
Hungry for Sex: Differential Roles for Ustilago maydisb Locus Components in Haploid Cells vis à vis Nutritional Availability

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Growth Conditions
2.2. Genetic Manipulation and Vector Construction
2.3. RNA Isolation and Expression Analysis
2.4. Mating Assay and Plant Pathogenesis
3. Results
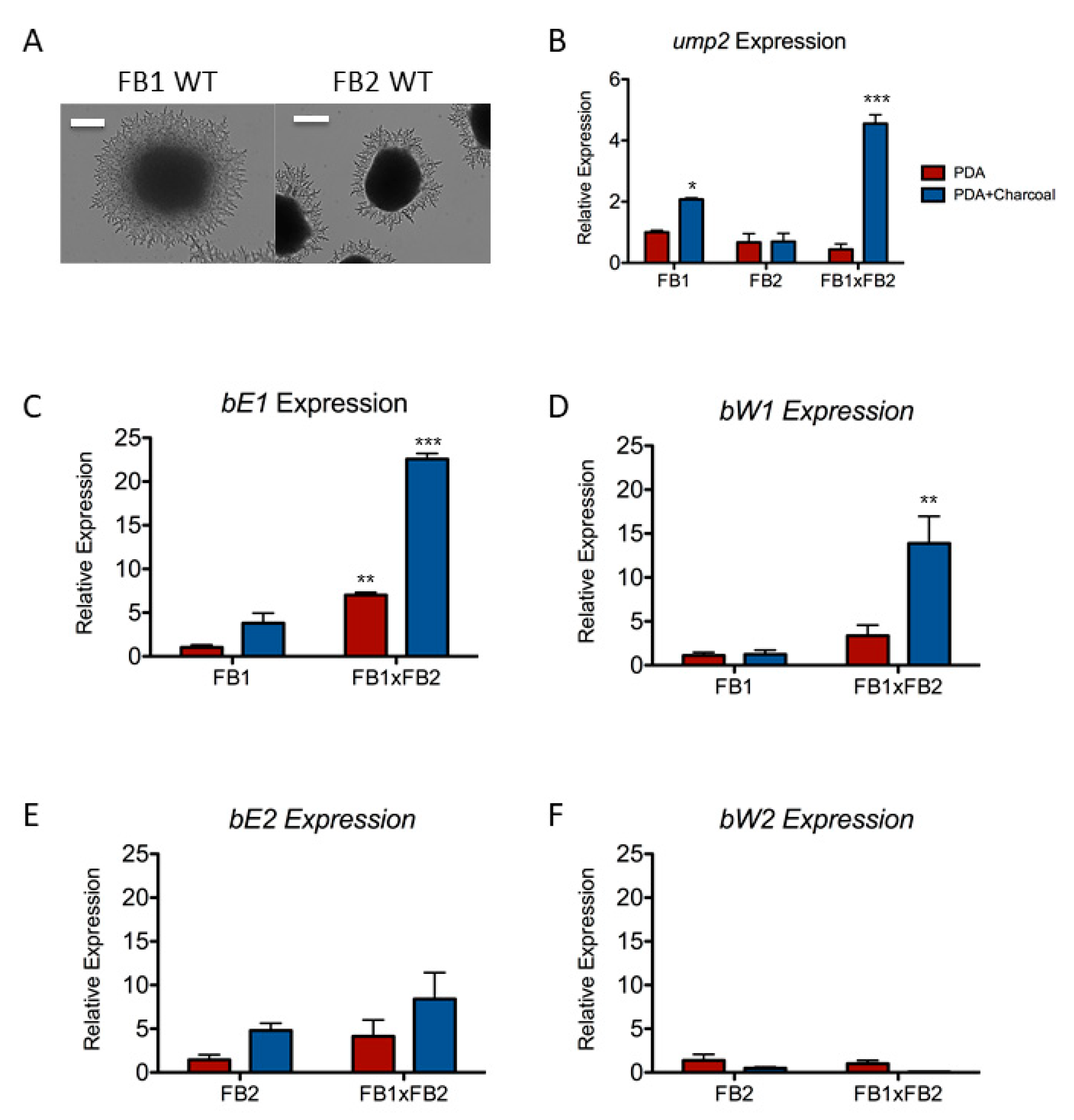
3.1. Under Mating Conditions, FB1 and FB2 Do Not Respond in the Same Way at the Level of Regulation of Gene Expression
3.2. The Components of the b Mating Type Locus Are Not Expressed Equally
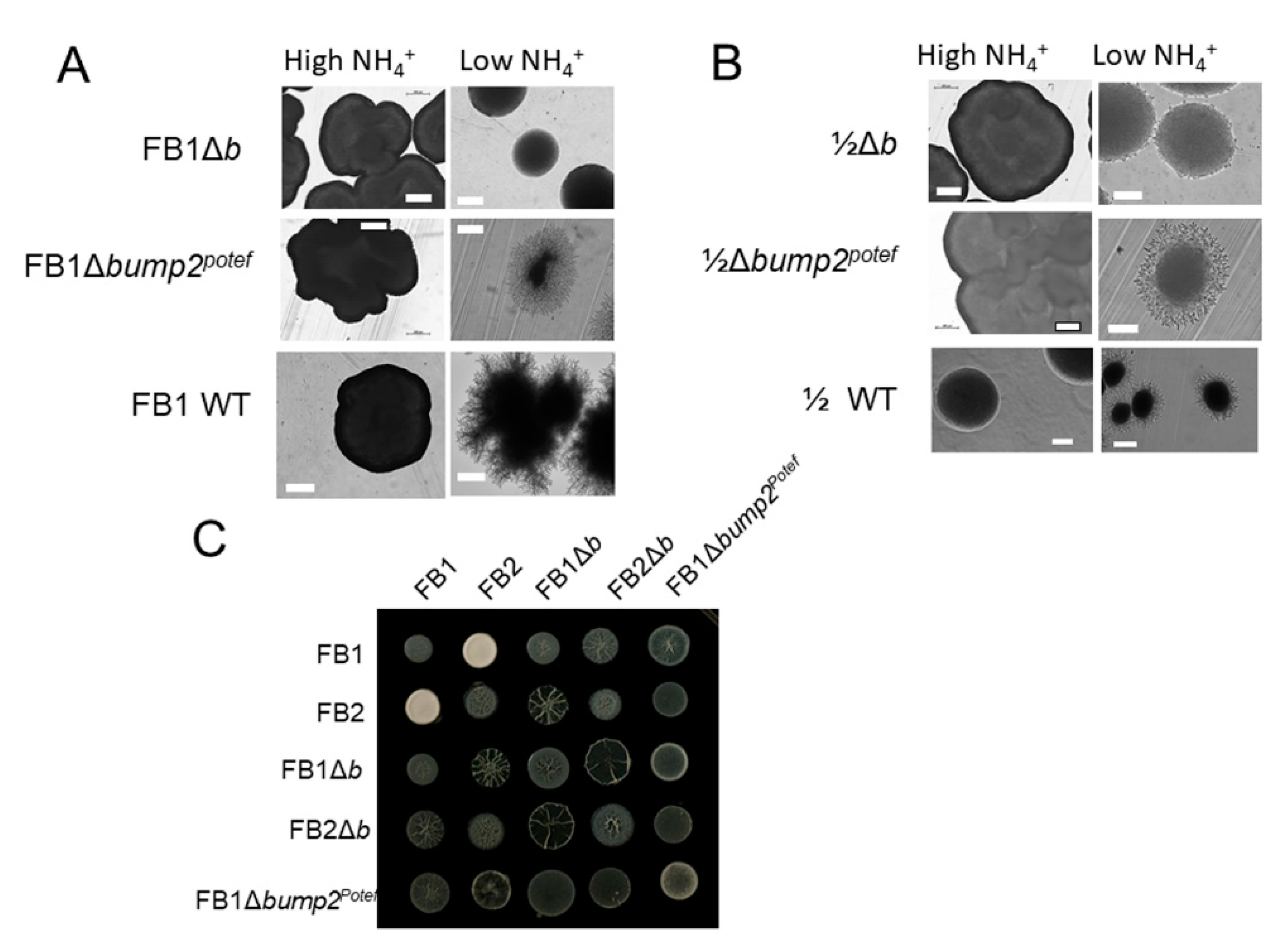
3.3. Overexpression of ump2 in Haploid FB1 Lacking the Entire b Mating Type Locus Rescues the Loss of Filamentous Phenotype in Response to Low NH4
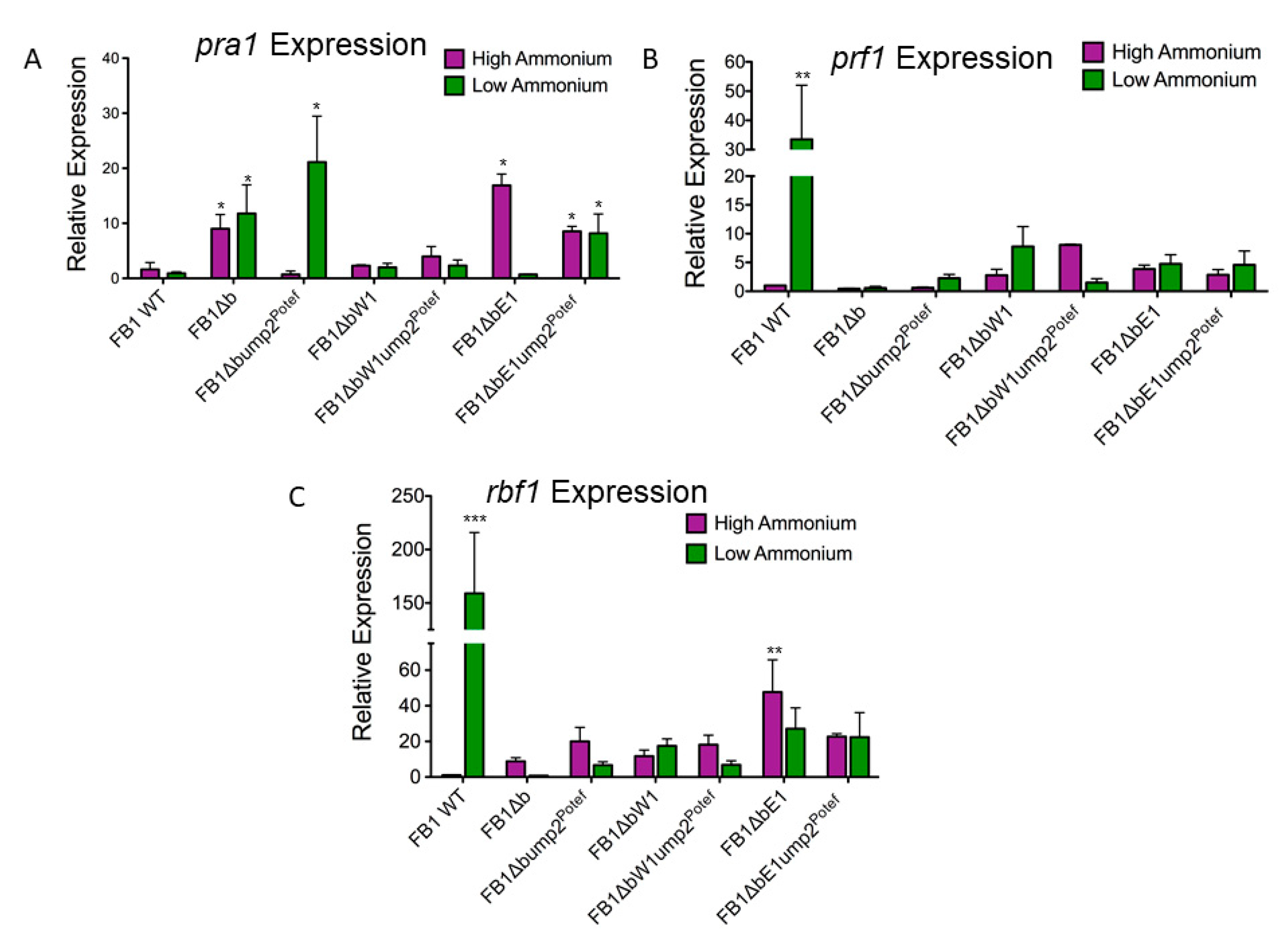
3.4. Haploid Strains Have Different Phenotypes under Low-NH4 Conditions When Only Part of the b Mating Type Locus Is Deleted
3.5. Regulation of Gene Expression Is Different in Partial b Mutants Overexpressing ump2
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kämper, J.; Friedrich, M.W.; Kahmann, R. Creating novel specificities in a fungal nonself recognition system by single step homologous recombination events. New Phytol. 2020, 228, 1001–1010. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, A.H.; Berta, P.; Palmer, M.S.; Hawkins, J.R.; Griffiths, B.L.; Smith, M.J.; Foster, J.W.; Frischauf, A.M.; Lovell-Badge, R.; Goodfellow, P.N. A gene from the human sex-determining region encodes a protein with homology to a conserved DNA-binding motif. Nature 1990, 346, 240–244. [Google Scholar] [CrossRef] [Green Version]
- Astell, C.R.; Ahlstrom-Jonasson, L.; Smith, M. The Sequence of the DNAs Coding for the Mating-Type Loci of Saccharomyces cerevisiae. Cell 1981, 27, 15–23. [Google Scholar] [CrossRef]
