High-Throughput Volatilome Fingerprint Using PTR–ToF–MS Shows Species-Specific Patterns in Mortierella and Closely Related Genera

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of Mortierella s. l. Strains
2.2. Identification of Obtained Mortierella Isolates
2.3. Cultivation of Mortierella Isolates for Volatilome Analysis
2.4. Analysis of PTR–ToF–Mass Spectrometer
2.5. Data Analysis
3. Results
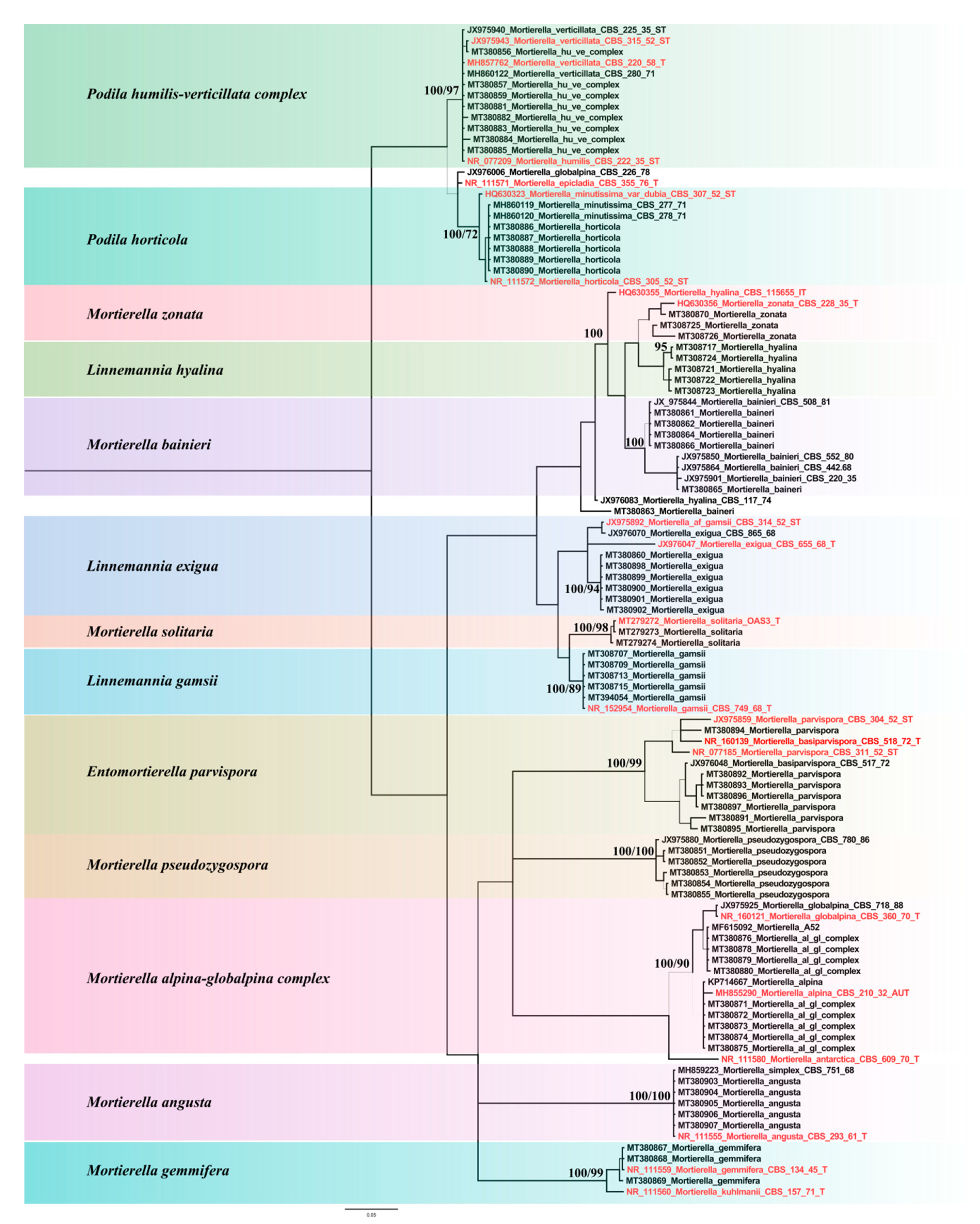
3.1. Origin and Identification of Isolated Mortierella Strains
3.2. The Vast Majority of Detected VOCs was Produced by All Mortierella Species
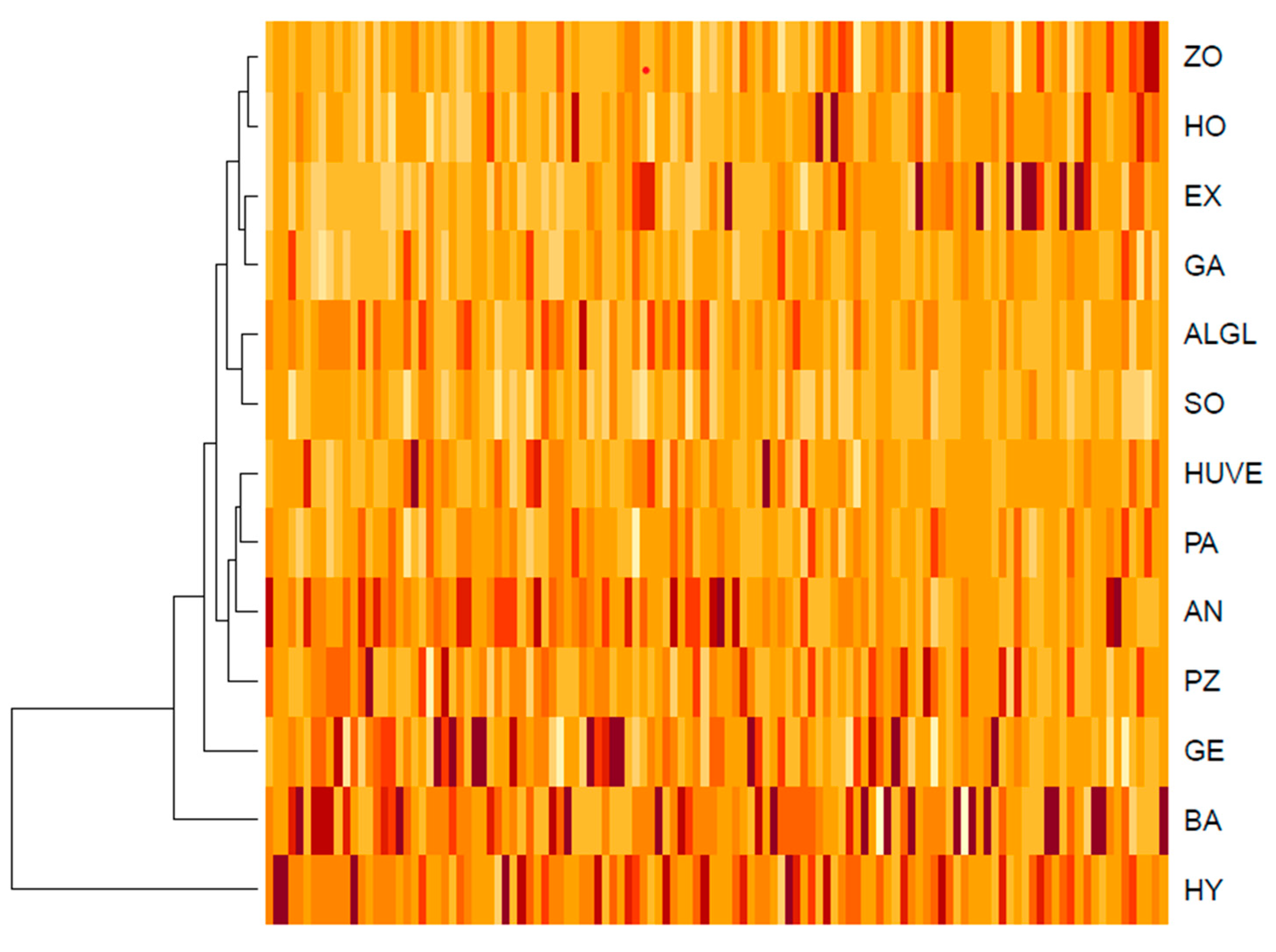
3.3. Mortierella Species Differ in Their Volatilomes
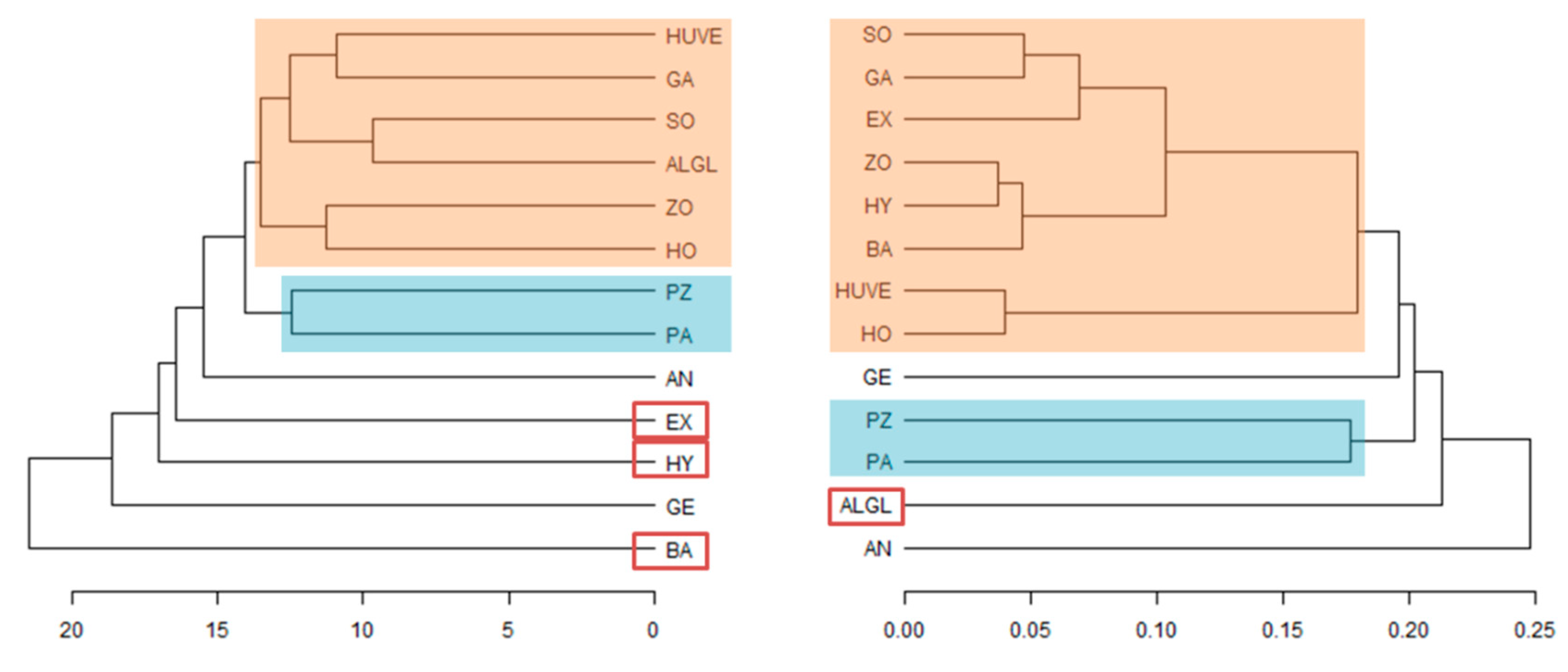
3.4. Mortierella Phylogeny Corresponds to Volatilome
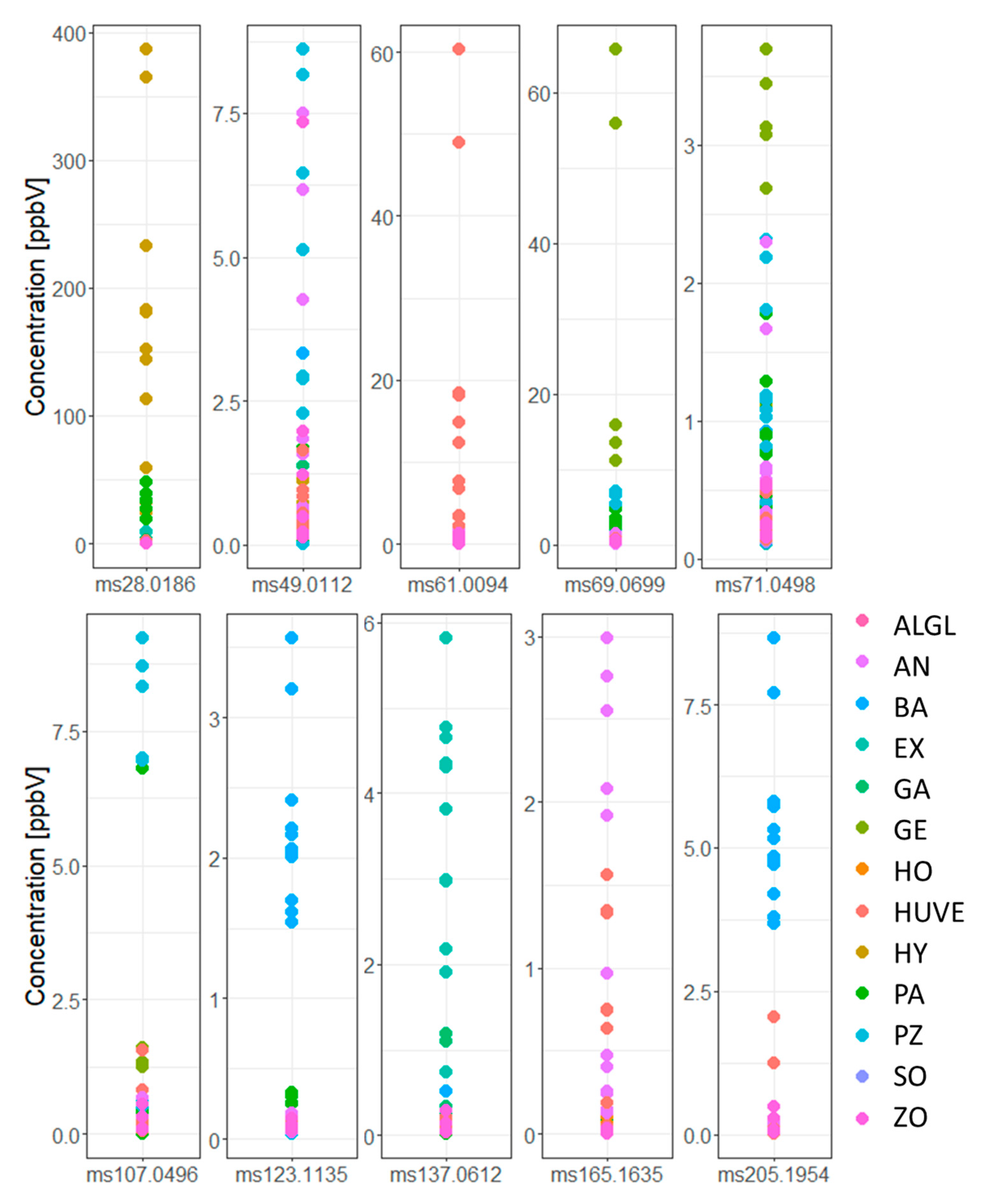
3.5. Discriminating Mortierella s. l. Species by Which VOCs?
3.6. Location Effects on Mortierella Volatilomes
3.7. Mortierella and Their Species-Specifically Enriched VOCs
4. Discussion
4.1. PTR–ToF–MS as a High-Throughput Method for VOC Detection
4.2. All Mortierella Species Produce a Variety of VOCs, But VOC Concentrations Are Species-Specific
4.3. Potential Function of VOCs Produced by Mortierella Species
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farh, M.E.A.; Jeon, J. Roles of fungal volatiles from perspective of distinct lifestyles in filamentous fungi. Plant Pathol. J. 2020, 36, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Wenke, K.; Kai, M.; Piechulla, B. Belowground volatiles facilitate interactions between plant roots and soil organisms. Planta 2010, 231, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Schulz-Bohm, K.; Martín-Sánchez, L.; Garbeva, P. Microbial volatiles: Small molecules with an important role in intra- and inter-kingdom interactions. Front. Microbiol. 2017, 8, 2484. [Google Scholar] [CrossRef] [PubMed]
- Hung, R.; Lee, S.; Bennett, J.W. Fungal volatile organic compounds and their role in ecosystems. Appl. Microbiol. Biotechnol. 2015, 99, 3395–3405. [Google Scholar] [CrossRef] [PubMed]
