
Wheat Resistance to Stripe and Leaf Rusts Conferred by Introgression of Slow Rusting Resistance Genes

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Evaluation of Wheat Genotypes Response to Stripe and Leaf Rusts
2.1.1. At Seedling Stage
2.1.2. At Adult Stage
2.2. Molecular Markers Experiment
2.2.1. DNA Isolation and PCR Protocol
2.2.2. Introgression of Resistant Genes for Stripe and Leaf Rusts in Wheat Genotypes
2.2.3. Molecular Markers of F2 Plants
2.3. Statistical Analysis
3. Results
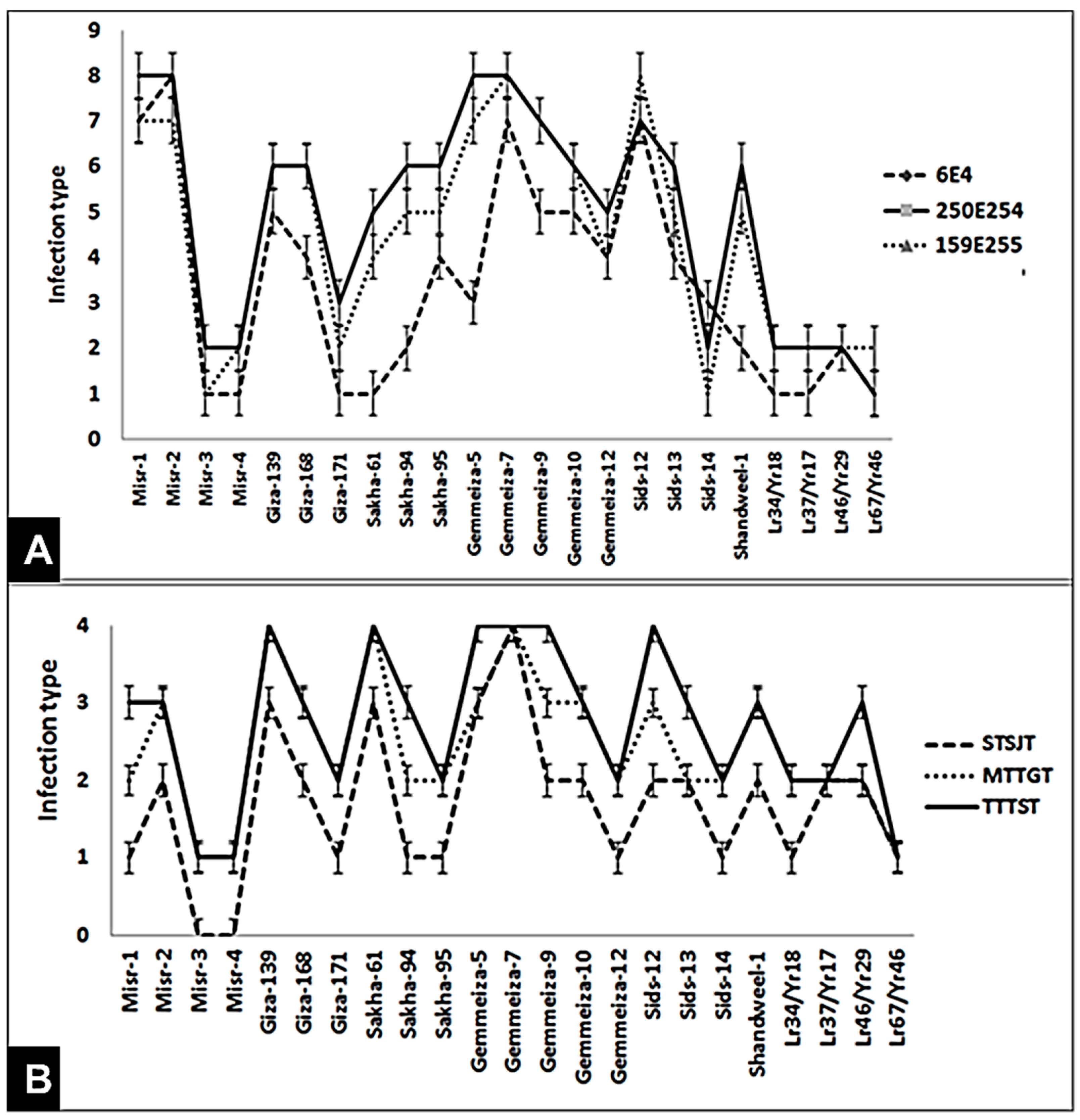
3.1. Evaluation of Genotypes against Stripe and Leaf Rusts at Seedling Stage
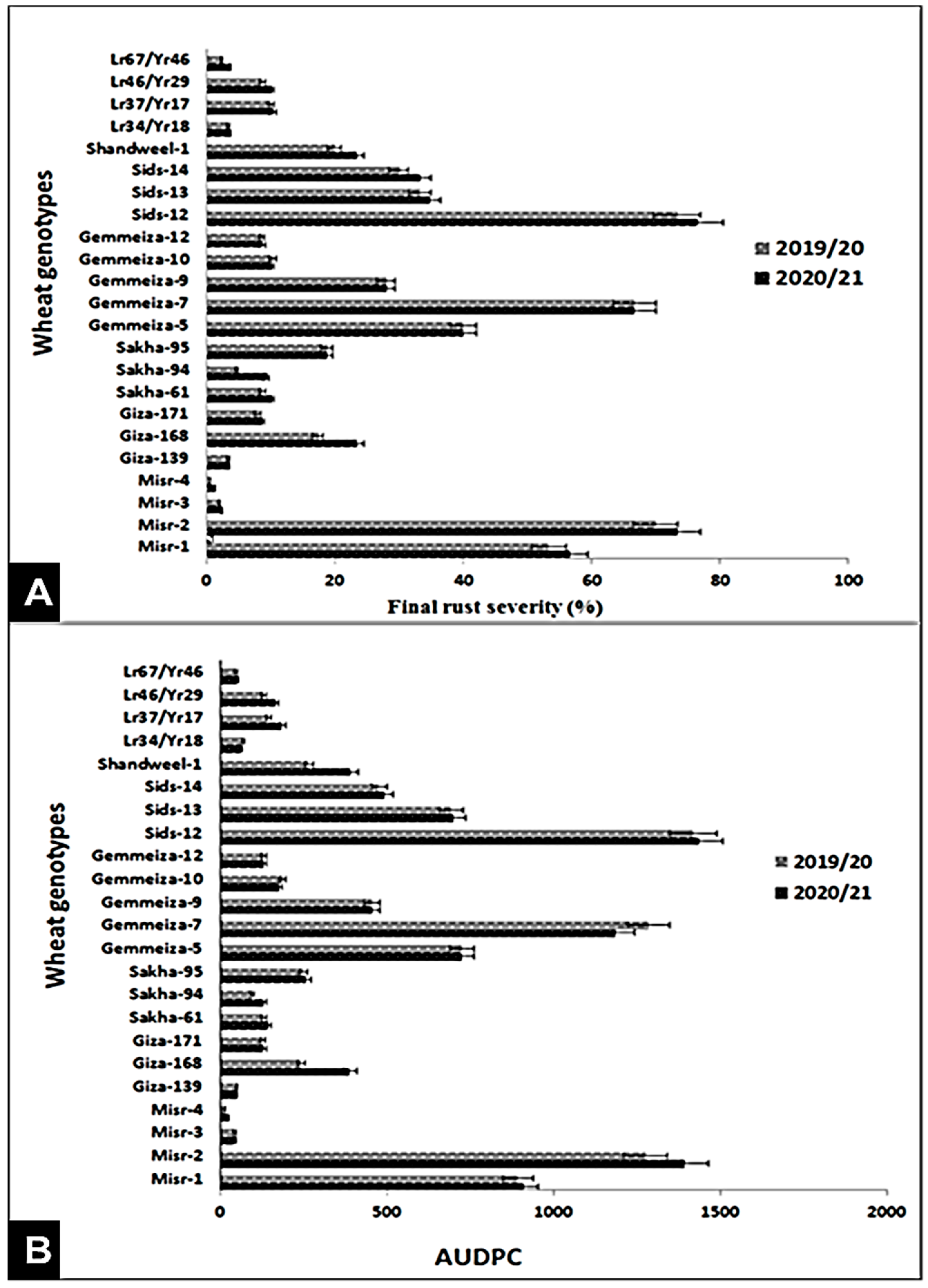
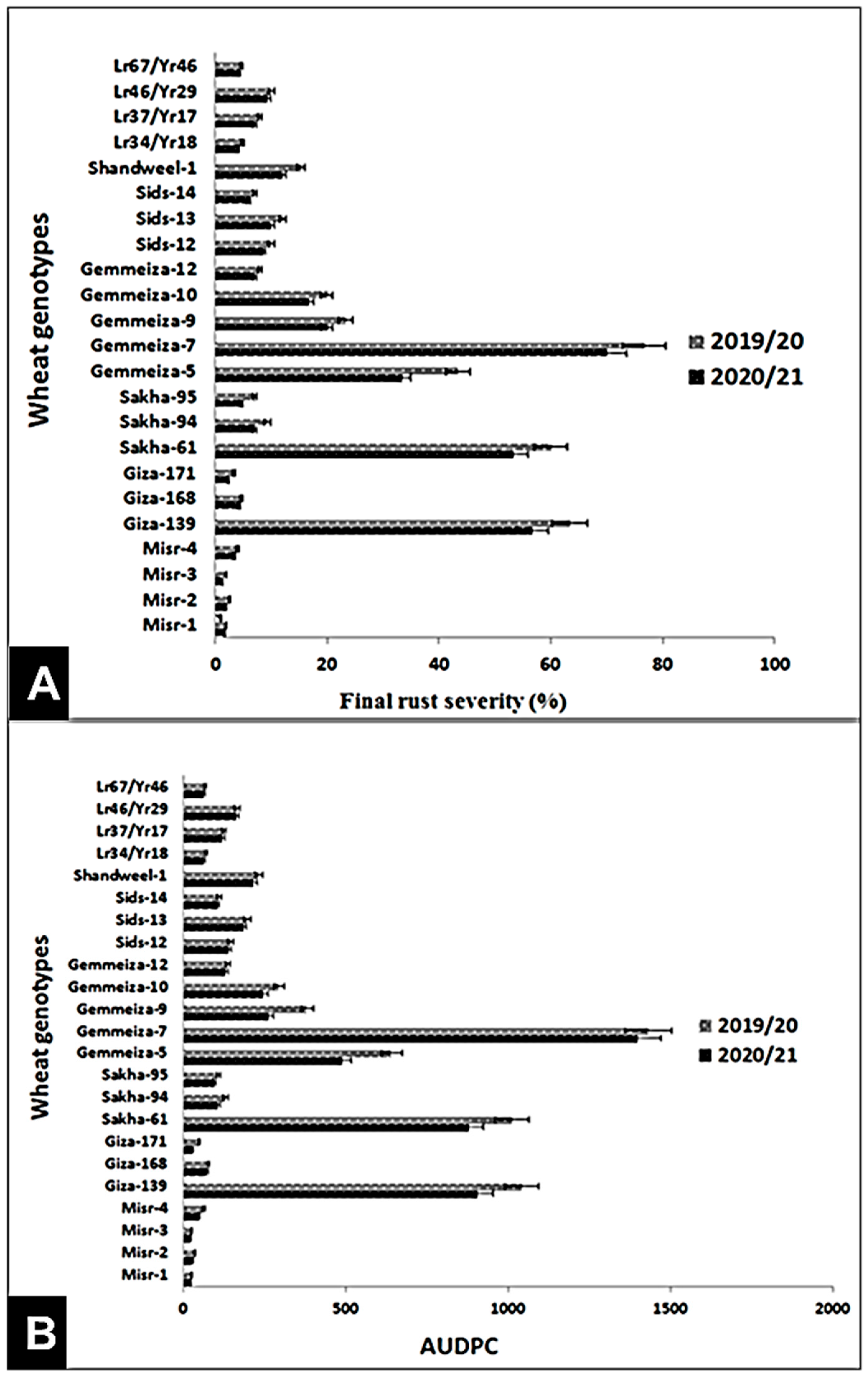
3.2. Evaluation of Genotypes against Stripe and Leaf Rusts under Field Conditions
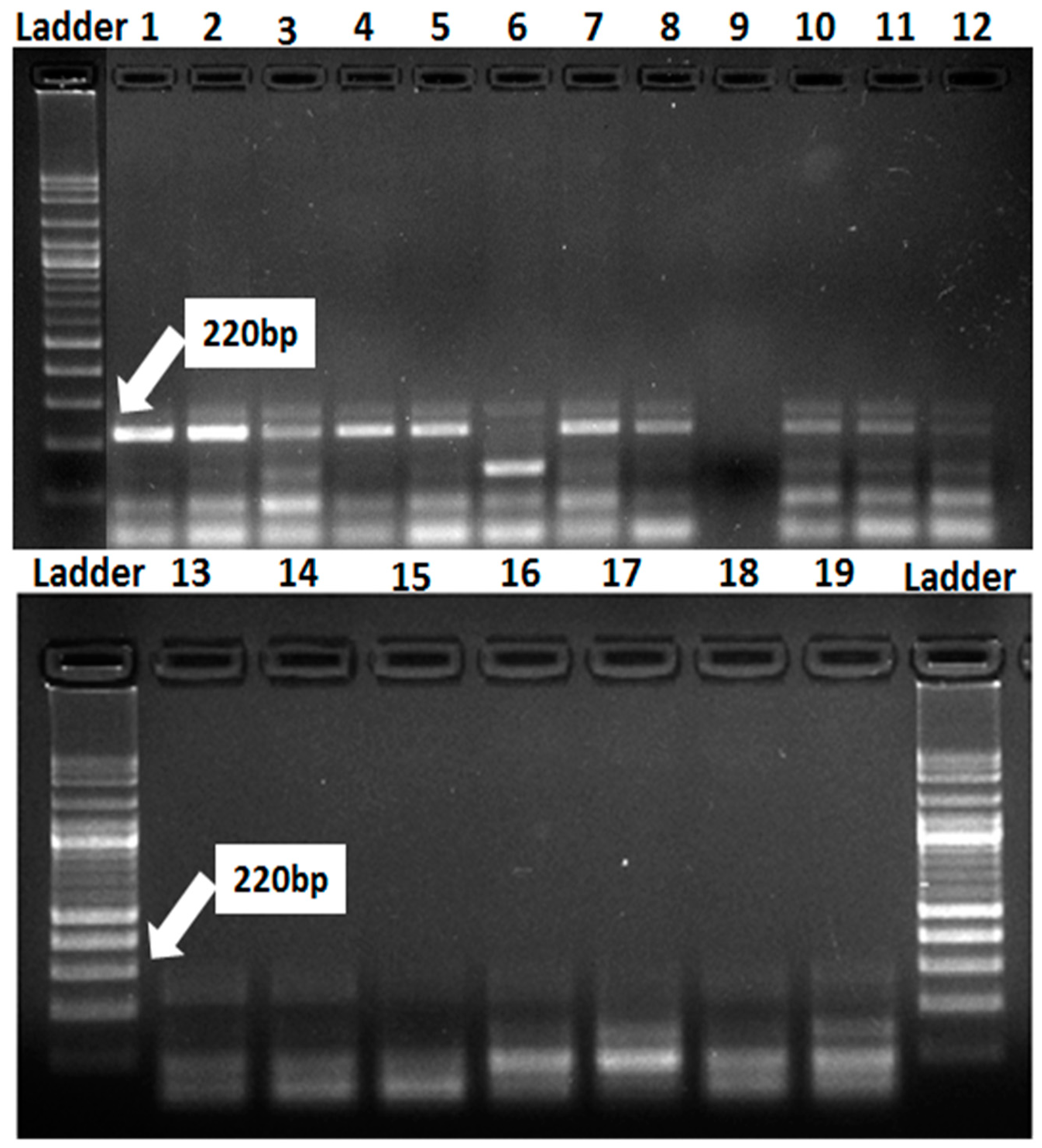
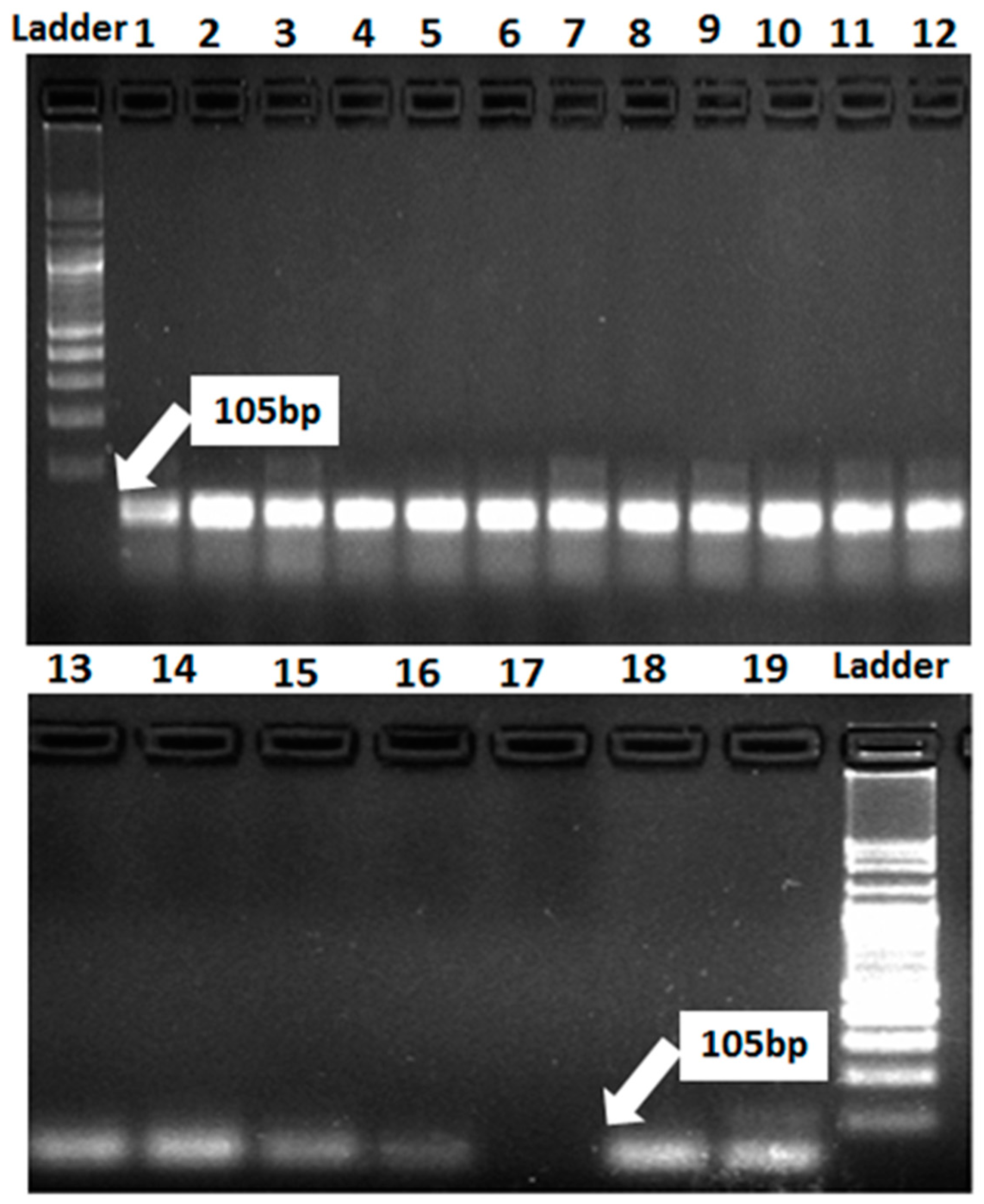
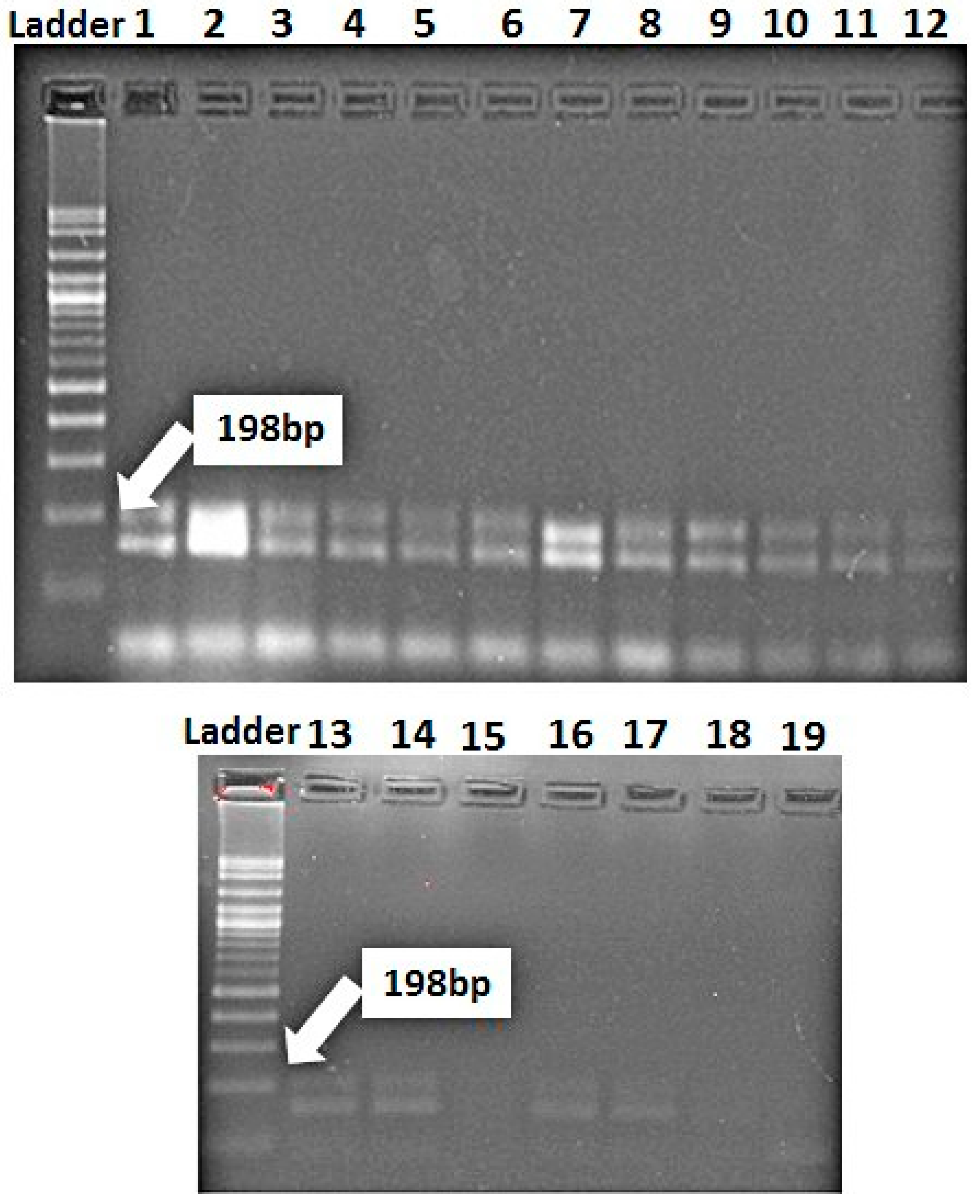
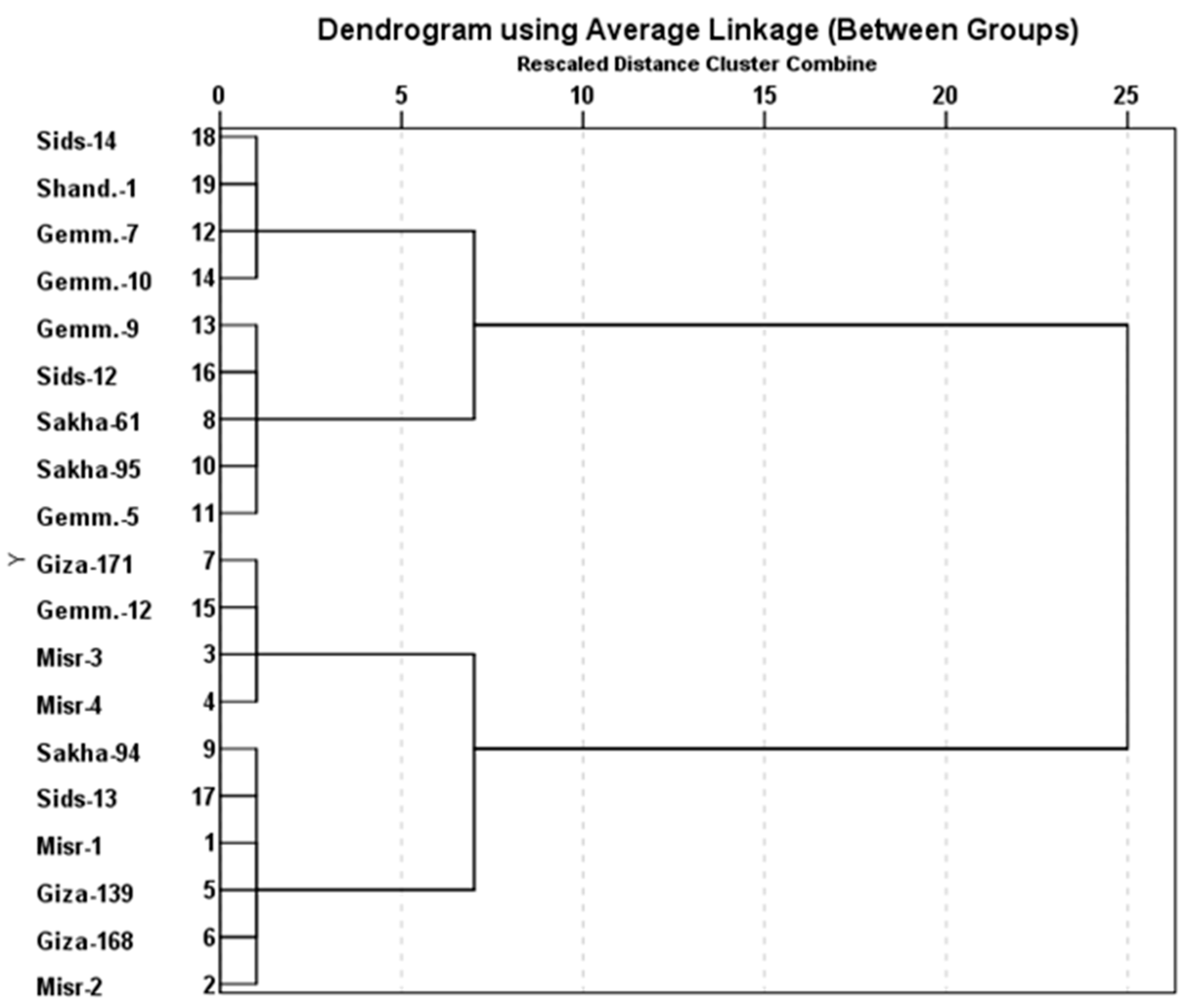
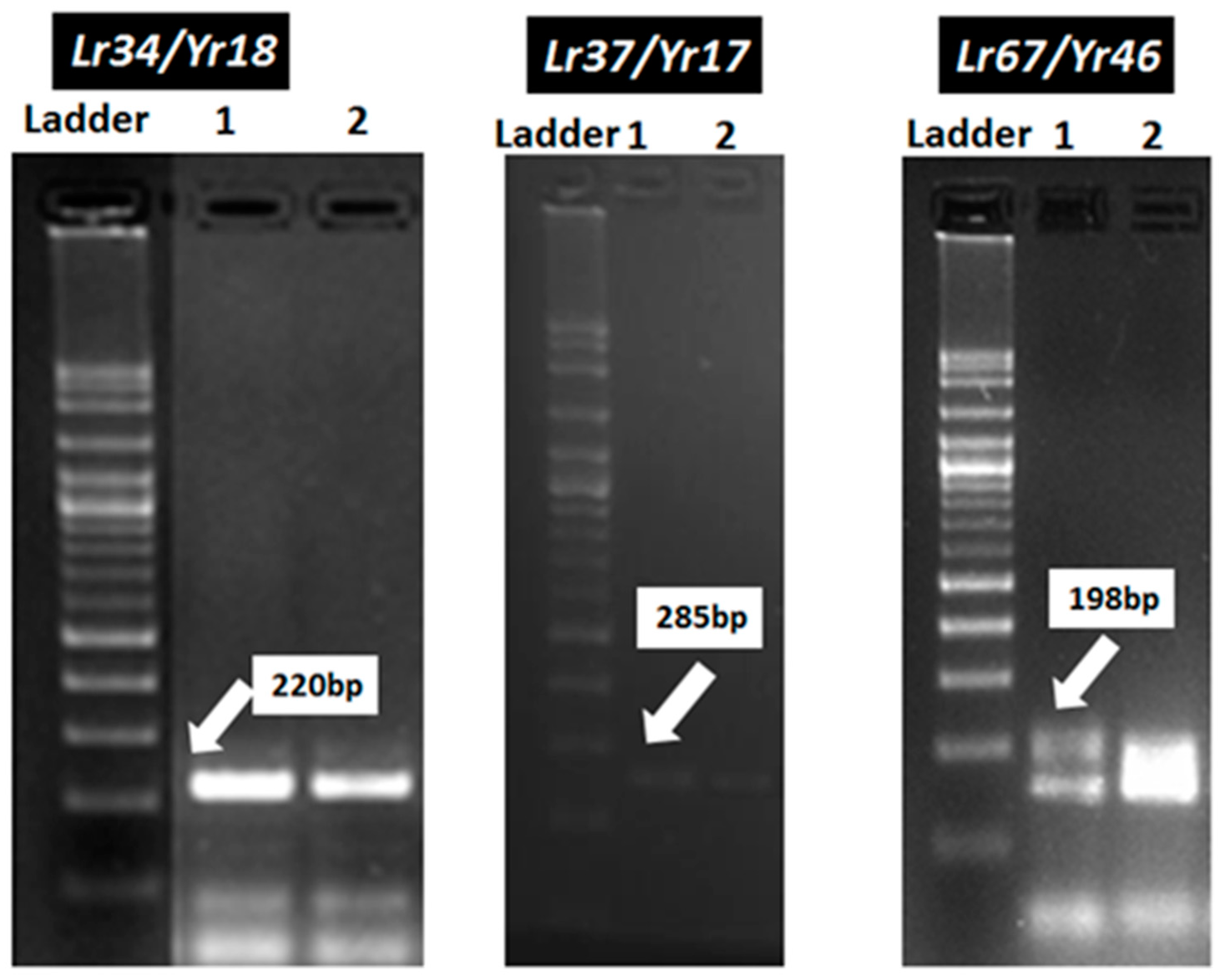
3.3. Identification of Resistance Genes of Stripe and Leaf Rusts in Wheat Genotypes Using Molecular Markers
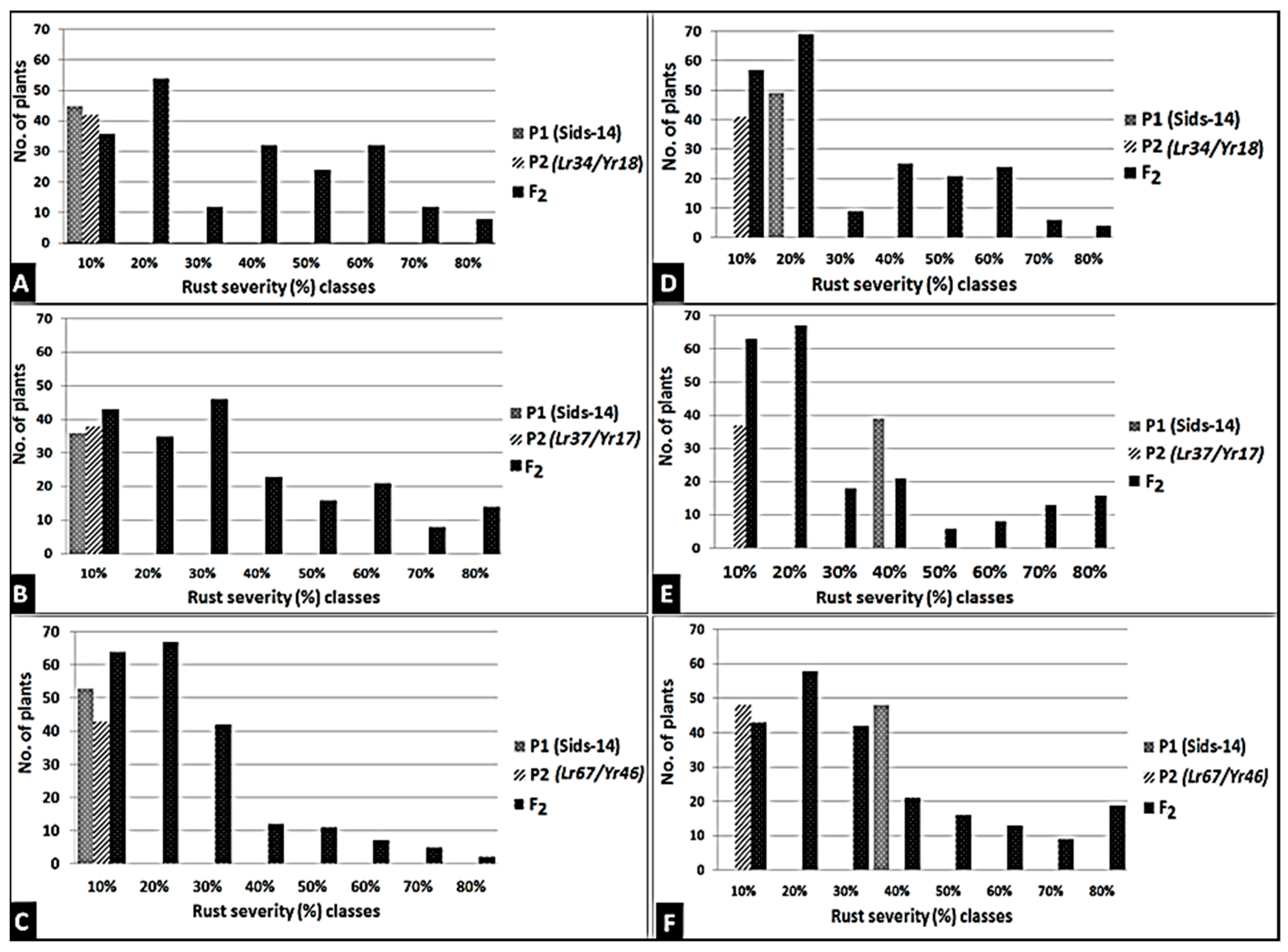
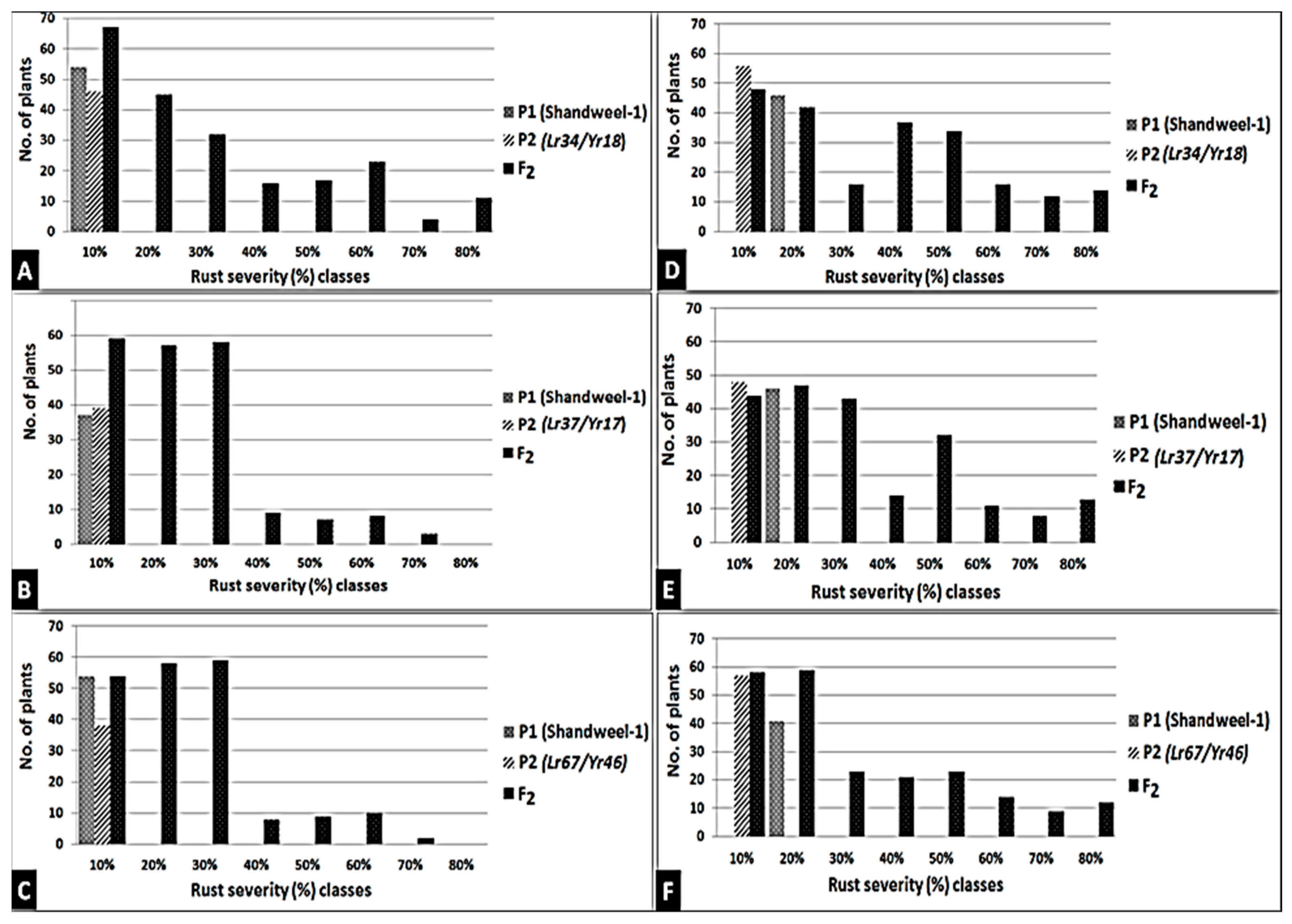
3.4. Introgression of Three Slow-Rusting Genes into the Two Wheat Genotypes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ali, R.G.; Omara, R.I.; Ali, Z.A. Effect of leaf rust infection on yield and technical properties in grains of some Egyptian wheat cultivars. Menoufia J. Plant Prot. 2016, 1, 19–35. [Google Scholar] [CrossRef]
