K.; Schelley, S.L.; Tejpar, S.; Mitchell, G.; Drouin, E.; Westermann, C.J.; Marchuk, D.A. A combined syndrome of juvenile polyposis and hereditary haemorrhagic telangiectasia associated with mutations in MADH4 (SMAD4). Lancet 2004, 363, 852–859. [Google Scholar] [CrossRef]
- Cole, S.G.; Begbie, M.E.; Wallace, G.M.; Shovlin, C.L. A new locus for hereditary haemorrhagic telangiectasia (HHT3) maps to chromosome 5. J. Med Genet. 2005, 42, 577–582. [Google Scholar] [CrossRef]
- Finkelstein, R.; Engel, A.; Simri, W.; Hemli, J.A. Brain abscesses: The lung connection. J. Intern. Med. 1996, 240, 33–36. [Google Scholar] [CrossRef]
- Dong, S.L.; Reynolds, S.F.; Steiner, I.P. Brain abscess in patients with hereditary hemorrhagic telangiectasia: Case report and literature review. J. Emerg. Med. 2001, 20, 247–251. [Google Scholar] [CrossRef]
- Gallitelli, M.; Lepore, V.; Pasculli, G.; Di Gennaro, L.; Logroscino, G.; Carella, A.; White, R.I.; Sabba, C. Brain abscess: A need to screen for pulmonary arteriovenous malformations. Neuroepidemiology 2005, 24, 76–78. [Google Scholar] [CrossRef]
- Tabakow, P.; Jarmundowicz, W.; Czapiga, B.; Czapiga, E. Brain abscess as the first clinical manifestation of multiple pulmonary arteriovenous malformations in a patient with hereditary hemorrhagic telangiectasia (Rendu-Osler-Weber disease). Folia Neuropathol. 2005, 43, 41–44. [Google Scholar]
- Dupuis-Girod, S.; Giraud, S.; Decullier, E.; Lesca, G.; Cottin, V.; Faure, F.; Merrot, O.; Saurin, J.C.; Cordier, J.F.; Plauchu, H. Hemorrhagic hereditary telangiectasia (Rendu-Osler disease) and infectious diseases: An underestimated association. Clin. Infect. Dis. 2007, 44, 841–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, P.; Schaeverbeke, T.; Baillet, L.; Lequen, L.; Bannwarth, B.; Dehais, J. Rendu-Osler familial telangiectasia angiomatosis and bacterial spondylodiscitis. Rev. Med. Interne 1998, 19, 938–939. [Google Scholar] [CrossRef]
- Duval, X.; Djendli, S.; Le Moing, V.; Longuet, P.; Barry, B.; Leport, C.; Vilde, J.L. Recurrent Staphylococcus aureus extracerebral infections complicating hereditary hemorrhagic telangiectasia (Osler-Rendu-Weber disease). Am. J. Med. 2001, 110, 671–672. [Google Scholar] [CrossRef]
- Shovlin, C.L.; Jackson, J.E.; Bamford, K.B.; Jenkins, I.H.; Benjamin, A.R.; Ramadan, H.; Kulinskaya, E. Primary determinants of ischaemic stroke/brain abscess risks are independent of severity of pulmonary arteriovenous malformations in hereditary haemorrhagic telangiectasia. Thorax 2008, 63, 259–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habib, G.; Lancellotti, P.; Antunes, M.J.; Bongiorni, M.G.; Casalta, J.P.; Del Zotti, F.; Dulgheru, R.; El Khoury, G.; Erba, P.A.; Iung, B.; et al. 2015 ESC Guidelines for the management of infective endocarditis: The Task Force for the Management of Infective Endocarditis of the European Society of Cardiology (ESC). Endorsed by: European Association for Cardio-Thoracic Surgery (EACTS), the European Association of Nuclear Medicine (EANM). Eur. Heart J. 2015, 36, 3075–3128. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Abdelmohsen, K.; Abe, A.; Abedin, M.J.; Abeliovich, H.; Acevedo Arozena, A.; Adachi, H.; Adams, C.M.; Adams, P.D.; Adeli, K.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 2016, 12, 1–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinauer, M.C. Disorders of neutrophil function: An overview. Methods Mol. Biol. 2014, 1124, 501–515. [Google Scholar] [CrossRef]