- Effmert, U.; Kalderás, J.; Warnke, R.; Piechulla, B. Volatile Mediated Interactions Between Bacteria and Fungi in the Soil. J. Chem. Ecol. 2012, 38, 665–703. [Google Scholar] [CrossRef]
- Linnemann, G. Pflanzenforschung: Die Mucorineen-Gattung Mortierella Coemans; Heft 23; Kolkwitz, R., Ed.; Gustav Fischer: Jena, Germany, 1941; ISBN B00CFCGREO. [Google Scholar]
- Degawa, Y.; Gams, W. A new species of Mortierella, and an associated sporangiiferous mycoparasite in a new genus, Nothadelphia. Stud. Mycol. 2004, 50, 567–572. [Google Scholar]
- Wagner, L.; Stielow, B.; Hoffmann, K.; Petkovits, T.; Papp, T.; Vágvölgyi, C.; de Hoog, G.S.; Verkley, G.; Voigt, K. A comprehensive molecular phylogeny of the Mortierellales (Mortierellomycotina) based on nuclear ribosomal DNA. Pers. Mol. Phylogeny Evol. Fungi 2013, 30, 77–93. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global diversity and geography of soil fungi. Science 2014, 346. [Google Scholar] [CrossRef] [Green Version]
- Vandepol, N.; Liber, J.; Desirò, A.; Na, H.; Kennedy, M.; Barry, K.; Grigoriev, I.V.; Miller, A.N.; O’Donnell, K.; Stajicjh, J.E.; et al. Resolving the Mortierellaceae phylogeny through synthesis of multi-gene phylogenetics and phylogenomics. Fungal Divers. 2020, 104, 267–289. [Google Scholar] [CrossRef]
- DiLegge, M.J.; Manter, D.K.; Vivanco, J.M. A novel approach to determine generalist nematophagous microbes reveals Mortierella globalpina as a new biocontrol agent against Meloidogyne spp. nematodes. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Wani, Z.A.; Kumar, A.; Sultan, P.; Bindu, K.; Riyaz-Ul-Hassan, S.; Ashraf, N. Mortierella alpina CS10E4, an oleaginous fungal endophyte of Crocus sativus L. enhances apocarotenoid biosynthesis and stress tolerance in the host plant. