
Different Ectopic Hoxa2 Expression Levels in Mouse Cranial Neural Crest Cells Result in Distinct Craniofacial Anomalies and Homeotic Phenotypes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Generation of the ROSA26CAG::(lox-stop-lox)-Hoxa2-IRES-EGFP-WPRE Mouse Line
2.3. Mouse Lines and Mating Scheme
2.4. Skeletal Staining
2.5. RT-qPCR Analysis of mRNA Expression Levels
3. Results
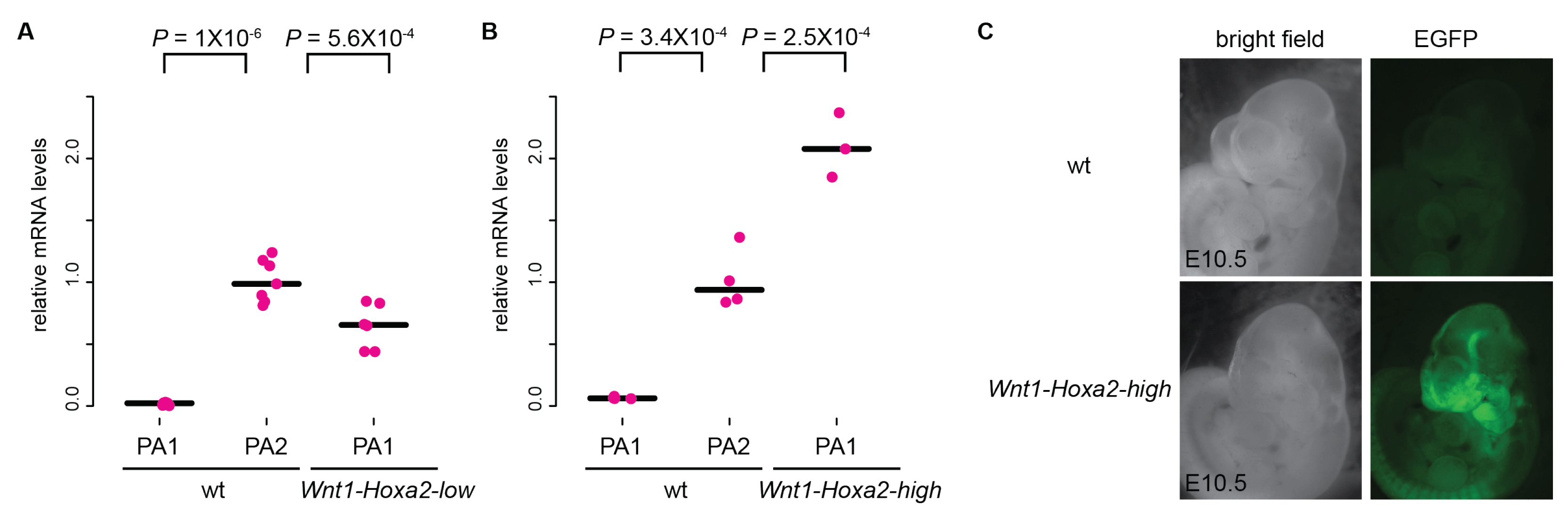
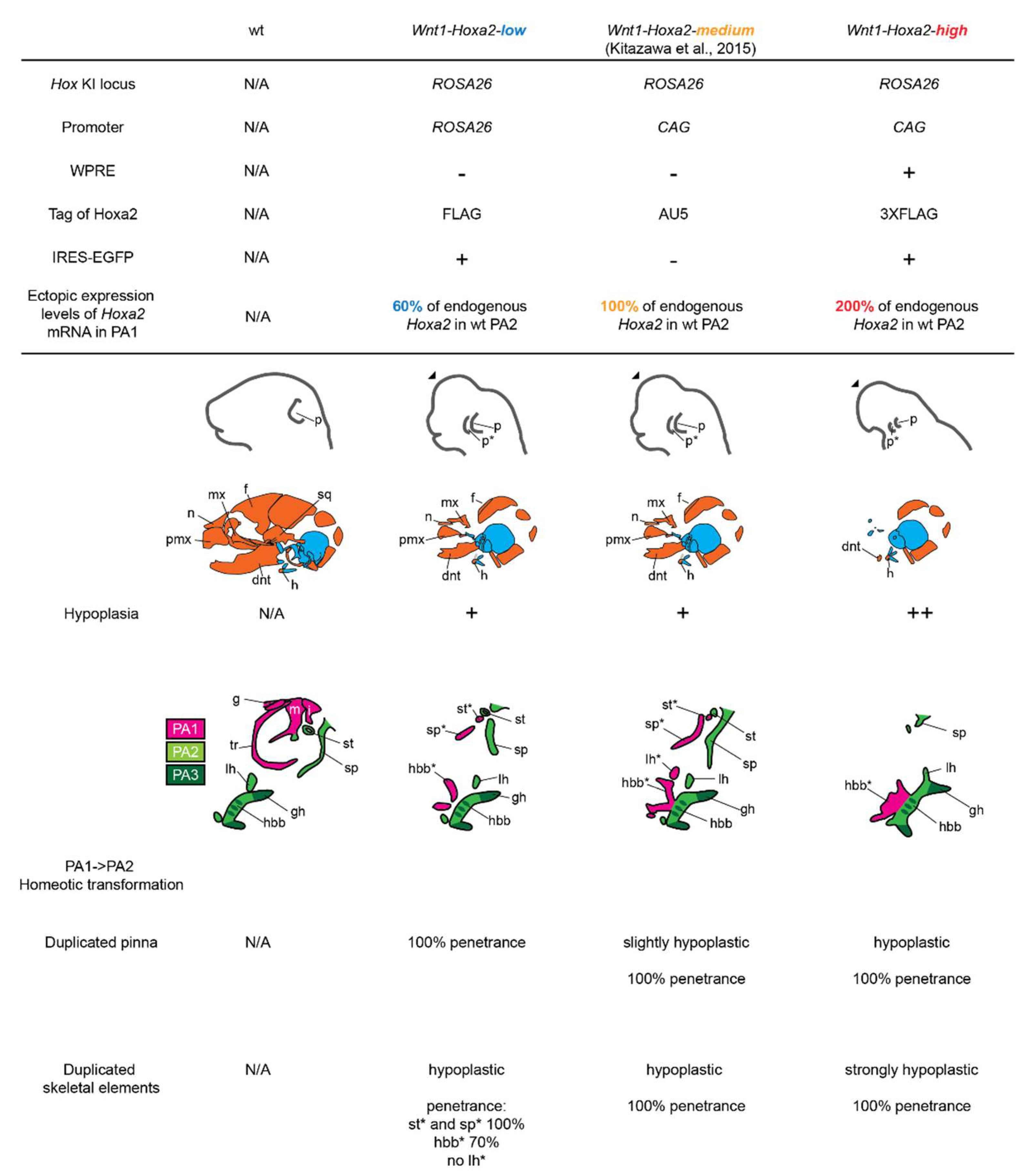
3.1. Analysis of Mouse Phenotypes Induced by Low Levels of Conditional Ectopic Hoxa2 Expression in CNCCs
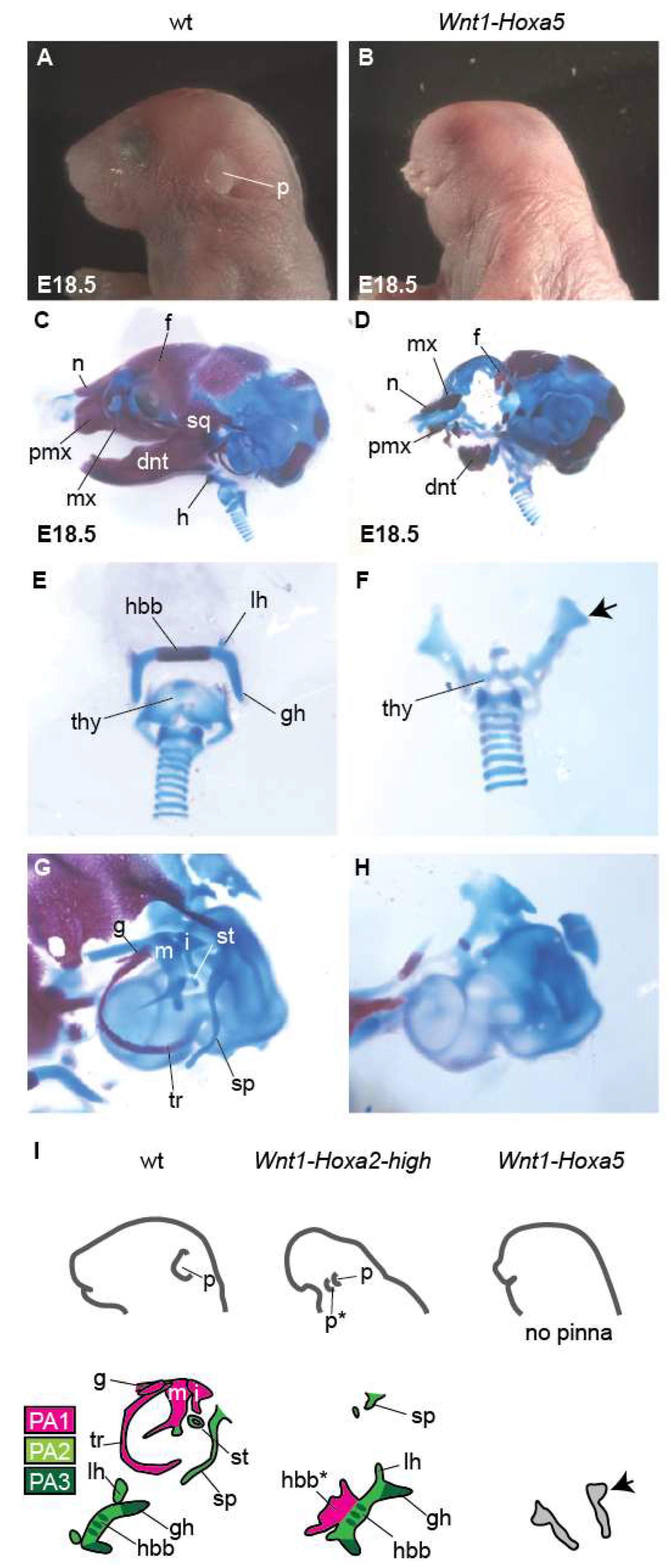
3.2. Analysis of Mouse Phenotypes Induced by High Levels of Conditional Ectopic Hoxa2 Expression in CNCCs
3.3. Analysis of Mouse Phenotypes Induced by Conditional Ectopic Hoxa5 Overexpression in CNCCs
4. Discussion
4.1. Homeotic Repatterning of Hox-Negative CNCCs by Hoxa2 is Dependent on Its Ectopic Expression Levels and CNCC Position within the Craniofacial Complex
4.2. Hoxa5 Overexpression Cannot Homeotically Repattern CNCCs and Generates Distinct Phenotypes Compared With Hoxa2 Overexpression
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Le Douarin, N.M.; Creuzet, S.; Couly, G.; Dupin, E. Neural crest cell plasticity and its limits. Development 2004, 131, 4637–4650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gammill, L.S.; Bronner-Fraser, M. Neural crest specification: Migrating into genomics. Nat. Rev. Neurosci. 2003, 4, 795–805. [Google Scholar] [CrossRef]
- Minoux, M.; Rijli, F.M. Molecular mechanisms of cranial neural crest cell migration and patterning in craniofacial development. Development 2010, 137, 2605–2621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santagati, F.; Rijli, F.M. Cranial neural crest and the building of the vertebrate head. Nat. Rev. Neurosci. 2003, 4, 806–818. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Sipp, D.; Enomoto, H. Tissue interactions in neural crest cell development and disease. Science 2013, 341, 860–863. [Google Scholar] [CrossRef] [PubMed]
