
Sclerotinia sclerotiorum SsCut1 Modulates Virulence and Cutinase Activity
by
 , , , , and
, , , , and
Yingdi Gong
1,2, 
Yanping Fu
2,
Jiatao Xie
1,2,
Bo Li
1,2,
Tao Chen
1,2,
Yang Lin
2 ,
,
Weidong Chen
3,
Daohong Jiang
1,2 and
Jiasen Cheng
1,2,* 1
State Key Laboratory of Agricultural Microbiology, Huazhong Agricultural University, Wuhan 430070, China
2
The Provincial Key Lab of Plant Pathology of Hubei Province, College of Plant Science and Technology, Huazhong Agricultural University, Wuhan 430070, China
3
United States Department of Agriculture, Agricultural Research Service, Washington State University, Pullman, WA 99164, USA
*
Author to whom correspondence should be addressed.
J. Fungi 2022, 8(5), 526; https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050526
Submission received: 21 April 2022
/
Revised: 13 May 2022
/
Accepted: 16 May 2022
/
Published: 20 May 2022
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The plant cuticle is one of the protective layers of the external surface of plant tissues. Plants use the cuticle layer to reduce water loss and resist pathogen infection. Fungi release cell wall-degrading enzymes to destroy the epidermis of plants to achieve the purpose of infection. Sclerotinia sclerotiorum secretes a large amount of cutinase to disrupt the cuticle layer of plants during the infection process. In order to further understand the role of cutinase in the pathogenic process of S. sclerotiorum, the S. sclerotiorum cutinsae 1 (SsCut1) gene was cloned and analyzed. The protein SsCut1 contains the conserved cutinase domain and a fungal cellulose-binding domain. RT-qPCR results showed that the expression of SsCut1 was significantly upregulated during infection. Split-Marker recombination was utilized for the deletion of the SsCut1 gene, ΔSsCut1 mutants showed reduced cutinase activity and virulence, but the deletion of the SsCut1 gene had no effect on the growth rate, colony morphology, oxalic acid production, infection cushion formation and sclerotial development. Complementation with the wild-type SsCut1 allele restored the cutinase activity and virulence to the wild-type level. Interestingly, expression of SsCut1 in plants can trigger defense responses, but it also enhanced plant susceptibility to SsCut1 gene knock-out mutants. Taken together, our finding demonstrated that the SsCut1 gene promotes the virulence of S. sclerotiorum by enhancing its cutinase activity.
1. Introduction
Sclerotinia sclerotiorum is a necrotrophic pathogen with a wide range of hosts, causing diseases and resulting in huge economic losses in many important crops such as oilseed rape, soybean, sunflower, and tomato [1]. Studies have shown that the pathogenicity mechanisms of S. sclerotiorum are very complicated. Many factors play important roles in the process of S. sclerotiorum infection, such as different plant cell wall-degrading enzymes and oxalic acid [2,3,4,5,6]. In addition, secreted proteins also play key roles in the pathogenic process of S. sclerotiorum, including SsCaf1, SsSSVP1, SsCP1 and SsITL [7,8,9,10]. At present, however, our understanding of the pathogenesis of S. sclerotiorum is still insufficient, and further research on its pathogenesis contributes to the development of new control strategies for Sclerotinia diseases [11,12,13].
The plant cuticle covers the leaf epidermis of higher plants and is the main barrier that isolates plants from direct contact with the atmosphere. The plant cuticle is mainly composed of cutin and wax, which is a hydrophobic property [14,15,16,17,18,19]. For different plant species, the composition of cutin and wax is quite different [20], but the ability to synthesize hydrophobic surface layers is evolutionarily conserved [16,18,21]. The plant cuticle is the first line of defense against pathogens, because the cuticle covers the outermost layer of the plant epidermis. The plant cuticle can resist pathogen invasion and endow the corresponding microbial environment [19,22,23,24,25,26,27,28,29]. Interestingly, many studies have shown that the cuticle is not only a physical barrier of protection, but also participates in plant growth and development, immunity, and signal transduction [30,31,32,33,34].
The plant cuticle is an important component for plant–fungi interactions, so alterations to its structure and permeability might change immunity to pathogenic fungi [17,35]. Most plant pathogens produce cutinase to destroy the plant cuticle to facilitate their infection [36,37,38,39,40]. Cutinase (EC 3.1.1.74) is a serine esterase belonging to the α/β hydrolase superfamily, capable of breaking down cutin polyesters. Most cutinases contain a conserved GYSQG domain and the similar DxVCxG [ST]-[LIVMF] (3)-x (3) H motif. They have the classic Ser–His–Asp catalytic triad and can hydrolyze a variety of substrates, including low-molecular-weight soluble esters as well as short- and long-chain triacylglycerols. Cutinase can also catalyze esterification and transesterification [37,41].
Although cutinases share similarities in conserved domains and amino acid sequences, cutinases from fungi of different lifestyles vary in properties such as molecular weight, optimum temperature and pH optimum for action, substrate specificity and thermostability [42]. Cutinase is able to degrade cutin polymers, thereby destroying the physical properties of the plant epidermis [29]. So cutinase plays important roles during the infection process of many plant pathogenic fungi. For example, disruption of cutinase gene Pbc1 can lead to the loss of virulence in Pyrenopeziza brassicae [43]. In the corn leaf pathogen Curvularia lunata, deletion of Clcut7 also reduced the virulence and cutin-decomposing ability [44]. The virulence of the cutinase gene CglCUT1 knock-out strain was also decreased in Colletotrichum gloeosporioides [45]. However, in some pathogens such as Nectria haematococca, Botrytis cinerea, Fusarium graminearum and Ustilaginoidea virens, the knock-out of specific cutinase genes showed no effect on the virulence [46,47,48,49], possibly due to redundant function of multiple cutinase genes. An obvious example is that the knock-out mutants of either AaCut3 or AaCut7 have a normal virulence phenotype in Alternaria alternata, while the virulence of the double-mutant strains of AaCut3 and AaCut7 are significantly reduced [50]. On the other hand, the enhancement of cutinase activity can increase the virulence of pathogenic fungi. The overexpression strain of MfCUT1 in Monilinia fructicola produced larger lesions on Prunus flower petals than the wild-type strain [51]. Interestingly, some studies have shown that cutinase can also activate plant immune response during infection. For instance, purified Verticillium dahliae protein VdCUT11 can induce cell death in tobacco, cotton and tomato plants and trigger a defense response [52]. The cutinase RcCUT1 of Rhizoctonia cerealis can also induce necrosis of plant cells, H2O2 accumulation and expression of defense-related genes [53]. These results indicate that the role of cutinase in the infection of pathogenic fungi is complicated, and it also participates in the interaction between host and pathogens.