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ozimek, E.; Jaroszuk-ściseł, J.; Bohacz, J.; Korniłłowicz-Kowalska, T.; Tyśkiewicz, R.; Słomka, A.; Nowak, A.; Hanaka, A. Synthesis of indoleacetic acid, gibberellic acid and acc-deaminase by Mortierella strains promote winter wheat seedlings growth under different conditions. Int. J. Mol. Sci. 2018, 19, 3218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topalović, O.; Hussain, M.; Heuer, H. Plants and Associated Soil Microbiota Cooperatively Suppress Plant-Parasitic Nematodes. Front. Microbiol. 2020, 11, 313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinmen, Y.; Shimizu, S.; Akimoto, K.; Kawashima, H.; Yamada, H. Production of arachidonic acid by Mortierella fungi—Selection of a potent producer and optimization of culture conditions for large-scale production. Appl. Microbiol. Biotechnol. 1989, 31, 11–16. [Google Scholar] [CrossRef]
- Baldeweg, F.; Warncke, P.; Fischer, D.; Gressler, M. Fungal Biosurfactants from Mortierella alpina. Org. Lett. 2019, 21, 1444–1448. [Google Scholar] [CrossRef] [PubMed]
- Yamada, H.; Shimizu, S.; Shinmen, Y. Production of arachidonic acid by Mortierella elongata 1s-5. Agric. Biol. Chem. 1987, 51, 785–790. [Google Scholar] [CrossRef]
- Xin, L.I.U.; Xiaochen, M.; He, H. others Analysisi for the volatile secondary metabolites of Mortierella alpina. Agric. Sci. Technol. 2009, 10, 12–14. [Google Scholar]
- Kanchiswamy, C.N.; Malnoy, M.; Maffei, M.E. Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 2015, 6, 151. [Google Scholar] [CrossRef] [Green Version]
- Lindinger, W.; Hansel, A.; Jordan, A. On-line monitoring of volatile organic compounds at pptv levels by means of Proton-Transfer-Reaction Mass Spectrometry (PTR-MS) Medical applications, food control and environmental research. Int. J. Mass Spectrom. Ion Process. 1998, 173, 191–241. [Google Scholar] [CrossRef]
- Mancuso, S.; Taiti, C.; Bazihizina, N.; Costa, C.; Menesatti, P.; Giagnoni, L.; Arenella, M.; Nannipieri, P.; Renella, G. Soil volatile analysis by proton transfer reaction-time of flight mass spectrometry (PTR-TOF-MS). Appl. Soil Ecol. 2015, 86, 182–191. [Google Scholar] [CrossRef]