- Dranginis, A.M. Binding of Yeast a1 and (alpha)2 as a Heterodimer to the Operator DNA of a Haploid-Specific Gene. Nature 1990, 18, 682–685. [Google Scholar] [CrossRef] [PubMed]
- Yamada-Inagawa, T.; Klar, A.J.; Dalgaard, J.Z. Schizosaccharomyces pombe switches mating type by the synthesis-dependent strand-annealing mechanism. Genetics 2006, 177, 255–265. [Google Scholar] [CrossRef] [Green Version]
- Hood, M.E. Dimorpic Mating-Type Chromosomes in the Fungus Microbotryum violaceum. Genetics 2002, 160, 457–461. [Google Scholar]
- Hood, M.E.; Petit, E.; Giraud, T. Extensive divergence between mating-type chromosomes of the anther-smut fungus. Genetics 2013, 193, 309–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kronstad, J.W.; Leong, S. Isolation of two alleles of the b locus of Ustilago maydis. Proc. Natl. Acad. Sci. USA 1989, 86, 978–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, H.A.; Kahmann, R.; Bolker, M. The pheromone response factor coordinates filamentous growth and pathogenicity in Ustilago maydis. EMBO J. 1996, 15, 1632–1641. [Google Scholar] [CrossRef] [PubMed]
- Banuett, F.; Herskowitz, I. Morphological Transitions in the Life Cycle of Ustilago maydis and Their Genetic Control by the a and b Loci. Exp Mycol. 1995, 18, 247–266. [Google Scholar] [CrossRef]
- Paul, J.A.; Wallen, R.M.; Zhao, C.; Shi, T.; Perlin, M.H. Coordinate regulation of Ustilago maydis ammonium transporters and genes involved in mating and pathogenicity. Fungal Biol. 2018, 122, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Heistinger, L.; Moser, J.; Tatto, N.E.; Valli, M.; Gasser, B.; Mattanovich, D. Identification and characterization of the Komagataella phaffii mating pheromone genes. FEMS Yeast Res. 2018, 18, foy051. [Google Scholar] [CrossRef] [PubMed]
- Holliday, R. Ustilago maydis. In Handbook of Genetics; King, R.C., Ed.; Plenum Press: New York, NY, USA, 1974; pp. 575–595. [Google Scholar]