- Omara, R.I.; El-Naggar, D.R.; Abd El-Malik, N.I.; Ketta, H.A. Losses assessment in some Egyptian wheat cultivars caused by stripe rust pathogen (Puccinia striiformis). Egypt J. Phytopathol. 2016, 44, 199–203. [Google Scholar] [CrossRef]
- Gill, H.S.; Li, C.; Sidhu, J.S.; Liu, W.; Wilson, D.; Bai, G.; Gill, B.S.; Sehgal, S.K. Fine mapping of the wheat leaf rust resistance gene Lr42. Int. J. Mol. Sci. 2019, 20, 2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamaludddin, M.; Khan, M.A.; Khan, M.A.; Alam, P.; Ahmed, S.; Abdin, M.Z.; Khan, M.N.; Bhatt, M.A. Genetic diversity analysis and development of SCAR marker associated with stripe rust resistance in spring wheat (Triticum aestivum L. em Thell). Indian J. Biotechnol. 2014, 13, 9–18. [Google Scholar]
- Asif, M.; Rahman, M.; Zafar, Y. Fingerprinting studies of some wheat (Triticum aestivum L.) genotypes using random amplified polymorphic DNA (RAPD) analysis. Pak. J. Bot. 2005, 37, 271–277. [Google Scholar]
- McIntosh, R.A.; Yamazaki, Y.; Dubcovsky, J.; Rogers, W.J.; Morris, C.; Appels, R.; Xia, X.C. Catalogue of Gene Symbols for Wheat. In Proceedings of the 12th International Wheat Genetics Symposium, Yokohama, Japan, 8–14 September 2013; pp. 8–13. [Google Scholar]
- Mebrate, S.A.; Dehne, H.; Pillen, K.; Oerka, E. Postulation of seedling leaf rust resistance genes in selected Ethiopian and German bread wheat cultivars. Crop Sci. 2008, 48, 507–511. [Google Scholar] [CrossRef]
- Khan, M.K.; Pandey, A.; Choudhary, S.; Hakki, E.E.; Akkaya, M.S.; Thomas, G. From RFLP to DArT: Molecular tools for wheat (Triticum spp.) diversity analysis. Genet. Resour. Crop Evol. 2014, 61, 1001–1032. [Google Scholar] [CrossRef]