S. sclerotiorum secretes a large number of hydrolytic enzymes when infecting plants, including cell wall-degrading enzymes, cutinase and proteases. It was predicted that there are eight genes encoding cutinase proteins in the S. sclerotiorum genome [54]. One of the cutinase genes, SsCUT, is highly expressed in the early stage of infection. Expression of SsCut in Nicotiana benthamiana can induce the death of host cells and activate the plant immune response [55,56]. However, the effect of SsCut on the phenotype of S. sclerotiorum was not involved in previous studies. Additionally, the functions of other cutinase genes have not been studied in S. sclerotiorum. In this study, the expression level, secretory activity and subcellular localization of SsCut1 were clarified. Most importantly, the SsCut1 gene was knocked out, and the cutinase activity and the virulence of the SsCut1 knock-out mutants decreased significantly, while the cutinase activity and virulence of the complementary strain all recovered to the level of wild-type strain. In addition, expression of SsCut1 in N. benthamiana plant can complement the lost virulence of the SsCut1 knock-out mutant. Our results suggest that SsCut1 plays important roles in the pathogenesis of S. sclerotiorum.
2. Materials and Methods
2.1. Strains and Culture Conditions
The S. sclerotiorum (Lib.) de Bary wild-type strain 1980 (ATCC 18683) was obtained from diseased bean plants in Scottsbluff, NE [57]. In this study. Fungal strains were cultured on potato-dextrose agar (PDA) plates at 20 °C. The SsCut1 gene knock-out mutants and complement mutants were cultured on PDA plates amended with hygromycin B or G418 at 100 µg/mL (Sigma-Aldrich, St. Louis, MO, USA).
Escherichia coli DH5α and Top10 (Transgen Biotech, Wuhan, China) for plasmid proliferation was cultured in Luria–Bertani (LB: 1% Tryptone, 0.5% Yeast extract, 0.5% NaCl and 1% agar, pH = 7) medium with appropriate antibiotics (Kanamycin or Ampicillin). The LB medium with appropriate antibiotics (Kanamycin and Rifampicin) was used to culture Agrobacterium tumefaciens EHA-105 and GV3101, which was used in fungal transformation and protein expression.
2.2. Bioinformatics Analysis
Sequences of SsCut1 were retrieved from the NCBI GenBank database. The cutinase signal peptide sequence was predicted by using SIGNAIP 5.0 tools (http://www.cbs.dtu.dk/services/SignalP/) (accessed on 17 May 2022, DTU, Copenhagen, Denmark). The structural domains of cutinase were predicted by NCBI Conserved Domain Search Tools (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/Structure/cdd/wrpsb.cgi) (accessed on 17 May 2022, Bethesda, MA, USA). The cutinase promoter was predicted by using Promoter 2.0 Prediction Server (http://www.cbs.dtu.dk/services/Promoter/) (accessed on 17 May 2022, DTU Bioinformatics, Copenhagen, Denmark). The cutinase transmembrane domains were predicted by using TMHMM Server, v. 2.0 (http://www.cbs.dtu.dk/services/TMHMM/) (accessed on 17 May 2022, DTU Bioinformatics, Copenhagen, Denmark). Other cutinase sequences were also retrieved from the NCBI GenBank database. Amino acid sequence alignments were generated via the ES Pript 3.0 Tools (https://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi) (accessed on 17 May 2022). The phylogenetic tree was constructed with MEGA 6.0 [58] using the maximum-likelihood method.
2.3. DNA/RNA Manipulation and RT-qPCR
Genomic DNA was isolated from mycelium by using the CTAB method [59]. Total RNA of fungi and plants were isolated with RNAiso Plus regent (Takara, Dalian, China) according to the manufacturer’s protocols. To detect the expression pattern of cutinase gene in infected tissues, the hyphae of S. sclerotiorum wild-type strain were inoculated on Arabidopsis thaliana plant leaves. The hyphae were collected at 0, 1, 3, 6, 9, 12 and 24 h after inoculation. After RNA extraction, the first-strand cDNA was synthesized by the Easy Script One-Step gDNA Removal and cDNA Synthesis SuperMix (TransGen Biotech, Beijing, China). Additionally, the gene expression was detected by qPCR analysis [60]. The β-tubulin gene in S. sclerotiorum, the UBQ5 gene in A. thaliana, and the elongation factor EF1α gene in N. benthamiana were used as internal controls, respectively. The real-time RT-qPCR analysis was repeated at least three times, with three biological replicates for each repeat.
2.4. The Yeast Secretion Trap Screen Assay
The secretion activity of SsCut1 was analyzed by using the yeast secretion trap assay [61,62]. In this study, the predicted signal peptide fragment of the SsCut1 gene was fused to the N-terminal of secretion-defective invertase gene (suc2) in the vector pSUC2 and Then transformed into the yeast competent strain YTK12. The candidate yeast transformants were cultured on YPDA medium, and selected on CMD-W (Takara, Dalian, China) medium and YPRAA (2% raffinose) medium (Macklin, Shanghai, China). TTC (2, 3, 5-Triphenyte-trazoliumchloride) was used to detect the activity of the secreted sucrase of the candidate yeast transformants. The candidate yeast transformed strain was cultured on sucrose medium, then collected by centrifugation, added with a final concentration of 0.1% TTC reagent, incubated at 35 °C for 35 min, and placed at room temperature for 5 min to observe the color change in the test tube. A positive reaction changes from colorless to dark red. The SPs of Avr1b and YTK12-pUSC2 were used as the positive and negative controls, respectively.