- Alexander, T.; Nolte, C.; Krumlauf, R. Hox genes and segmentation of the hindbrain and axial skeleton. Annu. Rev. Cell Dev. Biol. 2009, 25, 431–456. [Google Scholar] [CrossRef] [Green Version]
- Minoux, M.; Antonarakis, G.S.; Kmita, M.; Duboule, D.; Rijli, F.M. Rostral and caudal pharyngeal arches share a common neural crest ground pattern. Development 2009, 136, 637–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trainor, P.; Krumlauf, R. Plasticity in mouse neural crest cells reveals a new patterning role for cranial mesoderm. Nat. Cell Biol. 2000, 2, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Gendron-Maguire, M.; Mallo, M.; Zhang, M.; Gridley, T. Hoxa-2 mutant mice exhibit homeotic transformation of skeletal elements derived from cranial neural crest. Cell 1993, 75, 1317–1331. [Google Scholar] [CrossRef]
- Rijli, F.M.; Mark, M.; Lakkaraju, S.; Dierich, A.; Dolle, P.; Chambon, P. A homeotic transformation is generated in the rostral branchial region of the head by disruption of Hoxa-2, which acts as a selector gene. Cell 1993, 75, 1333–1349. [Google Scholar] [CrossRef]
- Minoux, M.; Kratochwil, C.F.; Ducret, S.; Amin, S.; Kitazawa, T.; Kurihara, H.; Bobola, N.; Vilain, N.; Rijli, F.M. Mouse Hoxa2 mutations provide a model for microtia and auricle duplication. Development 2013, 140, 4386–4397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santagati, F.; Minoux, M.; Ren, S.Y.; Rijli, F.M. Temporal requirement of Hoxa2 in cranial neural crest skeletal morphogenesis. Development 2005, 132, 4927–4936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltzinger, M.; Ori, M.; Pasqualetti, M.; Nardi, I.; Rijli, F.M. Hoxa2 knockdown in Xenopus results in hyoid to mandibular homeosis. Dev. Dyn. 2005, 234, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Hunter, M.P.; Prince, V.E. Zebrafish hox paralogue group 2 genes function redundantly as selector genes to pattern the second pharyngeal arch. Dev. Biol. 2002, 247, 367–389. [Google Scholar] [CrossRef] [Green Version]
- Grammatopoulos, G.A.; Bell, E.; Toole, L.; Lumsden, A.; Tucker, A.S. Homeotic transformation of branchial arch identity after Hoxa2 overexpression. Development 2000, 127, 5355–5365. [Google Scholar] [CrossRef]