- Khan, M.K.; Pandey, A.; Thomas, G.; Akkaya, M.S.; Kayis, S.A.; Ozsensoy, Y.; Ozsensoy, Y.; Hamurcu, M.; Gezgin, S.; Topal, A.; et al. Genetic diversity and population structure of wheat in India and Turkey. AoB Plants 2015, 7, plv083. [Google Scholar] [CrossRef]
- Bryan, G.J.; Collins, A.J.; Stephenson, P.; Orry, A.; Smith, J.B.; Gale, M.D. Isolation and characterization of microsatellites from hexaploid bread wheat. Theor. Appl. Genet. 1997, 94, 557–563. [Google Scholar] [CrossRef]
- Stakman, E.C.; Stewari, D.M.; Loegering, W.Q. Identification of physiologic races of Puccinia graminis var. tritici. USDA Agric. Res. Serv. Bull 1962, E-617, 53. [Google Scholar]
- McNeal, F.H.; Konzak, C.F.; Smith, E.P.; Tate, W.S.; Russell, T.S. A uniform system for recording and processing cereal data. Agric. Res. Serv. Bull. 1971, 34, 121–143. [Google Scholar]
- Das, M.K.; Rajaram, S.; Kronstad, W.E.; Mundt, C.C.; Singh, R.P. Association and genetics of three components of slow rusting in leaf rust of wheat. Euphytica 1993, 68, 99–109. [Google Scholar] [CrossRef]
- Shaner, G.; Finney, R.E. The effect of nitrogen fertilization on the expression of slow-mildewing resistance in Knox wheat. Phytopathology 1977, 67, 1051–1056. [Google Scholar] [CrossRef] [Green Version]
- Rogers, S.O.; Bendich, A.J. Extraction of total cellular DNA from plants, algae and fungi. In Plant Molecular Biology Manual, 2nd ed.; Gelvin, S.B., Schilperoort, R.A., Eds.; Kluwer Academic Press: Dordrecht, Germany, 1994; Volume D1, pp. 1–8. [Google Scholar]