- Kaplan, M.J.; Radic, M. Neutrophil extracellular traps: Double-edged swords of innate immunity. J. Immunol. 2012, 189, 2689–2695. [Google Scholar] [CrossRef] [Green Version]
- Vulcano, M.; Dusi, S.; Lissandrini, D.; Badolato, R.; Mazzi, P.; Riboldi, E.; Borroni, E.; Calleri, A.; Donini, M.; Plebani, A.; et al. Toll receptor-mediated regulation of NADPH oxidase in human dendritic cells. J. Immunol. 2004, 173, 5749–5756. [Google Scholar] [CrossRef] [Green Version]
- Mantegazza, A.R.; Savina, A.; Vermeulen, M.; Perez, L.; Geffner, J.; Hermine, O.; Rosenzweig, S.D.; Faure, F.; Amigorena, S. NADPH oxidase controls phagosomal pH and antigen cross-presentation in human dendritic cells. Blood 2008, 112, 4712–4722. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, M.; Niemiec, M.J.; Siler, U.; Urban, C.F.; Reichenbach, J. Restoration of anti-Aspergillus defense by neutrophil extracellular traps in human chronic granulomatous disease after gene therapy is calprotectin-dependent. J. Allergy Clin. Immunol. 2011, 127, 1243–1252.e7. [Google Scholar] [CrossRef] [PubMed]
- Abi Abdallah, D.S.; Lin, C.; Ball, C.J.; King, M.R.; Duhamel, G.E.; Denkers, E.Y. Toxoplasma gondii triggers release of human and mouse neutrophil extracellular traps. Infect. Immun. 2012, 80, 768–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Shovlin, C.L.; Guttmacher, A.E.; Buscarini, E.; Faughnan, M.E.; Hyland, R.H.; Westermann, C.J.; Kjeldsen, A.D.; Plauchu, H. Diagnostic criteria for hereditary hemorrhagic telangiectasia (Rendu-Osler-Weber syndrome). Am. J. Med. Genet. 2000, 91, 66–67. [Google Scholar] [CrossRef]
- Faughnan, M.E.; Palda, V.A.; Garcia-Tsao, G.; Geisthoff, U.W.; McDonald, J.; Proctor, D.D.; Spears, J.; Brown, D.H.; Buscarini, E.; Chesnutt, M.S.; et al. International guidelines for the diagnosis and management of hereditary haemorrhagic telangiectasia. J. Med Genet. 2011, 48, 73–87. [Google Scholar] [CrossRef]
- Hoag, J.B.; Terry, P.; Mitchell, S.; Reh, D.; Merlo, C.A. An epistaxis severity score for hereditary hemorrhagic telangiectasia. Laryngoscope 2010, 120, 838–843. [Google Scholar] [CrossRef]
- National Heart, Lung and Blood Institute. Lymphocytopenia. Available online: https://www.nhlbi.nih.gov/health-topics/lymphocytopenia (accessed on 19 February 2019).
- WHO. Haemoglobin concentrations for the diagnosis of anaemia and assessment of severity. In Vitamin and Mineral Nutrition Information System; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Cherayil, B.J. Iron and immunity: Immunological consequences of iron deficiency and overload. Arch. Immunol. Ther. Exp. 2010, 58, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Neeli, I.; Dwivedi, N.; Khan, S.; Radic, M. Regulation of extracellular chromatin release from neutrophils. J. Innate Immun. 2009, 1, 194–201. [Google Scholar] [CrossRef]
- Tzur, A.; Moore, J.K.; Jorgensen, P.; Shapiro, H.M.; Kirschner, M.W. Optimizing optical flow cytometry for cell volume-based sorting and analysis. PLoS ONE 2011, 6, e16053. [Google Scholar] [CrossRef]