2.5. Gene Deletion and Complementation
The cutinase gene SsCut1 was disrupted by using the split marker system [63]. The strategy for the deletion of SsCut1 is illustrated in Supporting Information Figure S2A. To generate SsCut1-deletion mutants, the 5′ and 3′ flanking fragments of the ORF of SsCut1 were amplified from S. sclerotiorum wild-type strain 1980 genomic DNA, then cloned into the pMD19-T plasmid using the T/A clone ligation kit (Takara, Shiga, Japan). Two flanking sequences with a truncated hygromycin-resistant gene sequence were named 5′-HY and YG-3′, respectively. The 5′-HY and YG-3′ fragments were amplified and purified. The purified fragments of 5′-HY and YG-3′ were mixed in equal molarity, and used for protoplast transformation [64]. ∆SsCut1 mutants were selected on plates containing hygromycin and confirmed by PCR with genomic DNA as template. Homokaryotic ∆SsCut1 mutants were obtained by single ascospore isolation. The homokaryotic state of the ∆SsCut1 mutants was confirmed by Southern blot. The genomic DNA was extracted from mycelium and digested with SpeI. Southern blot analysis with SsCut1 as a probe was detected by using Amersham AlkPhos Direct Labelling and Detection Systems (GE Healthcare, Piscataway, NJ, USA).
For the complementation of SsCut1, the binary vector pCENTS-SsCut1 was constructed. The complementary fragment was amplified from the S. sclerotiorum genomic DNA, including the open reading frame of SsCut1 with an approximately 1200 bp native promoter region and a 200 bp terminator region. The pCENTS was cut by XhoI and SpeI to remove the EF-1α promoter and then ligated with the complementary fragment. The complementary vector was transformed into Agrobacterium strain EHA105. Agrobacterium tumefaciens-mediated transformation (ATMT) of S. sclerotiorum was performed as described [65]. Cultivate the Agrobacterium broth and the mycelium blocks of cutinase knock-out mutants in the dark condition for three days. Then, cover with the PDA medium containing hygromycin B and G418 at 100 µg/mL resistance. The DNA of candidate complementary transformants were extracted by using the CTAB method for validation.
2.6. Plant Growth and Pathogenicity Assays
The A. thaliana and N. benthamiana were grown at 20 °C in growth chambers or glasshouse under long day (16 h: 8 h, light: dark) conditions. Pathogenicity assays of S. sclerotiorum were tested with mycelia plugs (diameter = 4.0 mm) from the margins of actively growing colonies on PDA medium. Inoculated tissues were incubated at 21 °C for 48 h before image acquisition and measurement of lesion area. The treatment was replicated 10 times and the experiments were repeated three times.
2.7. Generation of Plant Expression Plasmids