- Khomenko, I.; Stefanini, I.; Cappellin, L.; Cappelletti, V.; Franceschi, P.; Cavalieri, D.; Märk, T.D.; Biasioli, F. Non-invasive real time monitoring of yeast volatilome by PTR-ToF-MS. Metabolomics 2017, 13, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Jud, W.; Ghirardo, A.; Antritter, F.; Benz, J.P.; Schnitzler, J.P.; Rosenkranz, M. Sniffing fungi-phenotyping of volatile chemical diversity in Trichoderma species. New Phytol. 2020, 227, 244–259. [Google Scholar] [CrossRef] [PubMed]
- Brilli, F.; Gioli, B.; Zona, D.; Pallozzi, E.; Zenone, T.; Fratini, G.; Calfapietra, C.; Loreto, F.; Janssens, I.A.; Ceulemans, R. Simultaneous leaf- and ecosystem-level fluxes of volatile organic compounds from a poplar-based SRC plantation. Agric. For. Meteorol. 2014, 187, 22–35. [Google Scholar] [CrossRef] [Green Version]
- Graus, M.; Müller, M.; Hansel, A. High resolution PTR-TOF: Quantification and Formula Confirmation of VOC in Real Time. J. Am. Soc. Mass Spectrom. 2010, 21, 1037–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takashima, Y.; Degawa, Y.; Narisawa, K. Mortierella oedorhiza, a new species forming a dichotomously branched rhizoid at the sporangiophore base. Mycoscience 2019, 60, 361–365. [Google Scholar] [CrossRef]
- Ganthaler, A.; Stöggl, W.; Kranner, I.; Mayr, S. Foliar phenolic compounds in Norway spruce with varying susceptibility to Chrysomyxa rhododendri: Analyses of seasonal and infection-induced accumulation patterns. Front. Plant Sci. 2017, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Rainer, G.; Kuhnert, R.; Unterholzer, M.; Dresch, P.; Gruber, A.; Peintner, U. Host-specialist dominated ectomycorrhizal communities of Pinus cembra are not affected by temperature manipulation. J. Fungi 2015, 1, 55–75. [Google Scholar] [CrossRef]
- Kuhnert, R.; Oberkofler, I.; Peintner, U. Fungal Growth and Biomass Development is Boosted by Plants in Snow-Covered Soil. Microb. Ecol. 2012, 64, 79–90. [Google Scholar] [CrossRef]