- Tervet, I.; Cassel, R.C. The use of cyclone separation in race identification of cereal rusts. Phytopathology 1951, 41, 282–285. [Google Scholar]
- Abd El-Hamid, E.A.M. Genetic analysis of some bread crosses under normal and water stress conditions. Egypt. J. Plant Breed. 2013, 17, 42–56. [Google Scholar]
- Khaled, M.A.; Abd El-Dayem, S.M. Combining ability analysis for grain yield and its attributes in bread wheat under stress and normal irrigation conditions. J. Plant Prod. Mansoura Univ. 2014, 5, 255–266. [Google Scholar] [CrossRef]
- Kou, Y.; Wang, S. Broad-spectrum and durability: Understanding of quantitative disease resistance. Curr. Opin. Plant Biol. 2010, 13, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Abdelaal, K.A.A.; Hafez, Y.M.; Badr, M.M.; Youseef, W.A.; Esmail, S.M. Biochemical, histological and molecular changes in susceptible and resistant wheat cultivars inoculated with stripe rust fungus Puccinia striiformis f.sp. tritici. Egyp. J. Biol. Pest Cont. 2014, 24, 421–429. [Google Scholar]
- McVey, D.V.; Nazim, M.; Leonard, K.J.; Long, D.L. Patterns of virulence diversity in Puccinia triticina on wheat in Egypt and the United States in 1998–2000. Plant. Dis. 2004, 88, 271–279. [Google Scholar] [CrossRef] [Green Version]
- Esmail, S.M.; Omara, R.I.; Abdelaal, K.A.A.; Hafez, Y.M. Histological and biochemical aspects of compatible and incompatible wheat- Puccinia striiformis interactions. Physiol. Mol. Plant. Pathol. 2019, 106, 120–128. [Google Scholar] [CrossRef]