In order to express SsCut1 in N. benthamiana, a plasmid with the backbone of pCNF3 and pCNG were used [9]. The recombinant plasmid pCNF3-SsCut1 and pCNG-SsCut1 were transformed into Agrobacterium strains GV3101 for expression. The protein of N. benthamiana were extracted for Western blot at 48 h post-infiltration. The leaf specimens of N. benthamiana were prepared to observe the protein subcellular localization by using the confocal microscope (Olympics).
2.8. Cutinase Activity Assay
The ELISA Reagent test kit (Sinobestbio, Shanghai, China) was used to determine the cutinase activity of fungal hypha samples. The HRP-labeled detection antibody was added to the coated microwells pre-coated with cutinase antibody, incubated and washed thoroughly. The color is developed with the substrate TMB. TMB is converted into a blue color under the catalysis of peroxidase, and converted into the final yellow under the action of acid. The intensity of the color is positively correlated with the cutinase activity in the sample. The absorbance (OD value) at a wavelength of 450 nm was measured with a microplate reader to calculate the cutinase activity in the sample.
3. Results
3.1. SsCut1 Contains Cutinase Domain and Fungal Cellulose-Binding Domain
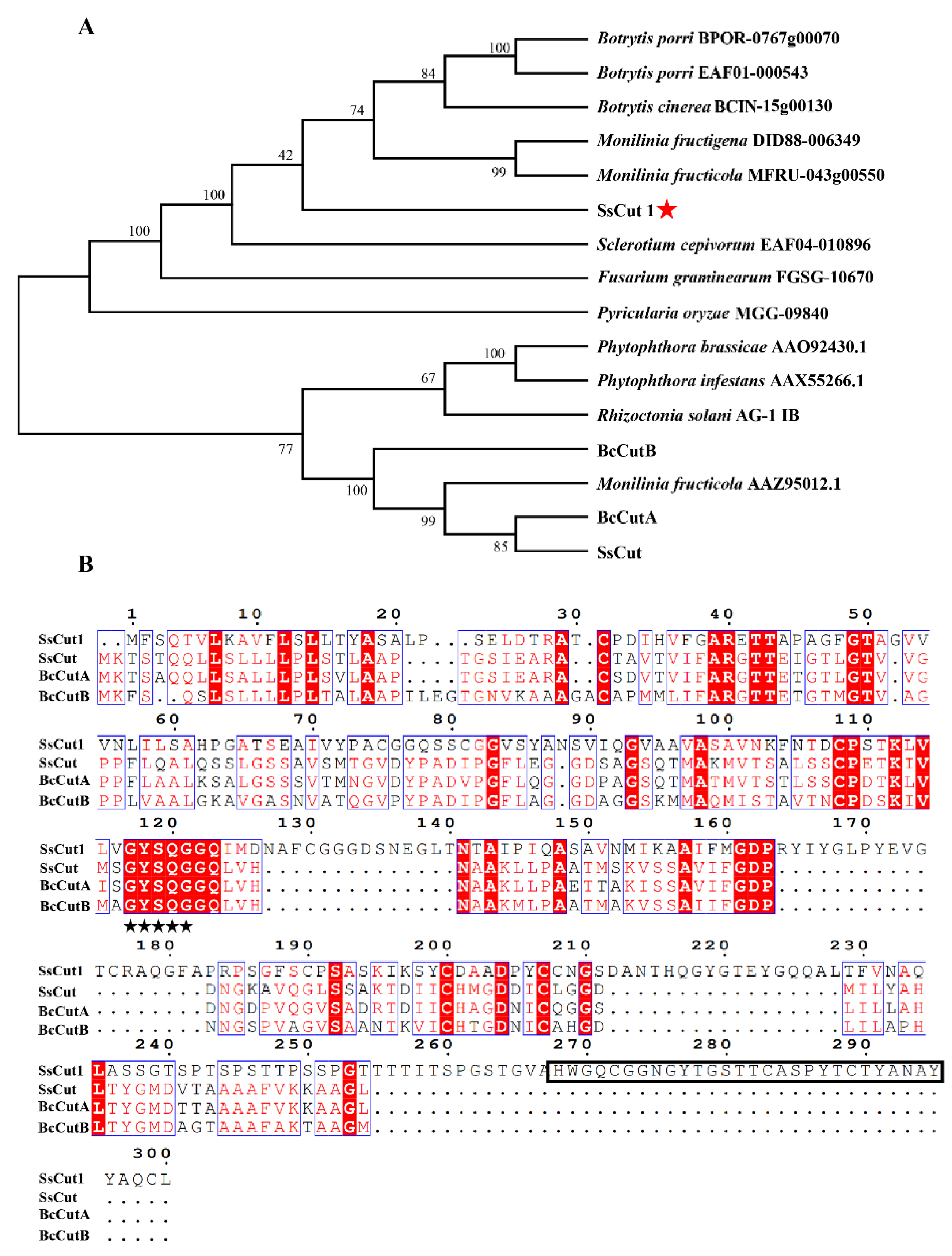
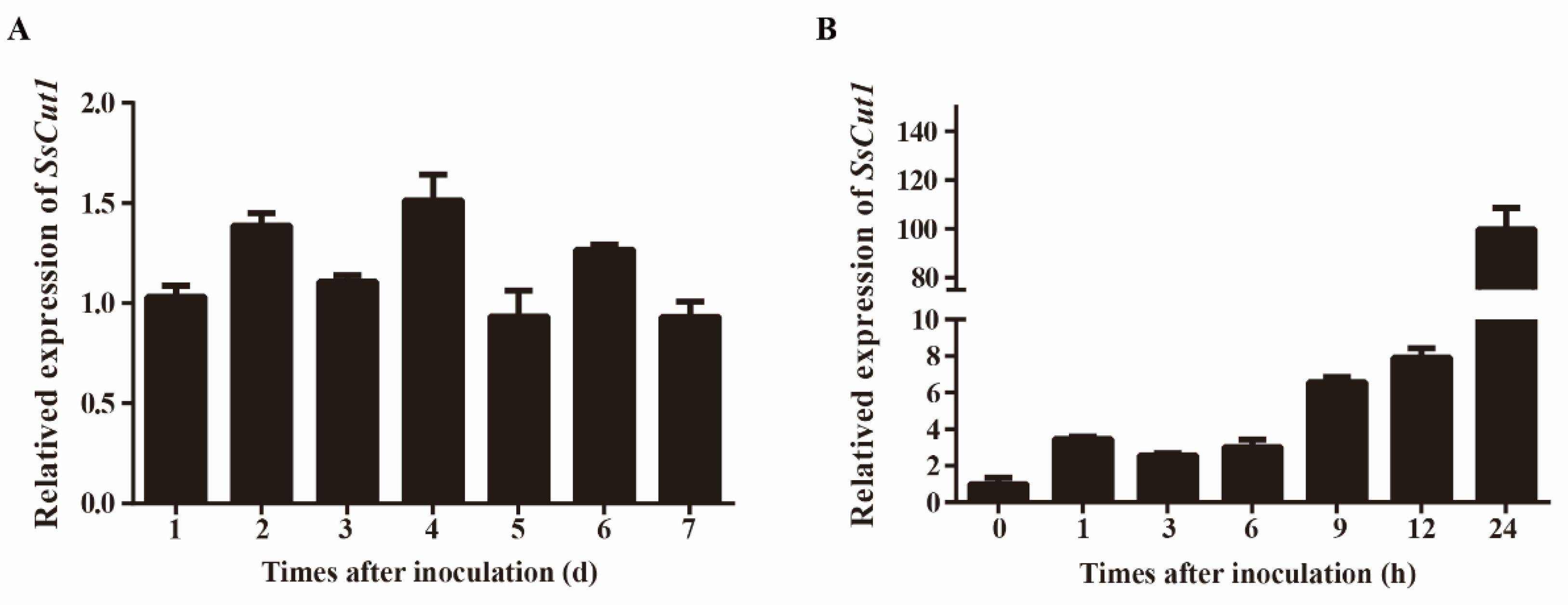
Sequence analysis showed that there are eight genes encoding cutinase proteins in the genome of S. sclerotiorum (Supplementary Figure S1). SS1G_08104 is one of the cutinase-encoding genes, which was named SsCut1 in this study. SsCut1 is 1077 bp in length and contains three exons and two introns. Its open reading frame encodes a 300 aa protein with a predicted N-terminal SP. SsCut1 possesses fourteen cysteine residues. Phylogenetic analysis showed that the homologous protein of SsCut1 was widely found in B. cinerea and other pathogenic fungi, but it did not cluster with the reported cutinase SsCut, BcCutA and BcCutB (Figure 1A). Protein structure analysis showed that the SsCut1 had conserved GYSQG catalytic site; however, it differed from SsCut, BcCutA and BcCutB in in certain features, consistent with the results of cluster analysis. SsCut1 contains a fungal cellulose-binding (CBM1) domain which was absent in SsCut, BcCutA and BcCutB (Figure 1B). The expression pattern of SsCut1 was detected by RT-qPCR. The results revealed that the expression of SsCut1 was relatively stable during the growth and development stage of S. sclerotiorum (Figure 2A). However, during the infection stage, the transcript level of SsCut1 was upregulated and increased by more than 100 fold at 24 h post-inoculation (hpi) on A. thaliana (Figure 2B), suggesting that SsCut1 may be involved in infection by S. sclerotiorum.