- Gold, S.E.; Brogdon, S.M.; Mayorga, M.; Kronstad, J.W. The Ustilago maydis regulatory subunit of a cAMP-dependent protein kinase is required for gall formation in maize. Plant Cell 1997, 9, 1585–1594. [Google Scholar]
- Gillissen, B.; Bergemann, J.; Sandmann, C.; Schroeer, B.; Bölker, M.; Kahmann, R. A Two-Component Regulatory System for Self/Non-Self Regulation in Ustilago maydis. Cell 1992, 68, 647–657. [Google Scholar] [CrossRef] [Green Version]
- Brachmann, A.; Konig, J.; Jullus, C.; Feldbrügge, M. A reverse genetic approach for generating gene replacement mutants in Ustilago maydis. Mol. Genet. Genom. 2004, 272, 216–226. [Google Scholar] [CrossRef]
- Smith, D.G.; Garcia-Pedrajas, M.D.; Gold, S.E.; Perlin, M.H. Isolation and characterization from pathogenic fungi of genes encoding ammonium permeases and their roles in dimorphism. Mol. Microbiol. 2003, 50, 259–275. [Google Scholar] [CrossRef]
- Weber, I.; Abmann, D.; Thines, E.; Steinberg, G. Polar Localizing Class V Myosin Chitin Synthases Are Essential during Early Plant Infection in the Plant Pathogenic Fungus Ustilago maydis. Plant Cell 2006, 18, 225–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brachmann, A.; Weinzieri, G.; Kämper, J.; Kahmann, R. Identification of genes in the bW/bE regulatory cascade in Ustilago maydis. Mol. Genet. Genom. 2001, 271, 102–110. [Google Scholar] [CrossRef]
- Kämper, J. A PCR-based system for highly efficient generation of gene replacement mutants in Ustilago maydis. Mol. Genet. Genom. 2004, 271, 103–110. [Google Scholar] [CrossRef]