- Kitazawa, T.; Fujisawa, K.; Narboux-Neme, N.; Arima, Y.; Kawamura, Y.; Inoue, T.; Wada, Y.; Kohro, T.; Aburatani, H.; Kodama, T.; et al. Distinct effects of Hoxa2 overexpression in cranial neural crest populations reveal that the mammalian hyomandibular-ceratohyal boundary maps within the styloid process. Dev. Biol. 2015, 402, 162–174. [Google Scholar] [CrossRef] [Green Version]
- Pasqualetti, M.; Ori, M.; Nardi, I.; Rijli, F.M. Ectopic Hoxa2 induction after neural crest migration results in homeosis of jaw elements in Xenopus. Development 2000, 127, 5367–5378. [Google Scholar] [CrossRef]
- Creuzet, S.; Couly, G.; Vincent, C.; Le Douarin, N.M. Negative effect of Hox gene expression on the development of the neural crest-derived facial skeleton. Development 2002, 129, 4301–4313. [Google Scholar] [CrossRef]
- Kanzler, B.; Kuschert, S.J.; Liu, Y.H.; Mallo, M. Hoxa-2 restricts the chondrogenic domain and inhibits bone formation during development of the branchial area. Development 1998, 125, 2587–2597. [Google Scholar] [CrossRef]
- Maheshwari, U.; Kraus, D.; Vilain, N.; Holwerda, S.J.B.; Cankovic, V.; Maiorano, N.A.; Kohler, H.; Satoh, D.; Sigrist, M.; Arber, S.; et al. Postmitotic Hoxa5 Expression Specifies Pontine Neuron Positional Identity and Input Connectivity of Cortical Afferent Subsets. Cell Rep. 2020, 31, 107767. [Google Scholar] [CrossRef]
- Moreno-Juan, V.; Filipchuk, A.; Anton-Bolanos, N.; Mezzera, C.; Gezelius, H.; Andres, B.; Rodriguez-Malmierca, L.; Susin, R.; Schaad, O.; Iwasato, T.; et al. Prenatal thalamic waves regulate cortical area size prior to sensory processing. Nat. Commun. 2017, 8, 14172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miguez, A.; Ducret, S.; Di Meglio, T.; Parras, C.; Hmidan, H.; Haton, C.; Sekizar, S.; Mannioui, A.; Vidal, M.; Kerever, A.; et al. Opposing roles for Hoxa2 and Hoxb2 in hindbrain oligodendrocyte patterning. J. Neurosci. 