- Ashmawy, M.A.; Shahin, A.A.; Esmail, S.M.; Abd El-Naby, H. Virulence dynamics and diversity of Puccinia striiformis populations in Egypt during 2017/18 and 2018/19 growing seasons. J. Plant. Prot. Pathol. Mansoura Univ. 2019, 10, 655–666. [Google Scholar]
- Abdelbacki, A.M.M.; Omara, R.I.; Najeeb, M.A.; Soliman, N.E. Identification of leaf rust resistant genes Lr9, Lr25, Lr28, Lr29 and Lr67 in ten Egyptian wheat cultivars using molecular markers. Int. J. Biotechnol. Res. 2014, 7, 89–96. [Google Scholar]
- Abdelbacki, A.M.M.; Omara, R.I.; Soliman, N.E.; Najeeb, M.A. Molecular markers identification of leaf rust resistance genes LR19, LR21, LR24, LR47 and LR51 in selected egyptian wheat cultivars. Int. J. Phytopathol. 2015, 04, 55–62. [Google Scholar] [CrossRef]
- El-Orabey, W.M.; Sallam, M.E.; Omara, R.I.; Abd El-Malik Nagwa, I. Geographical distribution of Puccinia triticina physiologic races in Egypt during 2012–2014 growing seasons. Afr. J. Agric. Res. 2015, 10, 4193–4203. [Google Scholar]
- El-Orabey, W.M.; Omara, R.I.; Abou-Zeid, M.A. Diversity and virulence dynamics within Puccinia triticina populations in Egypt. J. Plant. Prot. Path., Mansoura Univ. 2018, 9, 735–745. [Google Scholar] [CrossRef]
- Stepień, L.; Golka, L.; Chelkowski, J. Leaf rust resistance genes of wheat: Identification in cultivars and resistance sources. J. Appl. Genet. 2003, 44, 139–149. [Google Scholar]
- Li, Z.F.; Xia, X.C.; He, Z.H.; Li, X.; Zhang, L.J.; Wang, H.Y.; Meng, Q.F.; Yang, W.X.; Li, G.Q.; Liu, D.Q. Seedling and slow rusting resistance to leaf rust in Chinese wheat cultivars. Plant. Dis. 2010, 94, 45–53. [Google Scholar] [CrossRef] [PubMed]