- McDonald, J.; Wooderchak-Donahue, W.; VanSant Webb, C.; Whitehead, K.; Stevenson, D.A.; Bayrak-Toydemir, P. Hereditary hemorrhagic telangiectasia: Genetics and molecular diagnostics in a new era. Front. Genet. 2015, 6, 1. [Google Scholar] [CrossRef]
- Shovlin, C.L.; Condliffe, R.; Donaldson, J.W.; Kiely, D.G.; Wort, S.J.; British Thoracic, S. British Thoracic Society Clinical Statement on Pulmonary Arteriovenous Malformations. Thorax 2017, 72, 1154–1163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Samkari, H.; Kritharis, A.; Kuter, D.J. Infections and vaccination in hereditary hemorrhagic telangiectasia: Microbiological evidence-based considerations. Haematologica 2018, 103, e492–e495. [Google Scholar] [CrossRef] [PubMed]
- Kjeldsen, A.D.; Oxhoj, H.; Andersen, P.E.; Green, A.; Vase, P. Prevalence of pulmonary arteriovenous malformations (PAVMs) and occurrence of neurological symptoms in patients with hereditary haemorrhagic telangiectasia (HHT). J. Intern. Med. 2000, 248, 255–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amati, L.; Passeri, M.E.; Resta, F.; Triggiani, V.; Jirillo, E.; Sabba, C. Ablation of T-helper 1 cell derived cytokines and of monocyte-derived tumor necrosis factor-alpha in hereditary hemorrhagic telangiectasia: Immunological consequences and clinical considerations. Curr. Pharm. Des. 2006, 12, 1201–1208. [Google Scholar] [PubMed]
- Guilhem, A.; Malcus, C.; Clarivet, B.; Plauchu, H.; Dupuis-Girod, S. Immunological abnormalities associated with hereditary haemorrhagic telangiectasia. J. Intern. Med. 2013, 274, 351–362. [Google Scholar] [CrossRef]
- Sallusto, F.; Geginat, J.; Lanzavecchia, A. Central memory and effector memory T cell subsets: Function, generation, and maintenance. Annu. Rev. Immunol. 2004, 22, 745–763. [Google Scholar] [CrossRef]
- Plauchu, H.; de Chadarevian, J.P.; Bideau, A.; Robert, J.M. Age-related clinical profile of hereditary hemorrhagic telangiectasia in an epidemiologically recruited population. Am. J. Med Genet. 1989, 32, 291–297. [Google Scholar] [CrossRef]
- Lenato, G.M.; Suppressa, P.; Giordano, P.; Guanti, G.; Guastamacchia, E.; Triggiani, V.; Amati, L.; Resta, F.; Covelli, V.; Jirillo, E.; et al. Hereditary haemorrhagic telangiectasia: A rare disease as a model for the study of human atherosclerosis. Curr. Pharm. Des. 2007, 13, 3656–3664. [Google Scholar] [CrossRef]
- Catakovic, K.; Klieser, E.; Neureiter, D.; Geisberger, R. T cell exhaustion: From pathophysiological basics to tumor immunotherapy. Cell Commun. Signal. 2017, 15, 1. [Google Scholar] [CrossRef] [Green Version]
- Franceschi, C.; Capri, M.; Monti, D.; Giunta, S.; Olivieri, F.; Sevini, F.; Panourgia, M.P.; Invidia, L.; Celani, L.; Scurti, M.; et al. Inflammaging and anti-inflammaging: A systemic perspective on aging and longevity emerged from studies in humans. Mech. Ageing Dev. 2007, 128, 92–105. [Google Scholar] [CrossRef]
- Fulop, T.; Dupuis, G.; Witkowski, J.M.; Larbi, A. The Role of Immunosenescence in the Development of Age-Related Diseases. Rev. Investig. Clin. 2016, 68, 84–91. [Google Scholar] [PubMed]