2012, 32, 17172–17185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, A.E.; Vasudevan, H.N.; O'Neill, A.K.; Soriano, P.; Bush, J.O. The widely used Wnt1-Cre transgene causes developmental phenotypes by ectopic activation of Wnt signaling. Dev. Biol. 2013, 379, 229–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitazawa, T.; Takechi, M.; Hirasawa, T.; Adachi, N.; Narboux-Neme, N.; Kume, H.; Maeda, K.; Hirai, T.; Miyagawa-Tomita, S.; Kurihara, Y.; et al. Developmental genetic bases behind the independent origin of the tympanic membrane in mammals and diapsids. Nat. Commun. 2015, 6, 6853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, Y.; Jiang, X.; Ito, Y.; Bringas, P., Jr.; Han, J.; Rowitch, D.H.; Soriano, P.; McMahon, A.P.; Sucov, H.M. Fate of the mammalian cranial neural crest during tooth and mandibular morphogenesis. Development 2000, 127, 1671–1679. [Google Scholar] [CrossRef] [PubMed]
- Higashimoto, T.; Urbinati, F.; Perumbeti, A.; Jiang, G.; Zarzuela, A.; Chang, L.J.; Kohn, D.B.; Malik, P. The woodchuck hepatitis virus post-transcriptional regulatory element reduces readthrough transcription from retroviral vectors. Gene Ther. 2007, 14, 1298–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zufferey, R.; Donello, J.E.; Trono, D.; Hope, T.J. Woodchuck hepatitis virus posttranscriptional regulatory element enhances expression of transgenes delivered by retroviral vectors. J. Virol. 1999, 73, 2886–2892. [Google Scholar] [CrossRef] [Green Version]
- McBratney-Owen, B.; Iseki, S.; Bamforth, S.D.; Olsen, B.R.; Morriss-Kay, G.M. Development and tissue origins of the mammalian cranial base. Dev. Biol. 2008, 322, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Tavella, S.; Bobola, N. Expressing Hoxa2 across the entire endochondral skeleton alters the shape of the skeletal template in a spatially restricted fashion. Differentiation 2010, 79, 194–202. [Google Scholar] [CrossRef]
- Couly, G.; Grapin-Botton, A.; Coltey, P.; Ruhin, B.; Le Douarin, N.M. Determination of the identity of the derivatives of the cephalic neural crest: Incompatibility between Hox gene expression and lower jaw development. Development 1998, 125, 3445–3459. [Google Scholar] [CrossRef]
- Massip, L.; Ectors, F.; Deprez, P.; Maleki, M.; Behets, C.; Lengele, B.; Delahaut, P.; Picard, J.; Rezsohazy, R. Expression of Hoxa2 in cells entering chondrogenesis impairs overall cartilage development. Differentiation 2007, 75, 256–267. [Google Scholar] [CrossRef] [Green Version]