- German, S.E.; Kolmer, J.A. Leaf rust resistance in selected late maturity, common wheat cultivars from Uruguay. Euphytica 2014, 195, 57–67. [Google Scholar] [CrossRef]
- Boroujeni, F.R.; Arzani, A.; Afshari, F.; Torabi, M. Postulation of leaf rust resistance genes in Iranian wheat cultivars and breeding lines. Can. J. Plant. Pathol. 2011, 33, 550–558. [Google Scholar] [CrossRef]
- McIntosh, R.A.; Friebe, B.; Jiang, J.; Gill, B.S. Cytogenetical studies in wheat XVI. Chromosome location of a new gene for resistance to leaf rust in a Japanese wheat-rye translocation line. Euphytica 1995, 82, 141–147. [Google Scholar] [CrossRef]
- Herrera-Foessel, S.A.; Singh, R.P.; Huerta-Espino, J.; Rosewarne, G.M.; Periyannan, S.K.; Viccars, L.; Calvo-Salazar, V.; Lan, C.L.; Lagudah, E.S. Lr68: Anew gene conferring slow rusting resistance to leaf rust in wheat. Theor. Appl. Genet. 2012, 124, 1475–1486. [Google Scholar] [CrossRef] [PubMed]
- McDonald, B.A.; Linde, C. The population genetics of plant pathogens and breeding strategies for durable resistance. Euphytica 2002, 124, 163–180. [Google Scholar] [CrossRef]
- Kolmer, J.A.; Long, D.L.; Huges, M.E. Physiological specialization of Puccinia triticina on wheat in the united states in 2003. Plant. Dis. 2005, 89, 1201–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Genotype | Pedigree |
|---|---|---|
| 1 | Misr-1 | OASIS/SKAUZ//4*BCN/3/2*PASTOR.CMSSOYO1881T-050M-030Y-O3OM-30WGY-33M-0Y-0S. |
| 2 | Misr-2 | SKAUZ/BAV92. CMSS96M0361S-1M-010SY-010M-010SY-8M -0Y-0S. |