- Wang, Y.; Wehling-Henricks, M.; Welc, S.S.; Fisher, A.L.; Zuo, Q.; Tidball, J.G. Aging of the immune system causes reductions in muscle stem cell populations, promotes their shift to a fibrogenic phenotype, and modulates sarcopenia. FASEB J. 2018. [Google Scholar] [CrossRef] [PubMed]
- Wack, A.; Cossarizza, A.; Heltai, S.; Barbieri, D.; D’Addato, S.; Fransceschi, C.; Dellabona, P.; Casorati, G. Age-related modifications of the human alphabeta T cell repertoire due to different clonal expansions in the CD4+ and CD8+ subsets. Int. Immunol. 1998, 10, 1281–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franceschi, C.; Bonafe, M.; Valensin, S. Human immunosenescence: The prevailing of innate immunity, the failing of clonotypic immunity, and the filling of immunological space. Vaccine 2000, 18, 1717–1720. [Google Scholar] [CrossRef]
- Ekiz, C.; Agaoglu, L.; Karakas, Z.; Gurel, N.; Yalcin, I. The effect of iron deficiency anemia on the function of the immune system. Hematol. J. 2005, 5, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Hassan, T.H.; Badr, M.A.; Karam, N.A.; Zkaria, M.; El Saadany, H.F.; Abdel Rahman, D.M.; Shahbah, D.A.; Al Morshedy, S.M.; Fathy, M.; Esh, A.M.; et al. Impact of iron deficiency anemia on the function of the immune system in children. Medicine 2016, 95, e5395. [Google Scholar] [CrossRef]
- Boyer, S.W.; Rajendiran, S.; Beaudin, A.E.; Smith-Berdan, S.; Muthuswamy, P.K.; Perez-Cunningham, J.; Martin, E.W.; Cheung, C.; Tsang, H.; Landon, M.; et al. Clonal and Quantitative In Vivo Assessment of Hematopoietic Stem Cell Differentiation Reveals Strong Erythroid Potential of Multipotent Cells. Stem Cell Rep. 2019, 12, 801–815. [Google Scholar] [CrossRef] [Green Version]
- Ojeda-Fernandez, L.; Recio-Poveda, L.; Aristorena, M.; Lastres, P.; Blanco, F.J.; Sanz-Rodriguez, F.; Gallardo-Vara, E.; de las Casas-Engel, M.; Corbi, A.; Arthur, H.M.; et al. Mice Lacking Endoglin in Macrophages Show an Impaired Immune Response. PLoS Genet. 2016, 12, e1005935. [Google Scholar] [CrossRef] [Green Version]
- Guilhem, A. T lymphocyte changes and iron supplementation in HHT: A physiopathological link? In Proceedings of the 12th International HHT Scientific Conference, Dubrovnik, Croatia, 8–11 June 2017. [Google Scholar]
- Tanimura, Y.; Shimizu, K.; Tanabe, K.; Otsuki, T.; Yamauchi, R.; Matsubara, Y.; Iemitsu, M.; Maeda, S.; Ajisaka, R. Exercise-induced oxidative DNA damage and lymphocytopenia in sedentary young males. Med. Sci. Sports Exerc. 2008, 40, 1455–1462. [Google Scholar] [CrossRef]
- Tiwari, A.K.; Mahdi, A.A.; Chandyan, S.; Zahra, F.; Godbole, M.M.; Jaiswar, S.P.; Srivastava, V.K.; Negi, M.P. Oral iron supplementation leads to oxidative imbalance in anemic women: A prospective study. Clin. Nutr. 2011, 30, 188–193. [Google Scholar] [CrossRef]
- Cirulli, A.; Loria, M.P.; Dambra, P.; Di Serio, F.; Ventura, M.T.; Amati, L.; Jirillo, E.; Sabba, C. Patients with Hereditary Hemorrhagic Telangectasia (HHT) exhibit a deficit of polymorphonuclear cell and monocyte oxidative burst and phagocytosis: A possible correlation with altered adaptive immune responsiveness in HHT. Curr. Pharm. Des. 2006, 12, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Stojkov, D.; Amini, P.; Oberson, K.; Sokollik, C.; Duppenthaler, A.; Simon, H.U.; Yousefi, S. ROS and glutathionylation balance cytoskeletal dynamics in neutrophil extracellular trap formation. J. Cell Biol. 2017, 216, 4073–4090. [Google Scholar] [CrossRef] [PubMed]