- Perlin, M.H.; Amselem, J.; Fontanillas, E.; Toh, S.S.; Chen, Z.; Goldberg, J.; Duplessis, S.; Henrissat, B.; Young, S.; Zeng, Q.; et al. Sex and parasites: Genomic and transcriptomic analysis of Microbotryum lychnidis-dioicae, the biotrophic and plant-castrating anther smut fungus. BMC Genom. 2015, 16, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Banuett, F.; Herskowitz, I. Different a alleles of Ustilago maydis are necessary for maintenance of filamentous growth but not for meiosis. Proc. Natl. Acad. Sci. USA 1989, 86, 5878–5882. [Google Scholar] [CrossRef] [Green Version]
- Daniel, W.W. Kruskal–Wallis one-way analysis of variance by ranks. In Applied Nonparametric Statistics, 2nd ed.; PWS-Kent: Boston, MA, USA, 1990; pp. 226–234. [Google Scholar]
- Iyobe, T.; Asada, T.; Kawata, K.; Oikawa, K. Comparison of Removal Efficiencies for Ammonia and Amine Gases between Woody Charcoal and Activated Carbon. J. Health Sci. 2004, 50, 148–153. [Google Scholar]
- Snetselaar, K.M.; Mims, C.W. Sporidial Fusion and Infection of Maize Seedlings by the Smut Fungus Ustilago Maydis. Mycologiam 1992, 84, 193–203. [Google Scholar] [CrossRef]
- Bolker, M.; Urban, M.; Kahmann, R. The a Mating Type Locus of U. maydis Specifies Cell Signaling Components. Cell 1992, 8, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Heimel, K.; Scherer, M.; Schuler, D.; Kämper, J. The Ustilago maydis Clp1 protein orchestrates pheromone and b-dependent signaling pathways to coordinate the cell cycle and pathogenic development. Plant Cell 2010, 22, 2908–2922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimel, K.; Scherer, M.; Vranes, M.; Wahl, R.; Pothiratana, C.; Schuler, D.; Vincon, V.; Finkernagel, F.; Flor-Parra, I.; Kämper, J. The transcription factor Rbf1 is the master regulator for b-mating type controlled pathogenic development in Ustilago maydis. PLoS Pathog. 