- Dresch, P.; Falbesoner, J.; Ennemoser, C.; Hittorf, M.; Kuhnert, R.; Peintner, U. Emerging from the ice-fungal communities are diverse and dynamic in earliest soil developmental stages of a receding glacier. Environ. Microbiol. 2019, 21, 1864–1880. [Google Scholar] [CrossRef] [Green Version]
- Larcher, W.; Cernusca, A.; Schmidt, L.; Grabherr, G.; Nötzel, E.; Smeets, N. Mt. Patscherkofel, Austria. In Structure and function of tundra ecosystems; Rosswall, R., Heal, O.W., Eds.; Swedish Natural Science Research Council: Stockholm, Sweden, 1975; pp. 125–139. ISBN 91-546-0188-6. [Google Scholar]
- Walch, G.; Knapp, M.; Rainer, G.; Peintner, U. Colony-PCR is a rapid method for DNA amplification of hyphomycetes. J. Fungi 2016, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.A.; Gelfand, D.H.; Sninsky, J.J. PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree v1. 4: Tree Figure Drawing Tool. 2009. Available online: http://tree.bio.ed.ac.uk/website (accessed on 25 April 2014).
- Barber, S.; Blake, R.S.; White, I.R.; Monks, P.S.; Reich, F.; Mullock, S.; Ellis, A.M. Increased sensitivity in proton transfer reaction mass spectrometry by incorporation of a radio frequency ion funnel. Anal. Chem. 2012, 84, 5387–5391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cappellin, L.; Soukoulis, C.; Aprea, E.; Granitto, P.; Dallabetta, N.; Costa, F.; Viola, R.; Märk, T.D.; Gasperi, F.; Biasioli, F. PTR-ToF-MS and data mining methods: A new tool for fruit metabolomics. Metabolomics 2012, 8, 761–770. [Google Scholar] [CrossRef]
- Farneti, B.; Khomenko, I.; Grisenti, M.; Ajelli, M.; Betta, E.; Algarra, A.A.; Cappellin, L.; Aprea, E.; Gasperi, F.; Biasioli, F.; et al. Exploring blueberry aroma complexity by chromatographic and direct-injection spectrometric techniques. Front. Plant Sci. 2017, 8, 617. [Google Scholar] [CrossRef] [PubMed]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Pohlert, T. The Pairwise Multiple Comparison of Mean Ranks Package (PMCMR). 2014. Available online: https://CRAN.R-project.org/package=PMCMR (accessed on 21 October 2020).