- Segal, B.H.; Veys, P.; Malech, H.; Cowan, M.J. Chronic granulomatous disease: Lessons from a rare disorder. Biol. Blood Marrow Transpl. 2011, 17, S123–S131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, L.A.; Sanz-Rodriguez, F.; Blanco, F.J.; Bernabeu, C.; Botella, L.M. Hereditary hemorrhagic telangiectasia, a vascular dysplasia affecting the TGF-beta signaling pathway. Clin. Med. Res. 2006, 4, 66–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz-Rodriguez, F.; Guerrero-Esteo, M.; Botella, L.M.; Banville, D.; Vary, C.P.; Bernabeu, C. Endoglin regulates cytoskeletal organization through binding to ZRP-1, a member of the Lim family of proteins. J. Biol. Chem. 2004, 279, 32858–32868. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Llorente, L.; Gallardo-Vara, E.; Rossi, E.; Smadja, D.M.; Botella, L.M.; Bernabeu, C. Endoglin and alk1 as therapeutic targets for hereditary hemorrhagic telangiectasia. Expert Opin. Ther. Targets 2017, 21, 933–947. [Google Scholar] [CrossRef] [Green Version]
- Sanz-Rodriguez, F.; Fernandez, L.A.; Zarrabeitia, R.; Perez-Molino, A.; Ramirez, J.R.; Coto, E.; Bernabeu, C.; Botella, L.M. Mutation analysis in Spanish patients with hereditary hemorrhagic telangiectasia: Deficient endoglin up-regulation in activated monocytes. Clin. Chem. 2004, 50, 2003–2011. [Google Scholar] [CrossRef]
- Schmidt-Weber, C.B.; Letarte, M.; Kunzmann, S.; Ruckert, B.; Bernabeu, C.; Blaser, K. TGF-{beta} signaling of human T cells is modulated by the ancillary TGF-{beta} receptor endoglin. Int. Immunol. 2005, 17, 921–930. [Google Scholar] [CrossRef]
- Zhang, L.; Magli, A.; Catanese, J.; Xu, Z.; Kyba, M.; Perlingeiro, R.C. Modulation of TGF-beta signaling by endoglin in murine hemangioblast development and primitive hematopoiesis. Blood 2011, 118, 88–97. [Google Scholar] [CrossRef] [Green Version]
- Fuhrmann, A.; Engler, A.J. The cytoskeleton regulates cell attachment strength. Biophys J. 2015, 109, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Cleghorn, W.M.; Branch, K.M.; Kook, S.; Arnette, C.; Bulus, N.; Zent, R.; Kaverina, I.; Gurevich, E.V.; Weaver, A.M.; Gurevich, V.V. Arrestins regulate cell spreading and motility via focal adhesion dynamics. Mol. Biol. Cell 2015, 26, 622–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.Y.; Santoso, S.; Chavakis, T. Mechanisms of neutrophil transendothelial migration. Front. Biosci. (Landmark Ed.) 2009, 14, 1596–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All Patients (n = 79) | Men (n = 35) | Women (n = 44) | |||
|---|---|---|---|---|---|
| age | years | 55 ± 13 | 56 ± 14 | 54 ± 13 | |
| missing | 0 | 0 | 0 | ||
| genetic testing | yes | 17 (22) | 6 (17) | 11 (25) | |
| no | 62 (78) | 29 (83) | 33 (75) | ||
| missing | 0 | 0 | 0 | ||
| genetic mutation | HHT 1 | 7 (54) | 1 (33) | 6 (60) | |
| HHT 2 | 5 (39) | 2 (67) | 3 (30) | ||
| no mutation detected | 1 (8) | 0 | 1 (10) | ||
| missing | 59 | 32 | 34 | ||
| positive FH | yes | 68 (92) | 30 (91) | 38 (93) | |