| 3 | Misr-3 | ATTILA*2/PBW65*2//KACHU CMSS06Y00582T-099TOPM-099Y-099ZTM-099Y-099M-10WGY-0B-0EGY |
| 4 | Misr-4 | NS-732/HER/3/PRL/SARA//TSI/VEE#5/FRET2/5/WHEAR/SOKOLL |
| 5 | Giza-139 | HINDI90/KENYA256G. |
| 6 | Giza-168 | MIL/BUC//Seri CM93046-8M-0Y-0M-2Y-0B |
| 7 | Giza-171 | Sakha 93/Gemmeiza 9 S.6-1GZ-4GZ-1GZ-2GZ-0S |
| 8 | Sakha-61 | INIA/RL4220//7CYR”S”. CM15430-2S-2S-0S-0S. |
| 9 | Sakha-94 | OPATA/RAYON//KAUZ. CMBW90Y3280-0TOPM-3Y-010M-010M-010Y-10M-015Y-0Y-0AP-0S. |
| 10 | Sakha-95 | PASTOR//SITE/MO/3/CHEN/AEGILOPS SQUARROSA(TAUS)//BCN/4/WBLL1CMSA01Y00158S-040P0Y-040M-030ZTM-040SY-26M-0Y-0SY-0S |
| 11 | Gemmeiza-5 | VEE”S”/SWM6525. GM4017-1GM-6GM-3GM-0GM. |
| 12 | Gemmeiza-7 | CMH74A.630/SX//SER182/3/AGENT. GM4611-2GM-3GM-1GM -0GM. |
| 13 | Gemmeiza-9 | ALD”S”/HUAC”S”//CMH74A.630/SX. GM4583-5GM-1GM-0GM. |
| 14 | Gemmeiza-10 | MAYA74”S”/0N//160-147/3/BB/GLL/4/CHAT”S”/5/CROW”S”. GM5820-3GM-1GM-2GM-0GM. |
| 15 | Gemmeiza-12 | OTUS/3/SARA/THB//VEE. CCMSS97Y00227S-5Y-010M-010Y -010M-2Y-1M-0Y-0GM |
| 16 | Sids-12 | BUC//7C/ALD/5/MAYA74/ON//1160-147/3/BB/GLL/4/CHAT”S”/6/MAYA/VUL-4SD-1SD-1SD-0SD. |
| 17 | Sids-13 | KAUZ “S”//TSI/SNB”S”. ICW94-0375-4AP-2AP-030AP-0APS-3AP-0APS-050AP-0AP-0SD. |
| 18 | Sids-14 | SW8488*2/KUKUNACGSS01Y00081T-099M-099Y-099M-099B-9Y-0B-0SD. |
| 19 | Shandaweel-1 | SITE/MO/4/NAC/TH.AC//3*PVN/3/MIRLO/BUC CMSS93B00567S-72Y-010M-010Y-010M-3Y-0M-0THY-0SH |
| 20 | Lr34/Yr18 | TC*6/P158548(RL6058) |
| 21 | Lr37/Yr17 | TC*6/VPM (RL6081) |
| 22 | Lr46/Yr29 | Pavon 76 |
| 23 | Lr67/Yr46 | RL6077 |