3.2. SsCut1 Is a Secreted Protein and Localized in the Plant Cell Wall
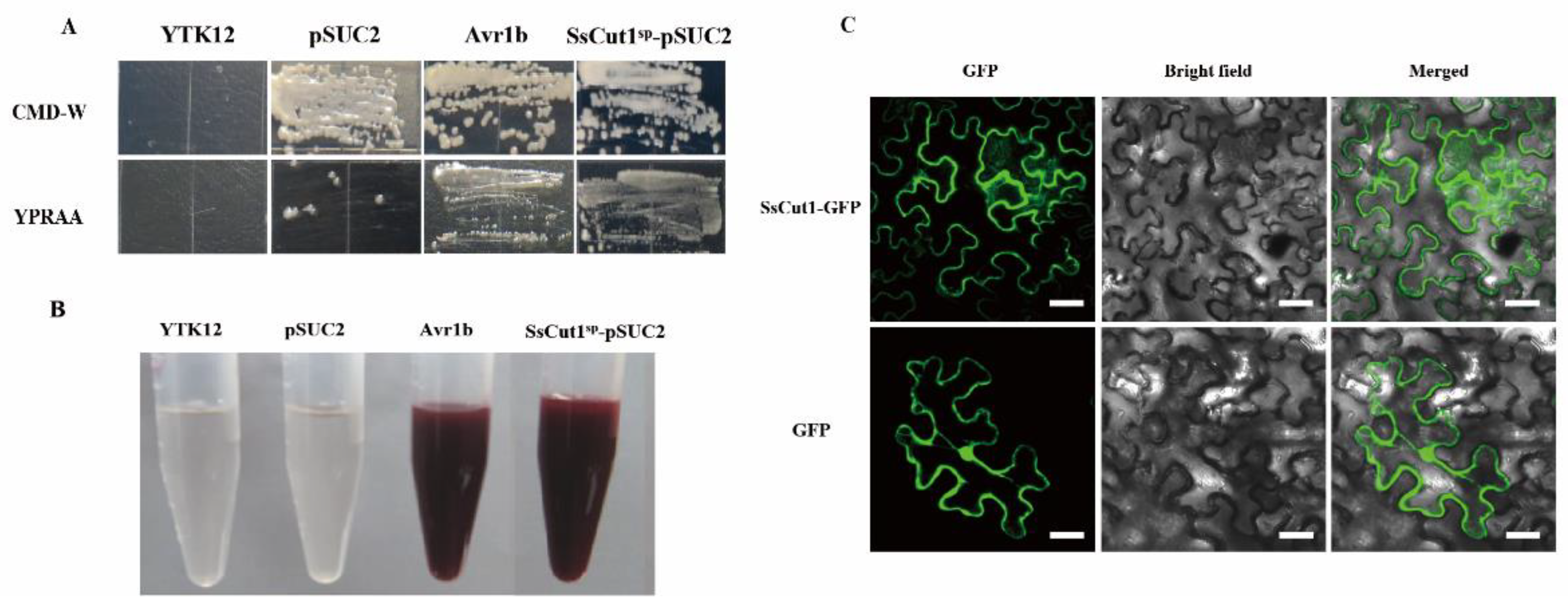
To investigate whether SsCut1 is normally secreted by S. sclerotiorum, the function of SsCut1 signal peptide was verified by the yeast secretion trap system [61,62]. The Avr1b was used as positive control and the YTK12 empty strain and YTK12 with pSUC2 vector were used as negative controls. The invertase mutant yeast strain YTK12 of pSUC2-Avr1b and pSUC2-SsCut1sp could regularly grow on CMD-W and YPRAA media (Figure 3A). When treating with 2, 3, 5 triphenyltetrazolium chloride (TTC), the mutant strains of pSUC2-Avr1b and pSUC2-SsCut1sp generated the insoluble red-colored triphenylformazan, but the yeast strains YTK12 and pSUC2 remained colorless after treatment (Figure 3B). These results indicated that the signal peptide of SsCut1 was functional in the yeast secretion trap system, and SsCut1 might be a secreted protein.
SsCut1 contains a fungal cellulose-binding domain of CBM1 superfamily at the 268–296 amino acid positions (Figure 1B). This domain has high specific affinity to cellulosics [66]. We speculated that SsCut1 could localize in the plant cell wall. In order to clarify the localization of SsCut1 in plant cells, we performed a protein cellular localization assay in N. benthamiana. Fluorescence observation results showed that SsCut1 can be localized in the cell wall when expressed in plant cells (Figure 3C).
3.3. SsCut1 Is Important for Virulence of S. sclerotiorum
The biological function of SsCut1 in S. sclerotiorum was investigated by generating SsCut1-deletion mutants, using a homologous recombination strategy (Supplementary Figure S2A). Homokaryotic ΔSsCut1 mutants were obtained by single ascospore isolation. Three SsCut1-deletion mutants (ΔSsCut1-1, ΔSsCut1-3 and ΔSsCut1-7) were obtained, and confirmed by PCR and Southern blot analysis. The results showed that the targeted gene was replaced by a single copy marker gene in mutants ΔSsCut1-1 and ΔSsCut1-3 (Supplementary Figure S2B,C). Two complementary transformants, ΔSsCut1-3-C8 and ΔssCut1-3-C12, were generated by introducing wild-type allele to ΔSsCut1-3 mutants using the ATMT method (Supplementary Figure S2D).