- Zeileis, A.; Fisher, J.C.; Hornik, K.; Ihaka, R.; McWhite, C.D.; Murrell, P.; Stauffer, R.; Wilke, C.O. colorspace: A toolbox for manipulating and assessing colors and palettes. arXiv 2019, arXiv:1903.06490. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H.; et al. Community ecology package. R Packag. Version 2013, 1–2. [Google Scholar]
- Galili, T. dendextend: An R package for visualizing, adjusting and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, R.; Etalo, D.W.; de Jager, V.; Gerards, S.; Zweers, H.; de Boer, W.; Garbeva, P. Microbial small talk: Volatiles in fungal-bacterial interactions. Front. Microbiol. 2016, 6, 1495. [Google Scholar] [CrossRef] [Green Version]
- Zhang, E.; Cao, Y.; Xia, Y. Ethanol dehydrogenase I contributes to growth and sporulation under low oxygen condition via detoxification of acetaldehyde in Metarhizium acridum. Front. Microbiol. 2018, 9, 1932. [Google Scholar] [CrossRef]
- Dimitrova, L.; Zaharieva, M.M.; Popova, M.; Kostadinova, N.; Tsvetkova, I.; Bankova, V.; Najdenski, H. Antimicrobial and antioxidant potential of different solvent extracts of the medicinal plant Geum urbanum L. Chem. Cent. J. 2017, 11, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, S.; Basukriadi, A.; Pawiroharsono, S.; Sjamsuridzal, W. Insecticidal activity of ethyl acetate extracts from culture filtrates of mangrove fungal endophytes. Mycobiology 2015, 43, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Ponpandian, L.N.; Rim, S.O.; Shanmugam, G.; Jeon, J.; Park, Y.H.; Lee, S.K.; Bae, H. Phylogenetic characterization of bacterial endophytes from four Pinus species and their nematicidal activity against the pine wood nematode. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Calvo, H.; Mendiara, I.; Arias, E.; Gracia, A.P.; Blanco, D.; Venturini, M.E. Antifungal activity of the volatile organic compounds produced by Bacillus velezensis strains against postharvest fungal pathogens. Postharvest Biol. Technol. 2020, 166, 111208. [Google Scholar] [CrossRef]
- Zhang, K.; Bonito, G.; Hsu, C.M.; Hameed, K.; Vilgalys, R.; Liao, H.L. Mortierella elongata increases plant biomass among non-leguminous crop species. Agronomy 2020, 10, 754. [Google Scholar] [CrossRef]
- Ullah, I.; Khan, A.L.; Ali, L.; Khan, A.R.; Waqas, M.; Hussain, J.; Lee, I.J.; Shin, J.H. Benzaldehyde as an insecticidal, antimicrobial, and antioxidant compound produced by Photorhabdus temperata M1021. J. Microbiol. 2015, 53, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Kadota, H.; Ishida, Y. Production of Volatile Sulfur Compounds by Microorganisms. Annu. Rev. Microbiol. 1972, 26, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Birkinshaw, J.H.; Findlay, W.P.K.; Webb, R.A. Biochemistry of the wood-rotting fungi. Biochem. J. 1942, 36, 526–529. [Google Scholar] [CrossRef] [Green Version]
- Amich, J.; Schafferer, L.; Haas, H.; Krappmann, S. Regulation of Sulphur Assimilation Is Essential for Virulence and Affects Iron Homeostasis of the Human-Pathogenic Mould Aspergillus fumigatus. PLoS Pathog. 2013, 9, e1003573. [Google Scholar] [CrossRef] [Green Version]
- Amich, J. mSphere of Influence: The Importance of Metabolism for Pathogen Adaptation to Host-Imposed Stresses. mSphere 2019, 4, e00566-19. [Google Scholar] [CrossRef] [Green Version]
- Luque-Almagro, V.M.; Cabello, P.; Sáez, L.P.; Olaya-Abril, A.; Moreno-Vivián, C.; Roldán, M.D. Exploring anaerobic environments for cyanide and cyano-derivatives microbial degradation. Appl. Microbiol. Biotechnol. 2018, 102, 1067–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rijavec, T.; Lapanje, A. Hydrogen cyanide in the rhizosphere: Not suppressing plant pathogens, but rather regulating availability of phosphate. Front. Microbiol. 2016, 7, 1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tattini, M.; Loreto, F.; Fini, A.; Guidi, L.; Brunetti, C.; Velikova, V.; Gori, A.; Ferrini, F. Isoprenoids and phenylpropanoids are part of the antioxidant defense orchestrated daily by drought-stressed Platanus × acerifolia plants during Mediterranean summers. New Phytol. 2015, 207, 613–626. [Google Scholar] [CrossRef]