| Gene | Marker | Sequence of Primers 5′–3′ | Fragment Size | Reference |
|---|---|---|---|---|
| Lr34/Yr18 | Cslv34 F | GTT GGT TAA GAC TGG TGA TGG | 220 | Lagudah et al. 2006 |
| Cslv34R | GTG TTG CGC AAG TTT GTG A | |||
| Lr37/Yr17 | Ventriup | AGGGGCTACTGACCAAGGCT | 285 | Helguera et al. 2003 |
| LN2 | TGCAGCTACAGCAGTATGTACACAAAA | |||
| Lr46/Yr29 | Xgwm259F | AGG GAA AAG ACA TCT TTT TTT TC | 105 | William et al. 2003 |
| Xgwm259R | CGA CCG ACT TCG GGT TC | |||
| Lr67/Yr46 | CFD71F | CAA TAA GTA GGC CGG GAC AA | 198 | Forrest et al. 2014 |
| CFD71R | TGT GCC AGT TGA GTT TGC TC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Omara, R.I.; Shahin, A.A.; Ahmed, S.M.; Mostafa, Y.S.; Alamri, S.A.; Hashem, M.; Elsharkawy, M.M. Wheat Resistance to Stripe and Leaf Rusts Conferred by Introgression of Slow Rusting Resistance Genes. J. Fungi 2021, 7, 622. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080622
Omara RI, Shahin AA, Ahmed SM, Mostafa YS, Alamri SA, Hashem M, Elsharkawy MM. Wheat Resistance to Stripe and Leaf Rusts Conferred by Introgression of Slow Rusting Resistance Genes. Journal of Fungi. 2021; 7(8):622. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080622
Chicago/Turabian StyleOmara, Reda Ibrahim, Atef Abdelfattah Shahin, Shaimaa Mahmoud Ahmed, Yasser Sabry Mostafa, Saad Abdulrahman Alamri, Mohamed Hashem, and Mohsen Mohamed Elsharkawy. 2021. "Wheat Resistance to Stripe and Leaf Rusts Conferred by Introgression of Slow Rusting Resistance Genes" Journal of Fungi 7, no. 8: 622. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7080622