- Compagnucci, C.; Debiais-Thibaud, M.; Coolen, M.; Fish, J.; Griffin, J.N.; Bertocchini, F.; Minoux, M.; Rijli, F.M.; Borday-Birraux, V.; Casane, D.; et al. Pattern and polarity in the development and evolution of the gnathostome jaw: Both conservation and heterotopy in the branchial arches of the shark, Scyliorhinus canicula. Dev. Biol. 2013, 377, 428–448. [Google Scholar] [CrossRef] [PubMed]
- Couly, G.; Creuzet, S.; Bennaceur, S.; Vincent, C.; Le Douarin, N.M. Interactions between Hox-negative cephalic neural crest cells and the foregut endoderm in patterning the facial skeleton in the vertebrate head. Development 2002, 129, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Minoux, M.; Holwerda, S.; Vitobello, A.; Kitazawa, T.; Kohler, H.; Stadler, M.B.; Rijli, F.M. Gene bivalency at Polycomb domains regulates cranial neural crest positional identity. Science 2017, 355, eaal2913. [Google Scholar] [CrossRef] [PubMed]
- Rux, D.R.; Wellik, D.M. Hox genes in the adult skeleton: Novel functions beyond embryonic development. Dev. Dyn. 2017, 246, 310–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitazawa, T.; Minoux, M.; Ducret, S.; Rijli, F.M. Different Ectopic Hoxa2 Expression Levels in Mouse Cranial Neural Crest Cells Result in Distinct Craniofacial Anomalies and Homeotic Phenotypes. J. Dev. Biol. 2022, 10, 9. https://0-doi-org.brum.beds.ac.uk/10.3390/jdb10010009
Kitazawa T, Minoux M, Ducret S, Rijli FM. Different Ectopic Hoxa2 Expression Levels in Mouse Cranial Neural Crest Cells Result in Distinct Craniofacial Anomalies and Homeotic Phenotypes. Journal of Developmental Biology. 2022; 10(1):9. https://0-doi-org.brum.beds.ac.uk/10.3390/jdb10010009
Chicago/Turabian StyleKitazawa, Taro, Maryline Minoux, Sebastien Ducret, and Filippo M. Rijli. 2022. "Different Ectopic Hoxa2 Expression Levels in Mouse Cranial Neural Crest Cells Result in Distinct Craniofacial Anomalies and Homeotic Phenotypes" Journal of Developmental Biology 10, no. 1: 9. https://0-doi-org.brum.beds.ac.uk/10.3390/jdb10010009