The Smk1 MAPK and Its Activator, Ssp2, Are Required for Late Prospore Membrane Development in Sporulating Saccharomyces cerevisiae

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Plasmids
2.2. Immunoblotting
2.3. Microscopy
3. Results
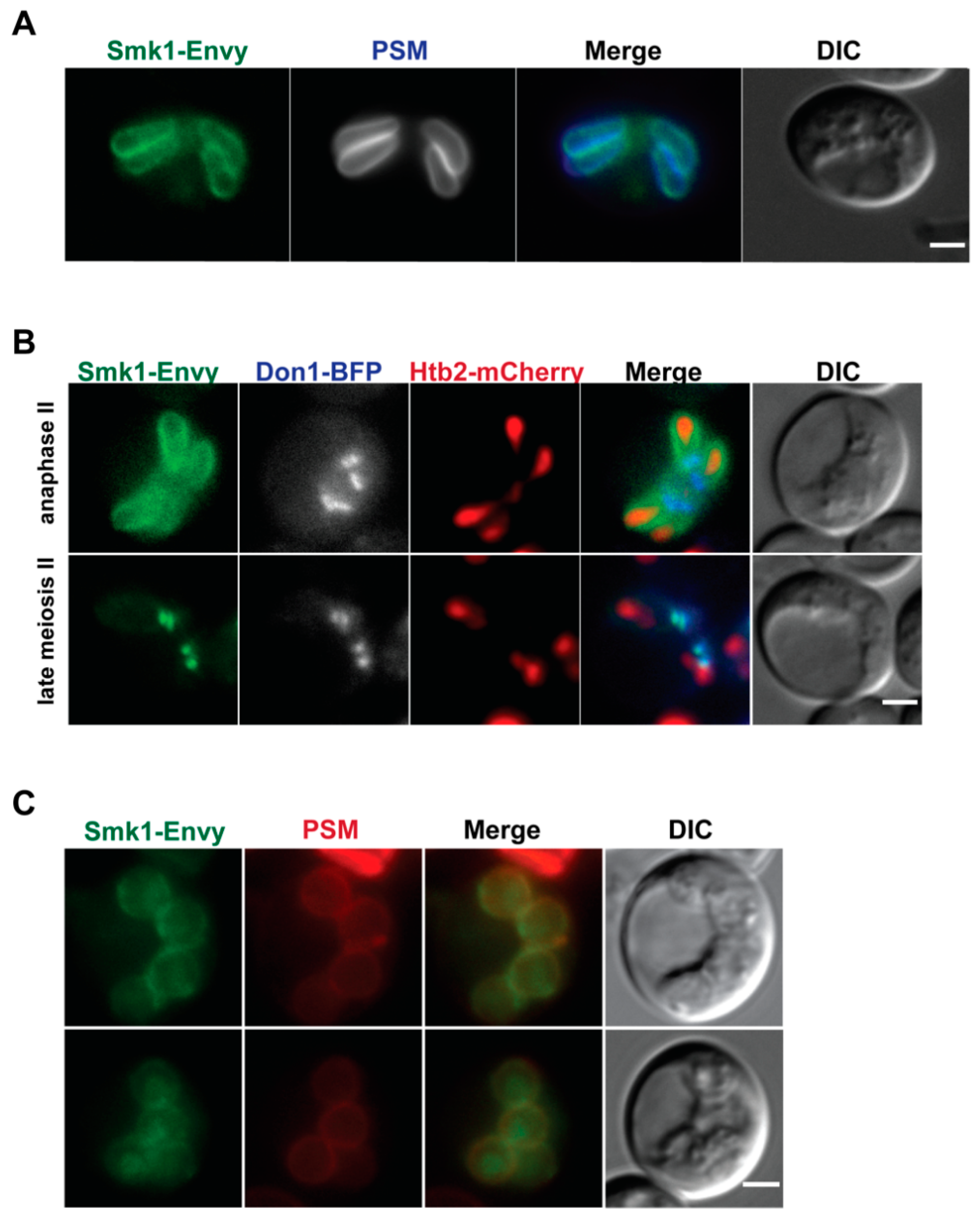
3.1. Smk1 Localizes to the Leading Edge of Elongated Prospore Membranes
3.2. Smk1 Localization at the Leading Edge of the Prospore Membrane Requires SSP2
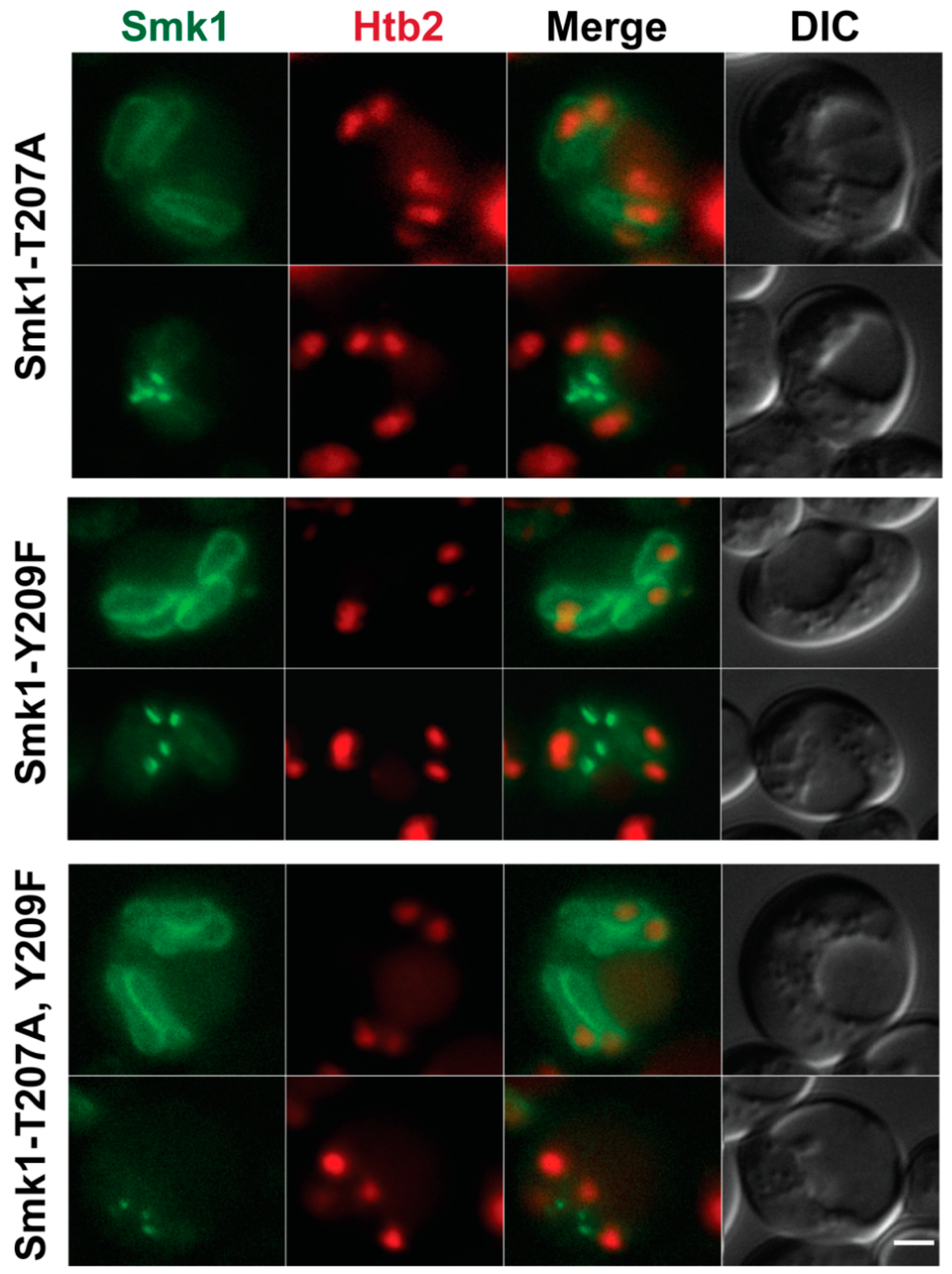
3.3. Smk1 Localization Is Independent of Activation State
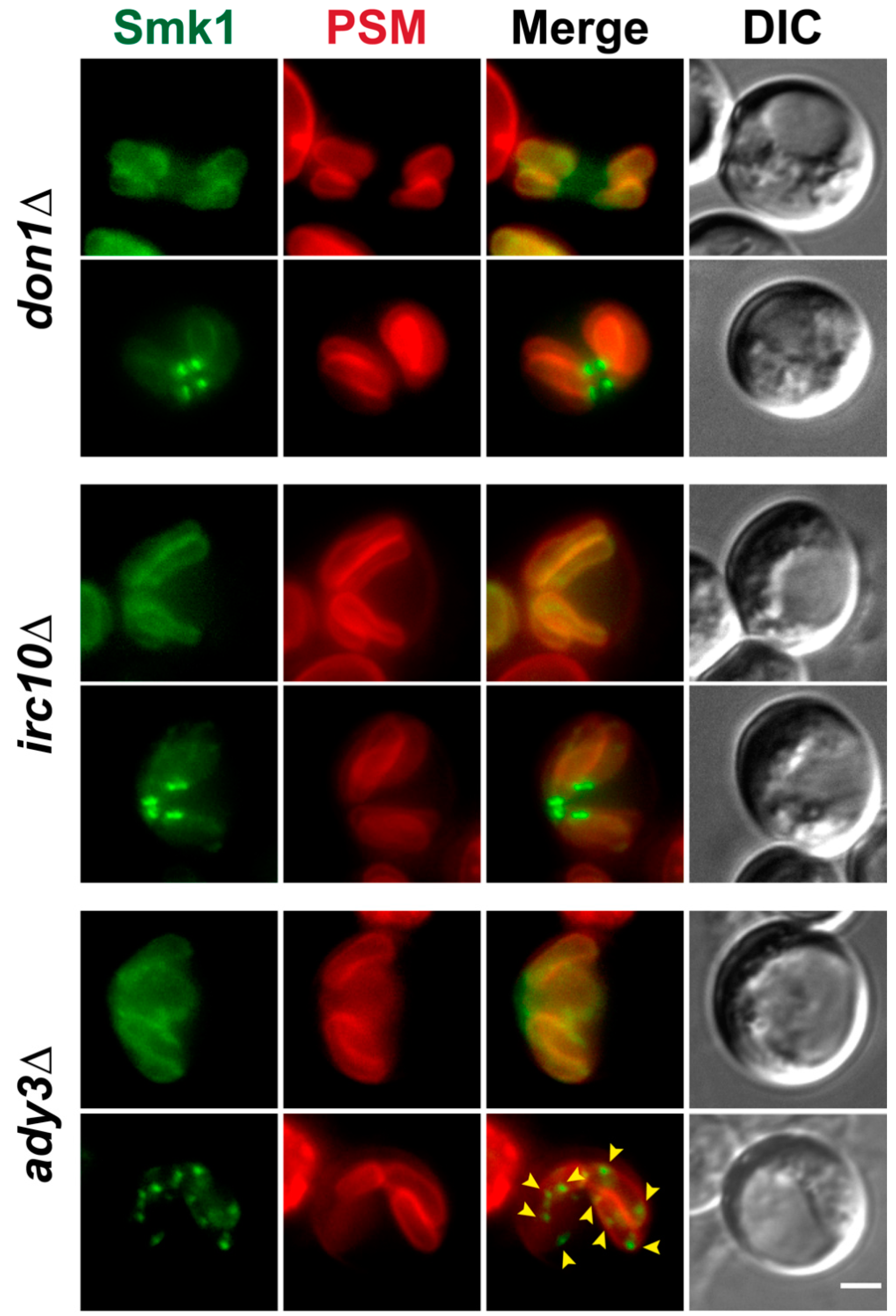
3.4. Localization of Smk1 Requires the LEP Member ADY3, But Not IRC10 or DON1
3.5. Smk1 Phosphorylation Does Not Require the LEP Complex
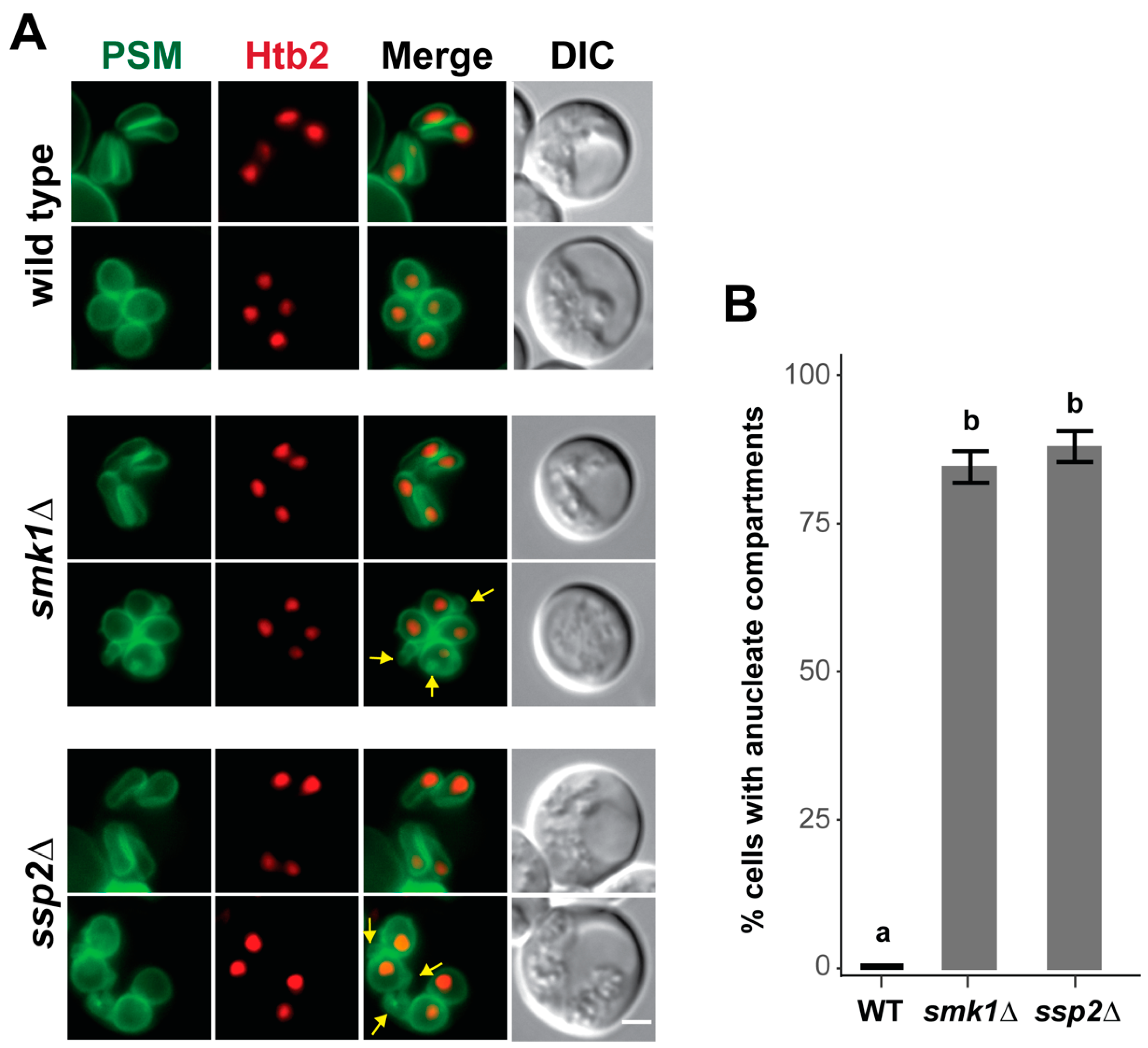
3.6. SMK1 and SSP2 Are Required for Late Prospore Membrane Development
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Neiman, A.M. Sporulation in the budding yeast Saccharomyces cerevisiae. Genetics 2011, 189, 737–765. [Google Scholar] [CrossRef] [Green Version]
- Neiman, A.M. Prospore Membrane Formation Defines a Developmentally Regulated Branch of the Secretory Pathway in Yeast. J. Cell Biol. 1998, 140, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Mathieson, E.M.; Suda, Y.; Nickas, M.; Snydsman, B.; Davis, T.N.; Muller, E.G.D.; Neiman, A.M.; Brennwald, P.J. Vesicle Docking to the Spindle Pole Body Is Necessary to Recruit the Exocyst During Membrane Formation in Saccharomyces cerevisiae. Mol. Biol. Cell 2010, 21, 3693–3707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suda, Y.; Tachikawa, H.; Inoue, I.; Kurita, T.; Saito, C.; Kurokawa, K.; Nakano, A.; Irie, K. Activation of Rab GTPase Sec4 by its GEF Sec2 is required for prospore membrane formation during sporulation in yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2018, 18, fox095. [Google Scholar] [CrossRef] [PubMed]