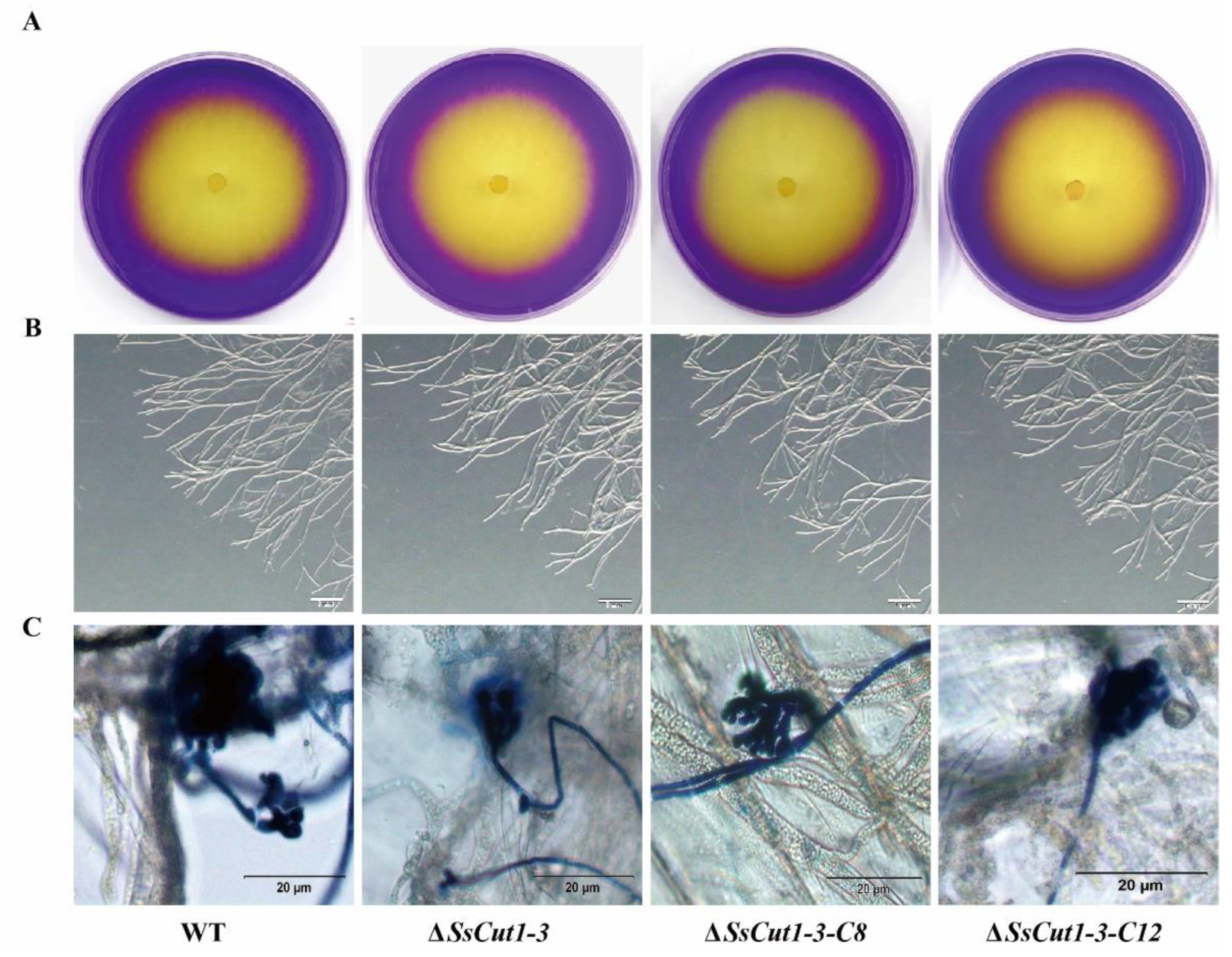
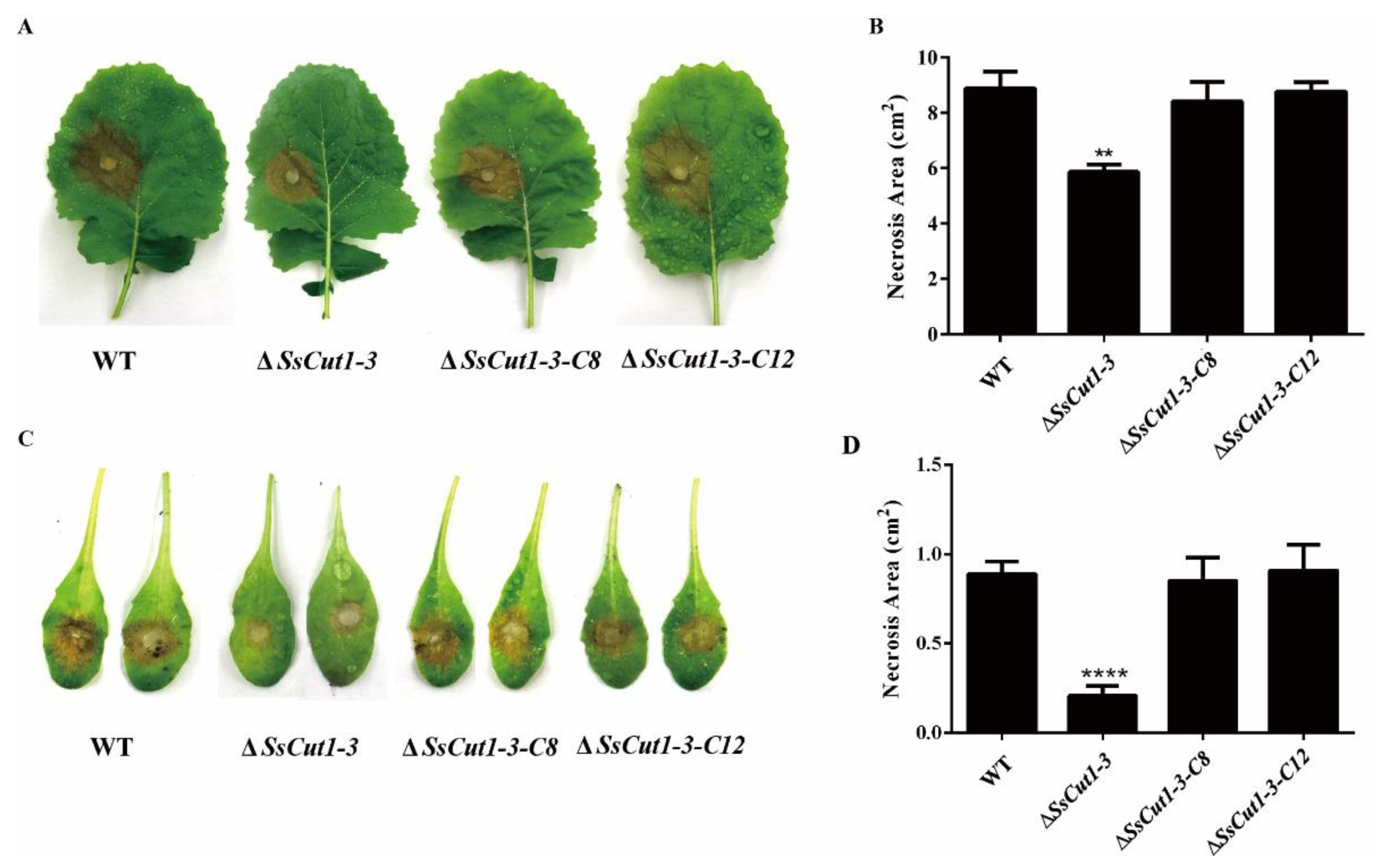
Deletion of Sscut1 had no effect on the saprophytic growth and life cycle of S. sclerotiorum. ΔSsCut1 mutants showed no significant differences from the wild-type strain in colony morphology, oxalic acid production, infection cushion formation, sclerotial development and growth rate (Figure 4 and Figure S3). To determine the function of the SsCut1 in the virulence of S. sclerotiorum, the wild-type strain (1980), SsCut1 knock-out mutants, and the complementary transformants were inoculated on the leaves of oilseed rape and A. thaliana plants. Compared with the wild-type strain, both knock-out mutants caused smaller lesions on the oilseed rape and A. thaliana leaves. Complementation of SsCut1 in the knock-out strain restored virulence to the level of the wild-type strain (Figure 5 and Figure S4). These results indicated that SsCut1 is not required for completion of life cycles, but required for full virulence of S. sclerotiorum.