| no | 6 (8) | 3 (9) | 3 (7) | ||
| missing | 5 | 2 | 3 | ||
| TAE | yes | 71 (98) | 34 (97) | 43 (98) | |
| no | 2 (2) | 1 (3) | 1 (2) | ||
| missing | 0 | 0 | 0 | ||
| visceral lesions | GI | yes | 27 (49) | 12 (50) | 15 (48) |
| no | 28 (51) | 12 (50) | 16 (52) | ||
| missing | 24 | 11 | 13 | ||
| PAVM | yes | 21 (37) | 7 (30) | 14 (41) | |
| no | 36 (63) | 16 (70) | 20 (59) | ||
| missing | 22 | 12 | 10 | ||
| HVM | yes | 15 (33) | 4 (20) | 11 (42) | |
| no | 31 (67) | 16 (80) | 15 (58) | ||
| missing | 33 | 15 | 18 | ||
| CVM | yes | 6 (11) | 3 (13) | 3 (10) | |
| no | 48 (89) | 20 (87) | 28 (90) | ||
| missing | 25 | 12 | 13 | ||
| epistaxis | yes | 77 (98) | 34 (97) | 43 (98) | |
| no | 2 (2) | 1 (3) | 1 (2) | ||
| missing | 0 | 0 | 0 | ||
| ESS | score | 6 ± 2 | 6 ± 2 | 6 ± 2 | |
| missing | 18 | 6 | 12 | ||
| course of epistaxis | worse | 57 (75) | 24 (69) | 33 (81) | |
| equal | 10 (13) | 5 (14) | 5 (12) | ||
| better | 9 (12) | 6 (17) | 3 (7) | ||
| missing | 3 | 0 | 3 | ||
| course of disease | worse | 61 (81) | 27 (82) | 34 (81) | |
| equal | 12 (16) | 4 (12) | 8 (19) | ||
| better | 2 (3) | 2 (6) | 0 (0) | ||
| missing | 4 | 2 | 2 | ||
| iron intake | yes | 47 (60) | 18 (51) | 29 (66) | |
| no | 32 (40) | 17 (49) | 15 (34) | ||
| missing | 0 | 0 | 0 | ||
| type of iron intake | orally | 22 (53) | 8 (44) | 14 (48) | |
| i.v. | 25 (47) | 10 (66) | 15 (52) | ||
| missing | 32 | 17 | 15 | ||
| smoking habits | smoker | 12 (17) | 8 (26) | 4 (10) | |
| ex-smoker | 18 (26) | 10 (32) | 8 (21) | ||
| non-smoker | 40 (57) | 13 (42) | 27 (69) | ||
| missing | 9 | 4 | 5 | ||
| pack years | number of years | 8 ± 14 | 13 ± 17 | 4 ± 9 | |
| missing | 23 | 11 | 12 | ||
| abscess | yes | 6 (8) | 4 (11) | 2 (5) | |
| no | 73 (92) | 31 (89) | 42 (95) | ||
| missing | 0 | 0 | 0 | ||
| yes | 7 (9) | 1 (3) | 6 (14) | ||
| stroke/embolism | no | 72 (91) | 34 (97) | 38 (86) | |
| missing | 0 | 0 | 0 | ||
| CIDT | yes | 18 (23) | 8 (23) | 10 (23) | |
| no | 61 (77) | 27 (77) | 34 (77) | ||
| missing | 0 | 0 | 0 | ||
| Healthy (n = 23) | HHT (n = 58) | p | ||
|---|---|---|---|---|
| Age | years | 50 (36–53) | 53 (48–59) | 0.08 |
| Gender, male | n, % | 8 (35%) | 24 (41%) | 0.68 |
| Current smoking | n, % | 5 (22%) | 10 (17%) | 0.88 |
| WBC | 106/mL | 6.6 (5.7–8.6) | 5.7 (4.6–6.7) | 0.10 |
| Neutrophils | % WBC 106/mL | 57 (51–68) 3.8 (2.9–6.0) | 63 (57–72) 3.4 (2.6–4.5) | 0.03 0.42 |
| Monocytes | % WBC 106/mL | 5.5 (4.3–7) 0.43 (0.32–0.50) | 6.8 (5.3–8.6) 0.38 (0.32–0.47) | 0.03 0.51 |
| Lymphocytes | % WBC 106/mL | 32 (25–38) 1.9 (1.4–2.1) | 22 (17–28) 1.3 (0.9–1.7) | 0.003 0.02 |
| B cells | % lym 106/mL | 14 (11–16) 0.30 (0.18–0.38) | 17 (13–25) 0.21 (0.13–0.33) | 0.04 0.23 |
| NK cells | % lym 106/mL | 12 (10–14) 0.19 (0.14–0.23) | 17 (14–21) 0.22 (0.14–0.28) | 0.002 0.52 |
| T cells | % lym 106/mL | 70 (66–77) 1.3 (0.8–1.6) | 61 (51–68) 0.7 (0.5–1.1) | 0.0008 0.03 |
| CD4+ | % T cells 106/mL | 76 (69–82) 0.88 (0.41–1.14) | 76 (66–81) 0.47 (0.35–0.70) | 0.72 0.03 |
| Central memory | % T cells | 32 (25–36) | 36 (32–40) | 0.03 |
| Effector memory | % T cells | 20 (19–27) | 26 (20–32) | 0.18 |
| Naive | % T cells | 42 (39–48) | 30 (25–41) | 0.01 |