- Diamond, A.E.; Park, J.-S.; Inoue, I.; Tachikawa, H.; Neiman, A.M. The Anaphase Promoting Complex Targeting Subunit Ama1 Links Meiotic Exit to Cytokinesis during Sporulation in Saccharomyces cerevisiae. Mol. Biol. Cell 2009, 20, 134–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maier, P.; Rathfelder, N.; Finkbeiner, M.G.; Taxis, C.; Mazza, M.; Panse, S.L.; Haguenauer-Tsapis, R.; Knop, M. Cytokinesis in yeast meiosis depends on the regulated removal of Ssp1p from the prospore membrane. EMBO J. 2007, 26, 1843–1852. [Google Scholar] [CrossRef]
- Knop, M.; Strasser, K. Role of the spindle pole body of yeast in mediating assembly of the prospore membrane during meiosis. EMBO J. 2000, 19, 3657–3667. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Borchart, A.C.; Strasser, K.; Finkbeiner, M.G.; Shevchenko, A.; Shevchenko, A.; Knop, M. Prospore membrane formation linked to the leading edge protein (LEP) coat assembly. EMBO J. 2001, 20, 6846–6857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nickas, M.E.; Neiman, A.M. Ady3p Links Spindle Pole Body Function to Spore Wall Synthesis in Saccharomyces cerevisiae. Genetics 2002, 160, 1439–1450. [Google Scholar] [PubMed]
- Lam, C.; Santore, E.; Lavoie, E.; Needleman, L.; Fiacco, N.; Kim, C.; Neiman, A.M. A visual screen of protein localization during sporulation identifies new components of prospore membrane-associated complexes in budding yeast. Eukaryot. Cell 2014, 13, 383–391. [Google Scholar] [CrossRef] [Green Version]