3.4. Deletion of SsCut1 Affects the Cutinase Activity of S. sclerotiorum
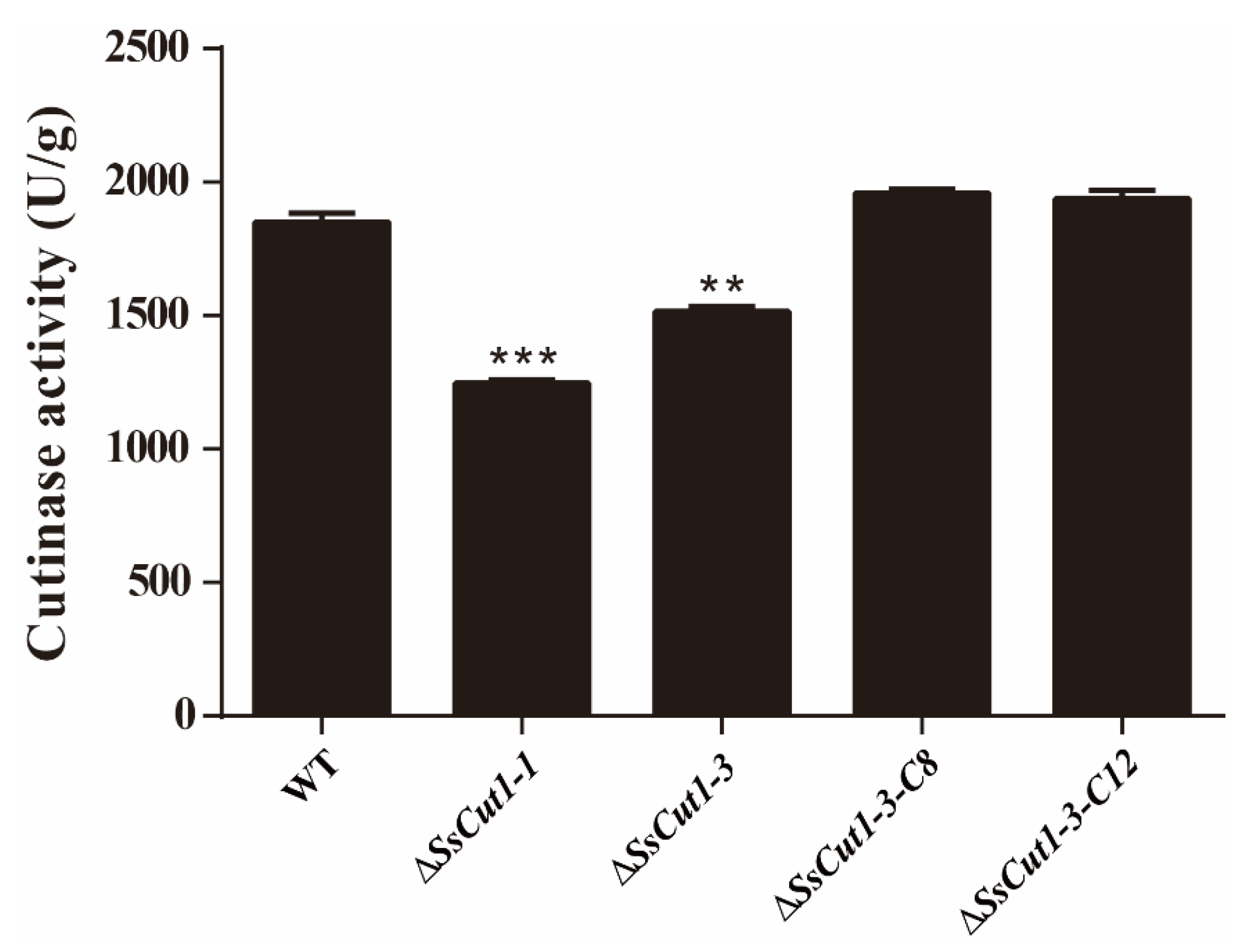
To investigate if SsCut1 is associated with the cutinase activity in S. sclerotiorum, wild-type strain and different mutants of SsCut1 were cultured on the PDA medium for 36 h. Then, the hyphae were collected for determination of cutinase activity. Results showed that the cutinase activity of SsCut1 knock-out transformants were significantly decreased compared with wild-type strain, while the cutinase activity of SsCut1 complementary transformants were recovered to the level of wild-type strain (Figure 6), indicating SsCut1 does encode a cutinase in S. sclerotiorum. The decrease in virulence of SsCut1 knock-out transformants should be related to the decrease in cutinase activity.