2010, 6, e1001035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merchan, F.; van den Ende, H.; Fernandez, E.; Beck, C.F. Low-expression genes induced by nitrogen starvation and subsequent sexual differentiation in Chlamydomonas reinhardtii, isolated by the differential display technique. Planta 2001, 213, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Ekwall, K.; Thon, G. Selecting Schizosaccharomyces pombe Diploids. Cold Spring Harb. Protoc. 2017. [Google Scholar] [CrossRef] [PubMed]
- McCann, M.P.; Snetselaar, K.M. A genome-based analysis of amino acid metabolism in the biotrophic plant pathogen Ustilago maydis. Fungal Genet. Biol. 2008, 45, S77–S87. [Google Scholar] [CrossRef] [PubMed]
- Stakman, E.C.; Kernkamp, M.F.; King, T.H.; Martin, W.J. Genetics Factors for Mutability and Mutant Characteristics in Ustilago zeae. Am. J. Bot. 1943, 37–48. [Google Scholar] [CrossRef]
- Lanver, D.; Müller, A.N.; Happel, P.; Schweizer, G.; Haas, F.B.; Franitza, M.; Pellegrin, C.; Reissmann, S.; Altmüller, J.; Rensing, S.A.; et al. The Biotrophic Development of Ustilago maydis Studied by RNA-Seq Analysis. Plant Cell 2018, 30, 300–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yee, A.R.; Kronstad, J.W. Dual Sets of Chimeric Alleles Identify Specificity Sequences for the bE and bW Mating and Pathogenicity Genes of Ustilago maydis. Mol. Cell Biol. 1998, 18, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Reference |
|---|---|---|
| FB1 WT | a1 b1 | [10] |
| FB1Δb | a1 b1 bE/W::hygR | [11] |
| FB1Δb ump2Otef | a1 b1 bE/W::hygR Potefump2, cbxR | This Study |
| FB2 WT | a2 b2 | [10] |
| 1/2 WT | a1 b1 | [14] |
| 2/9 WT | a2 b2 | [14] |
| RK1613 | a1 bE1 bW1::hygR | [15] |
| RK1725 | a1 bW1 bE1::hygR | [15] |
| RK1613 ump2Otef | a1 bE1 bW1::hygR Potefump2, cbxR | This study |
| RK1725 ump2Otef | a1 bW1 bE1::hygR Potefump2, cbxR | This study |
| Gene Name | Primers | Sequences (5′→3′) |
|---|---|---|
| Guanine nucleotide exchange factor (um04869) | rt-eif-2B-F | CAAATGCGATCCCGAACAG |