- Maier, P.; Rathfelder, N.; Maeder, C.I.; Colombelli, J.; Stelzer, E.H.K.; Knop, M. The SpoMBe pathway drives membrane bending necessary for cytokinesis and spore formation in yeast meiosis. EMBO J. 2008, 27, 2363–2374. [Google Scholar] [CrossRef] [Green Version]
- Paulissen, S.M.; Slubowski, C.J.; Roesner, J.M.; Huang, L.S. Timely closure of the prospore membrane requires SPS1 and SPO77 in Saccharomyces cerevisiae. Genetics 2016, 203, 1203–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krisak, L.; Strich, R.; Winters, R.S.; Hall, J.P.; Mallory, M.J.; Kreitzer, D.; Tuan, R.S.; Winter, E. SMK1, a developmentally regulated MAP kinase, is required for spore wall assembly in Saccharomyces cerevisiae. Genes Dev. 1994, 8, 2151–2161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, M.; Briza, P.; Pierce, M.; Winter, E. Distinct Steps in Yeast Spore Morphogenesis Require Distinct SMK1 MAP Kinase Thresholds. Genetics 1999, 15, 1327–1340. [Google Scholar]
- Huang, L.S.; Doherty, H.K.; Herskowitz, I. The Smk1p MAP kinase negatively regulates Gsc2p, a 1,3-glucan synthase, during spore wall morphogenesis in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2005, 102, 12431–12436. [Google Scholar] [CrossRef] [Green Version]
- Tio, C.W.; Omerza, G.; Sunder, S.; Winter, E. Autophosphorylation of the Smk1 MAPK is spatially and temporally regulated by Ssp2 during meiotic development in yeast. Mol. Biol. Cell 2015, 26, 3546–3555. [Google Scholar] [CrossRef]