- McGenity, T.J.; Crombie, A.T.; Murrell, J.C. Microbial cycling of isoprene, the most abundantly produced biological volatile organic compound on Earth. ISME J. 2018, 12, 931–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, C.M.; Helmig, D.; Fierer, N. Bacteria and fungi associated with isoprene consumption in soil. Elementa 2015, 3, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, S.; Cain, R.B.; Rao, G.V.; Kirby, G.W. Isolation and identification of two novel butenolides as products of dimethylbenzoate metabolism by Rhodococcus rhodochrous N75. FEMS Microbiol. Lett. 1994, 120, 93–98. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Location | Season | Matrix | Accession | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Haggen | Kühtai | Obergugl | Patscherkofel | Pfitscherjoch | Praxmar | Snowcover | Exposed | Meshbags | Soil | Numbers | |
| M. alpina/globalpina | - | - | 12 | - | - | - | 8 | 4 | 12 | - | MT380876, MT380877, MT380878, MT380879, MT380880, KP714667, MF615092, MT380871, MT380872, MT380873, MT380874, MT380875 |
| M. angusta | - | 2 | - | 1 | 1 | 1 | 2 | 3 | - | 5 | MT380903, MT380904, MT380905, MT380906, MT380907 |
| M. bainieri | - | - | 1 | - | - | 5 | - | 6 | 1 | 5 | MT380861, MT380862, MT380863, MT380864, MT380865, MT380866 |
| L. exigua | - | 2 | - | 2 | - | 1 | 2 | 3 | - | 5 | MT380898, MT380899, MT380900, MT380901, MT380860 |
| L. gamsii | 1 | 1 | - | 1 | - | 2 | 1 | 4 | 1 | 4 | MT308707, MT308715, MT394054, MT308713, MT308709 |
| M. gemmifera | - | - | - | - | - | 2 | - | 2 | - | 2 | MT380867, MT380868, MT380869 |
| P. horticola | 5 | - | - | - | - | - | - | 5 | 5 | - | MT380886, MT380887, MT380888, MT380889, MT380890 |
| P. humilis/verticilata | 1 | 3 | - | 1 | 1 | 2 | 2 | 6 | 1 | 7 | MT380856, MT380857, MT380859, MT380881, MT380882, MT380883, MT380884, MT380885 |
| L. hyalina | - | 1 | - | 1 | - | 3 | 1 | 4 | - | 5 | MT308724, MT308721, MT308722, MT308717, MT308723 |
| E. parvispora | 1 | 4 | - | 1 | - | 1 | 5 | 2 | 1 | 6 | MT380891, MT380892, MT380893, MT380894, MT380895, MT380896, MT380897 |
| M. pseudozygospora | - | 2 | - | - | 2 | 1 | 2 | 3 | - | 5 | MT380851, MT380852, MT380853, MT380854, MT380855 |
| M. solitaria | - | - | 3 | - | - | - | - | 3 | 3 | - | MT279272, MT279273, MT279274 |
| M. zonata | - | - | - | 1 | 1 | 1 | 2 | 1 | - | 3 | MT308725, MT380870, MT308726 |
| Location | pH | CN | Organic [% Dry Matter] | Coordinates | Altitude Masl | Dominating Trees | Citation |
|---|---|---|---|---|---|---|---|
| Kühtai 3 | 4 | 24.3 | 30 | 47.217208, 11.036823 | 2030 | Pinus cembra | |
| Kühtai 2 | 3.5 | 19.9 | 70 | 47.208363, 11.006565 | 1880 | Pinus cembra | |
| Kühtai 1 | 3 | 19.3 | 60 | 47.214527, 10.991395 | 1910 | Pinus cembra | |
| Praxmar highest | 3 | 21.1 | 10 | 47.155964, 11.128367 | 1820 | Pinus cembra | [27] |
| Praxmar middle | 3.5 | 14.2 | 30 | 47.154348, 11.130158 | 1780 | Pinus cembra, Picea abies | |
| Praxmar lowest | 4 | 13.9 | 50 | 47.162253, 11.139553 | 1520 | Pinus cembra, Picea abies | |
| Haggen | 20.6 | 25 | 47.21256, 11.087578 | 2230 | Pinus cembra 35 years afforestation | [28] | |
| Pfitschtal | 4.5 | 35 | 46.994718, 11.668151 | 2261 | Salix retusa-reticulata snowbed communities | ||
| Obergurgl | 7.5 | 0,6 | 46°50′ N, 11°01′ E | 2400 | Bare ground, glacier forefield | [29,30] | |
| Patscherkofel | 3.5 | 22.5 | 70 | 4.9938158, 11.6629124 | 2260 | Pinus cembra | [31] |