| rt-eif-2B-R | GGGACACCACTTGTCAAGCA | |
| ump2 (um05889) | rt-Ump2-F | TGGGTCCCGTTCTCATTTTC |
| rt-Ump2-R | AGGCGATGGGATTGTAGACAA | |
| prf1 (um02713) | rt-Prf1-F | CAGCACCAAGGTGGAAAGGT |
| rt-Prf1-R | GAATTGCCACGTGTTTGCAA | |
| mfa1 (um02382) | rt-Mfa1-F | ATGCTTTCGATCTTCGCTCAG |
| rt-Mfa1-R | TAGCCGATGGGAGAACCGT | |
| bE1 (um00577) | b1E1ft123 | GCAACAAAAGATACCCAACGA |
| b1E1rt334 | TTCGACACCCTACATCAGGAC | |
| bW1 (um00578) | b1W1ft1559 | TCGAGTCTGCCTCAATTCCT |
| b1W1rt1827 | CTCTCCTATGCTGGCTCCAC | |
| bE2 | bE2 fw bE2 rv | CAGGGGCAATAGGAAAGTCA ACCATTTTCGACCTCGTCAG |
| bW2 | bW2F bW2R | CGTGGAGCCTACGGAATCA GGAGGTGACTCGTGTCTGGAA |
| rbf1 (um03172) | RT Rbf1 F | AGGGTGTGGCAAATCGTTCT |
| RT Rbf1 R | TCGGCATCAGCATGGTTTC | |
| pra1 (um02383) | Pra1 qrt F | ACTCGATGGTCTGGTGGAAG |
| Pra1 qrt R | CTCACGCTCAATTCGCAATA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wallen, R.M.; Richardson, K.; Furnish, M.; Mendoza, H.; Dentinger, A.; Khanal, S.; Perlin, M.H. Hungry for Sex: Differential Roles for Ustilago maydisb Locus Components in Haploid Cells vis à vis Nutritional Availability. J. Fungi 2021, 7, 135. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7020135
Wallen RM, Richardson K, Furnish M, Mendoza H, Dentinger A, Khanal S, Perlin MH. Hungry for Sex: Differential Roles for Ustilago maydisb Locus Components in Haploid Cells vis à vis Nutritional Availability. Journal of Fungi. 2021; 7(2):135. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7020135
Chicago/Turabian StyleWallen, R. Margaret, Kirsten Richardson, Madison Furnish, Hector Mendoza, Allison Dentinger, Sunita Khanal, and Michael H. Perlin. 2021. "Hungry for Sex: Differential Roles for Ustilago maydisb Locus Components in Haploid Cells vis à vis Nutritional Availability" Journal of Fungi 7, no. 2: 135. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7020135