- Tio, C.W.; Omerza, G.; Phillips, T.; Lou, H.J.; Turk, B.E.; Winter, E. Ssp2 Binding Activates the Smk1 Mitogen-Activated Protein Kinase. Mol. Cell Biol. 2017, 37, e00607-16. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, P.; Florczyk, M.; McDonough, K.; Nag, D. SSP2, a sporulation-specific gene necessary for outer spore wall assembly in the yeast Saccharomyces cerevisiae. Mol. Genet. Genom. 2002, 267, 348–358. [Google Scholar] [CrossRef]
- Coluccio, A.; Bogengruber, E.; Conrad, M.N.; Dresser, M.E.; Briza, P.; Neiman, A.M. Morphogenetic Pathway of Spore Wall Assembly in Saccharomyces cerevisiae. Eukaryot. Cell 2004, 3, 1464–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Agarwal, S.; Roeder, G.S. SSP2 and OSW1, two sporulation-specific genes involved in spore morphogenesis in Saccharomyces cerevisiae. Genetics 2007, 175, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Kane, S.M.; Roth, R. Carbohydrate Metabolism during Ascospore Development in Yeast. J. Bacteriol. 1974, 118, 8–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, M.D.; Fink, G.F. Methods in Yeast Genetics; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1990. [Google Scholar]
- Longtine, M.S.; Mckenzie III, A.; Demarini, D.J.; Shah, N.G.; Wach, A.; Brachat, A.; Philippsen, P.; Pringle, J.R. Additional modules for versatile and economical PCR-based gene deletion and modification in Saccharomyces cerevisiae. Yeast 1998, 14, 953–961. [Google Scholar] [CrossRef]
- Lee, S.; Lim, W.A.; Thorn, K.S. Improved Blue, Green, and Red Fluorescent Protein Tagging Vectors for S. cerevisiae. PLoS ONE 2013, 8, e67902. [Google Scholar] [CrossRef] [PubMed]