| Effector | % T cells | 3.0 (2.4–4.3) | 3.0 (2.2–6.0) | 1.00 |
| PD1+ | % CD4+ % CD8+ | 2.7 (2.0-3.0) 2.9 (1.8–3.6) | 3.9 (2.6–5.7) 3.8 (1.8–6) | 0.01 0.32 |
| Plasma TGFß | pg/mL | 13 (11–15) | 12 (11–13) | 0.10 |
| Erythrocytes /pL | Hemoglobing /dL | Hematocrit L/L | Iron µ/dL | ||
|---|---|---|---|---|---|
| WBC | 106/mL | 0.24 | 0.24 | 0.23 | 0.26 |
| Neutrophils | % WBC 106/mL | −0.20 0.19 | −0.18 0.22 | −0.10 0.22 | 0.02 0.32 |
| Monocytes | % WBC 106/mL | −0.19 0.06 | −0.28 −0.00 | −0.26 −0.03 | −0.21 0.14 |
| Lymphocytes | % WBC 106/mL | 0.22 0.33 | 0.21 0.38 | 0.13 0.28 | −0.03 0.14 |
| B cells | % lym 106/mL | 0.12 0.35 | −0.01 0.33 | 0.04 0.29 | −0.20 −0.00 |
| NK cells | % lym 106/mL | −0.21 0.15 | −0.17 0.23 | −0.12 0.22 | 0.01 0.21 |
| T cells | % lym 106/mL | 0.03 0.30 | 0.13 0.39 | 0.05 0.28 | 0.16 0.20 |
| CD4+ | % T cells 106/mL | 0.09 0.27 | −0.05 0.26 | −0.05 0.16 | −0.20 −0.02 |
| 1. No (n = 29) | 2. Oral (n = 22) | 3. I.v. (n = 21) | p | ||
|---|---|---|---|---|---|
| Erythrocytes | /pL | 4.8 (4.5–5.1) | 4.5 (4.0–4.8) | 4.2 (3.6–4.5) | p1/2 = 0.04 p1/3 = 0.0003 |
| Hemoglobin | g/dL | 13.1 (10.6–14.5) | 12.2 (11.2–13.3) | 9.7 (8.7–11.7) | p2/3 = 0.008 p1/3 = 0.004 |
| Hematocrit | L/L | 0.40 (0.35–0.43) | 0.37 (0.36–0.40) | 0.31 (0.30–0.36) | p2/3 = 0.02 p1/3= 0.004 |
| MCHC | g/dL | 33 (31–34) | 33 (32–34) | 31 (30–33) | 0.08 |
| Plasma iron | µg/dL | 53 (20–78) | 35 (25–62) | 27 (16–42) | 0.18 |
| WBC | 106/mL | 5.8 (4.8–7.0) | 5.0 (4.5–5.7) | 5.7 (4.6–6.8) | 0.23 |
| Neutrophils | 106/mL | 3.8 (2.8–4.4) | 2.8 (2.4–4.4) | 3.4 (2.7–4.6) | 0.48 |
| Monocytes | 106/mL | 0.39 (0.32–0.57) | 0.39 (0.32–0.50) | 0.36 (0.26–0.54) | 0.73 |
| Lymphocytes | 106/mL | 1.4 (0.9–1.8) | 1.2 (0.9–1.6) | 1.2 (0.8–1.6) | 0.56 |
| T cells | 106/mL | 0.74 (0.49–1.22) | 0.71 (0.58–1.09) | 0.63 (0.49–0.87) | 0.66 |
| Healthy (n = 7) | HHT (n = 9) | p | ||
|---|---|---|---|---|
| Age | years | 55 (42–62) | 52 (45–67) | 0.87 |
| Gender, male | n (%) | 2 (29%) | 3 (33%) | 0.73 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Droege, F.; Pylaeva, E.; Siakaeva, E.; Bordbari, S.; Spyra, I.; Thangavelu, K.; Lueb, C.; Domnich, M.; Lang, S.; Geisthoff, U.; et al. Impaired Release of Neutrophil Extracellular Traps and Anemia-Associated T Cell Deficiency in Hereditary Hemorrhagic Telangiectasia. J. Clin. Med. 2020, 9, 767. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm9030767
Droege F, Pylaeva E, Siakaeva E, Bordbari S, Spyra I, Thangavelu K, Lueb C, Domnich M, Lang S, Geisthoff U, et al. Impaired Release of Neutrophil Extracellular Traps and Anemia-Associated T Cell Deficiency in Hereditary Hemorrhagic Telangiectasia. Journal of Clinical Medicine. 2020; 9(3):767. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm9030767
Chicago/Turabian StyleDroege, Freya, Ekaterina Pylaeva, Elena Siakaeva, Sharareh Bordbari, Ilona Spyra, Kruthika Thangavelu, Carolin Lueb, Maksim Domnich, Stephan Lang, Urban Geisthoff, and et al. 2020. "Impaired Release of Neutrophil Extracellular Traps and Anemia-Associated T Cell Deficiency in Hereditary Hemorrhagic Telangiectasia" Journal of Clinical Medicine 9, no. 3: 767. https://0-doi-org.brum.beds.ac.uk/10.3390/jcm9030767