| ms61.0295 | ms73.0649 | ms33.0339 | ms48.0534 | ms57.0404 | ms60.0527 | ms31.0183 | ms43.0182 | ms43.0545 | ms41.0384 | ms57.0702 | ms39.0226 | ms87.0445 | ms89.0599 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Formula | C2H4O2H+ | C4H8OH+ | CH4OH+ | Isotope of C2H6OH+ | C3H4OH+ | Isotope of C3H6OH+ | CH2OH+ | C2H3O+ | C3H7+ | C3H5+ | C4H9+ | C3H3+ | C4H6O2H+ | C4H8O2H+ |
| Chemical | Acid | Butanone | Methanol | Ethanol | General | Acetone | Form- | General | General | General | Butanol | General | Diacetyl | Ester |
| Name | Acetic acid | Butanal | fragment | aldehyde | fragment | fragment | fragment | fragment | Ethyl Acetate | |||||
| ALGL | 12 ± 5.6 | 11 ± 4.6 | 16 ± 4 | 2.2 ± 0.91 | 6 ± 0.74 | 8 ± 2.5 | 1.9 ± 0.54 | 9 ± 3.6 | 21 ± 4.8 | 15 ± 3.3 | 4 ± 5 | 19 ± 4.5 | 0.36 ± 0.069 | 5 ± 2.8 |
| AN | 8 ± 2.3 | 24 ± 6 | 12 ± 2.3 | 2.6 ± 0.89 | 6 ± 1.2 | 7 ± 1.5 | 2 ± 0.36 | 8 ± 4.1 | 24 ± 7.6 | 16 ± 4.9 | 4 ± 1.2 | 21 ± 6.5 | 0.5 ± 0.22 | 3 ± 2.1 |
| BA | 7 ± 5.3 | 20 ± 7.3 | 43 ± 16.4 | 1 ± 0.56 | 6.6 ± 0.71 | 8 ± 1.4 | 2.2 ± 0.28 | 6 ± 3.3 | 28 ± 4.6 | 21 ± 2.9 | 10 ± 2.4 | 28 ± 3.9 | 0.4 ± 0.32 | 2 ± 0.64 |
| EX | 6 ± 13.2 | 17 ± 5.7 | 17 ± 3.5 | 1.1 ± 0.48 | 5 ± 1.3 | 6 ± 2 | 1.7 ± 0.37 | 6 ± 7.2 | 17 ± 4.5 | 12 ± 3.1 | 2.9 ± 0.8 | 15 ± 4.3 | 0.5 ± 0.14 | 2.7 ± 0.99 |
| GA | 7 ± 16.5 | 16 ± 2.8 | 14 ± 5.5 | 1.2 ± 0.48 | 5.5 ± 0.64 | 8 ± 2.3 | 2.1 ± 0.49 | 6 ± 8.8 | 14 ± 3.6 | 11 ± 2.1 | 3.6 ± 0.77 | 15 ± 3.1 | 0.5 ± 0.34 | 2 ± 1 |
| GE | 9 ± 19.3 | 15 ± 6.5 | 16 ± 5.4 | 1 ± 1.1 | 6.6 ± 0.85 | 7 ± 1.4 | 2 ± 0.36 | 15 ± 8.1 | 11 ± 8.1 | 17 ± 5.2 | 3 ± 1.7 | 23 ± 10.6 | 0.7 ± 0.2 | 13 ± 4.2 |
| HO | 8 ± 8 | 13 ± 4.4 | 19 ± 4.1 | 0.8 ± 0.56 | 5.2 ± 0.72 | 7 ± 1.5 | 1.8 ± 0.29 | 7 ± 4.2 | 16 ± 3.8 | 12 ± 2.8 | 4 ± 1 | 17 ± 3.9 | 0.38 ± 0.096 | 2.1 ± 0.64 |
| HUVE | 8 ± 9.9 | 22 ± 7.4 | 15 ± 3.8 | 1 ± 0.42 | 6 ± 1.6 | 8 ± 2.2 | 1.9 ± 0.4 | 6 ± 4.4 | 18 ± 5.3 | 13 ± 3.5 | 3 ± 1.1 | 17 ± 4.8 | 0.4 ± 0.18 | 1.6 ± 0.71 |
| HY | 12 ± 6.4 | 9 ± 6 | 21 ± 9.7 | 1.8 ± 0.98 | 6.3 ± 0.92 | 7 ± 2.2 | 2 ± 0.43 | 10 ± 3.8 | 22 ± 6.9 | 16 ± 4.7 | 4 ± 1.3 | 21 ± 6.4 | 0.9 ± 0.6 | 6 ± 2.2 |
| PA | 7 ± 8.1 | 19 ± 10.5 | 10 ± 2.1 | 2 ± 72.2 | 6 ± 4.1 | 6 ± 2.8 | 2 ± 3.8 | 7 ± 15.5 | 19 ± 6.9 | 15 ± 18.5 | 4 ± 52.4 | 20 ± 30.4 | 0.5 ± 0.77 | 3 ± 5.1 |
| PZ | 11 ± 9.3 | 17 ± 5 | 13 ± 2.4 | 2 ± 1.5 | 6 ± 1.6 | 6 ± 1.7 | 1.8 ± 0.85 | 11 ± 5.3 | 23 ± 6.6 | 17 ± 4.9 | 3 ± 1.3 | 22 ± 8.4 | 1 ± 1.4 | 2 ± 3.3 |
| SO | 10 ± 3.1 | 6 ± 8.4 | 14 ± 2.1 | 1.5 ± 0.31 | 5.7 ± 0.67 | 6 ± 1.1 | 1.6 ± 0.27 | 8 ± 1.6 | 19 ± 5.4 | 15 ± 3.1 | 3 ± 0.79 | 20 ± 4.5 | 0.4 ± 0.24 | 5 ± 1.4 |
| ZO | 8 ± 41.9 | 13 ± 4 | 19 ± 9.3 | 0.9 ± 0.4 | 5.6 ± 0.59 | 7 ± 1.6 | 1.8 ± 0.29 | 6 ± 21.8 | 18 ± 4.5 | 13 ± 2.9 | 4 ± 1.1 | 17 ± 4.1 | 0.4 ± 0.16 | 2.1 ± 0.92 |
| kruskal | 9.3 × 10−5 | 2.1 × 10−14 | 2.3 × 10−14 | 5.7 × 10−13 | 4.8 × 10−4 | 1.0 × 10−3 | 9.7 × 10−3 | 1.2 × 10−6 | 5.4 × 10−11 | 1.2 × 10−10 | 2.8 × 10−8 | 4.1 × 10−10 | 1.4 × 10−6 | 6.9 × 10−13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Telagathoti, A.; Probst, M.; Khomenko, I.; Biasioli, F.; Peintner, U. High-Throughput Volatilome Fingerprint Using PTR–ToF–MS Shows Species-Specific Patterns in Mortierella and Closely Related Genera. J. Fungi 2021, 7, 66. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010066
Telagathoti A, Probst M, Khomenko I, Biasioli F, Peintner U. High-Throughput Volatilome Fingerprint Using PTR–ToF–MS Shows Species-Specific Patterns in Mortierella and Closely Related Genera. Journal of Fungi. 2021; 7(1):66. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010066
Chicago/Turabian StyleTelagathoti, Anusha, Maraike Probst, Iuliia Khomenko, Franco Biasioli, and Ursula Peintner. 2021. "High-Throughput Volatilome Fingerprint Using PTR–ToF–MS Shows Species-Specific Patterns in Mortierella and Closely Related Genera" Journal of Fungi 7, no. 1: 66. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7010066