- Slubowski, C.J.; Funk, A.D.; Roesner, J.M.; Paulissen, S.M.; Huang, L.S. Plasmids for C-terminal tagging in Saccharomyces cerevisiae that contain improved GFP proteins, Envy and Ivy. Yeast 2015, 32, 379–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.S.; Numajiri, Y.; Okumura, Y.; Hidaka, J.; Tanaka, T.; Inoue, I.; Suda, Y.; Takahashi, T.; Nakanishi, H.; Gao, X.D.; et al. Dynamic localization of a yeast development-specific PP1 complex during prospore membrane formation is dependent on multiple localization signals and complex formation. Mol. Biol. Cell 2017, 28, 3881–3895. [Google Scholar] [CrossRef] [Green Version]
- Paulissen, S.M.; Hunt, C.A.; Seitz, B.C.; Slubowski, C.J.; Yu, Y.; Mucelli, X.; Truong, D.; Wallis, Z.; Nguyen, H.T.; Newman-Toledo, S.; et al. A noncanonical hippo pathway regulates spindle disassembly and cytokinesis during meiosis in Saccharomyces cerevisiae. Genetics 2020, 216, 447–462. [Google Scholar] [CrossRef]
- Lin, C.P.C.; Kim, C.; Smith, S.O.; Neiman, A.M. A Highly Redundant Gene Network Controls Assembly of the Outer Spore Wall in S. cerevisiae. PLoS Genet. 2013, 9, e1003700. [Google Scholar] [CrossRef] [Green Version]
- Sikorski, R.S.; Hieter, P. A System of Shuttle Vectors and Yeast Host Strains Designed for Efficient Manipulation of DNA in Saccharomyces cerevisiae. Genetics 1989, 122, 19–27. [Google Scholar] [CrossRef]
- Philips, J.; Herskowitz, I. Identification of Kel1p, a Kelch Domain-containing Protein Involved in Cell Fusion and Morphology in Saccharomyces cerevisiae. J. Cell Biol. 1998, 143, 375–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, H.; de los Santos, P.; Neiman, A.M. Positive and Negative Regulation of a SNARE Protein by Control of Intracellular Localization. Mol. Biol. Cell 2004, 15, 1802–1815. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, K.; Bell, R.B.; Yunus, J.; Amon, A.; Berchowitz, L.E. Phosphorylation-Mediated Clearance of Amyloid-like Assemblies in Meiosis. Dev. Cell 2018, 45, 392–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ to 3′) |
|---|---|
| OLH1756 | AATCGAGCTCAGTAATAAATACTGTGTTGTTTG |
| OLH1757 | GAAAATGTTGGACCATG |
| OLH1773 | GTACTCCTGATAAAGATATTCTG |
| OLH1774 | CAGCACCGTCACCTAAAGACGAGGAGGACAAATC |
| OLH1758 | TATAAAAAAATCCCATGGTC |
| OLH1759 | ATTGCTCGAGTTATTTGTACAATTCGTCCATTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Durant, M.; Roesner, J.M.; Mucelli, X.; Slubowski, C.J.; Klee, E.; Seitz, B.C.; Wallis, Z.; Huang, L.S. The Smk1 MAPK and Its Activator, Ssp2, Are Required for Late Prospore Membrane Development in Sporulating Saccharomyces cerevisiae. J. Fungi 2021, 7, 53. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010053
Durant M, Roesner JM, Mucelli X, Slubowski CJ, Klee E, Seitz BC, Wallis Z, Huang LS. The Smk1 MAPK and Its Activator, Ssp2, Are Required for Late Prospore Membrane Development in Sporulating Saccharomyces cerevisiae. Journal of Fungi. 2021; 7(1):53. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010053
Chicago/Turabian StyleDurant, Matthew, Joseph M. Roesner, Xheni Mucelli, Christian J. Slubowski, Erin Klee, Brian C. Seitz, Zoey Wallis, and Linda S. Huang. 2021. "The Smk1 MAPK and Its Activator, Ssp2, Are Required for Late Prospore Membrane Development in Sporulating Saccharomyces cerevisiae" Journal of Fungi 7, no. 1: 53. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010053