3.5. The Expression of SsCut1 in Plants Triggers Defense Responses
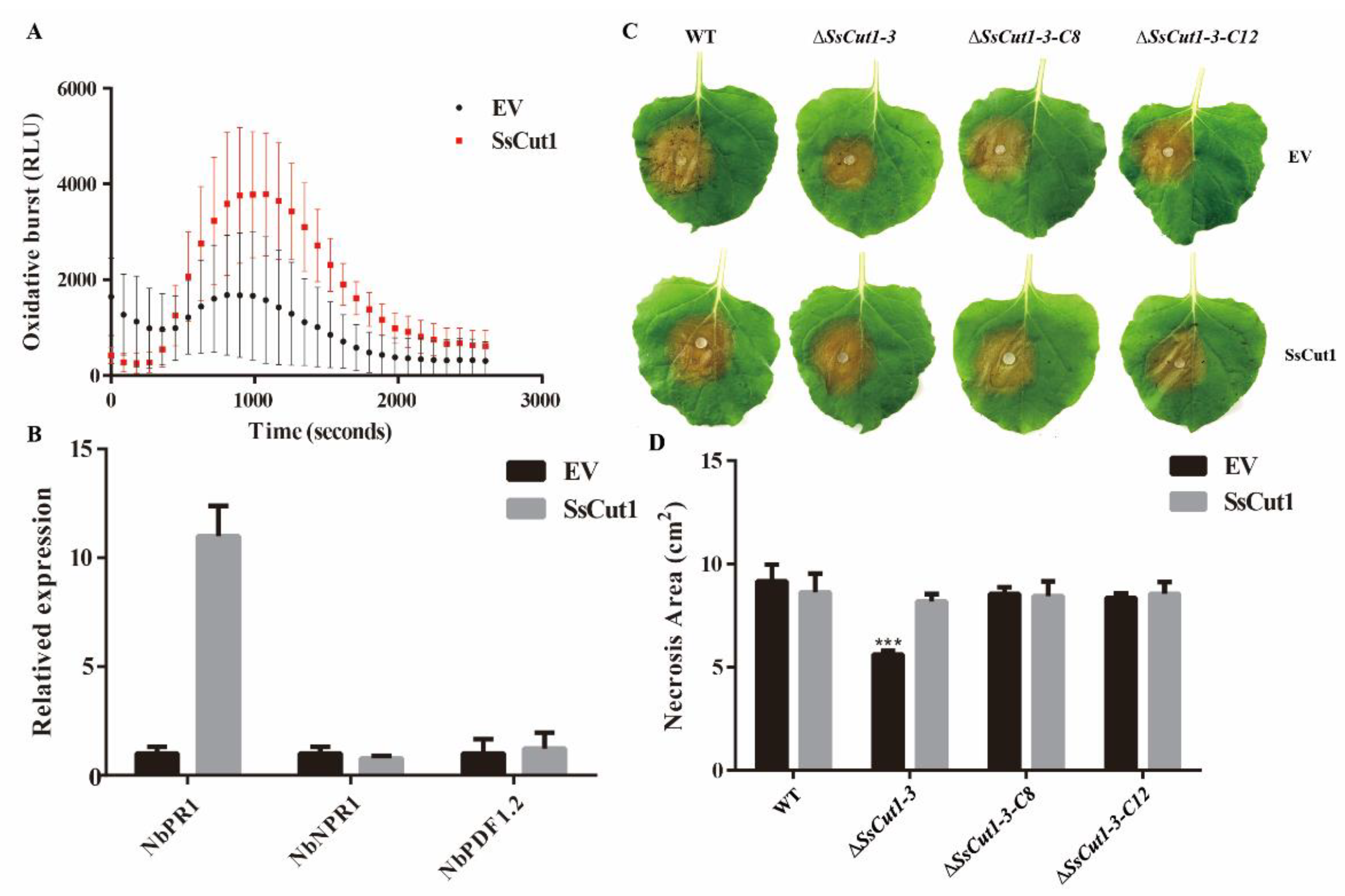
To evaluate whether SsCut1 participates in Sclerotinia–plant interaction, transient expression of SsCut1 was carried out in N. benthamiana. ROS burst was observed when we transiently expressed SsCut1-flag in N. benthamiana compared with that expressing mock vector (Figure 7A). Furthermore, the expression levels of NbPR1 were also significantly upregulated when SsCut1-flag was expressed (Figure 7B). These results suggested that the expression of SsCut1 in N. benthamiana can trigger plant defense responses. Interestingly, the lesion size of ΔSsCut1-3 mutant on SsCut1-expressing plants was similar to that of the wild-type strain level (Figure 7C, D), suggesting that SsCut1 expressed in plants can complement the virulence defect of the SsCut1-deletion mutant. It is another line of evidence that SsCut1 is crucial to the virulence of S. sclerotiorum and plays an important role in the Sclerotinia–plant interaction.
4. Discussion
Cutinase is important for the virulence of many fungal pathogens. The S. sclerotiorum genome contains eight cutinase genes [54], but only SsCut has been studied [55,56]. However, SsCut was only expressed in N. benthamiana to observe the plant defense response, but the effect of SsCut on S. sclerotiorum phenotype has not been studied. SsCut1 encodes a cutinase protein in S. sclerotiorum and shows increased expression during the early stage of S. sclerotiorum infection (Figure 2), indicated that SsCut1 plays an important role in the early stage of infection. Therefore, in this study, we explored the effects of SsCut1 on virulence and cutinase activity of S. sclerotiorum.
Most cutinase protein sequences contain a highly conserved GYSQG motif, which is crucial for maintaining the serine activity and function of cutinase [67]. In S. sclerotiorum, all the eight cutinase genes contain the conserved GYSQG domain, but the intron and exon structures of the eight cutinase genes are different (Supplementary Figure S1). It is suggested that the cutinase gene in S. sclerotiorum genome not only keeps the core GYSQG structure, but also increases the universality and adaptability of cutinase, which is beneficial to the expansion and stability of the host range of pathogenic fungus. The amino acid sequence alignment also shows that although the protein sequences of SsCut1, SsCut, BcCutA and BcCutB are highly conserved in key domains, there are some regions with unknown functions at the C-terminal of SsCut1 (Figure 1), and it is also unknown whether these regions endow new functions to SsCut1.
Plant epidermis layers, mainly composed of cutin, wax, and cellulose, are the first barrier for fungi to infect plants [68,69]. Plant pathogenic fungi, especially necrotrophic fungi, can destroy the integrity of plant cell walls through combined actions of a large number of cell wall-degrading enzymes to facilitate invasion. During this process, cutinase plays key roles and is usually released at the early stage of infection to degrade the plant cuticle, which is the first barrier of plant [70]. Therefore, knocking out cutinase or destroying its activity often has a great influence on the virulence of plant pathogenic fungi. For example, deletion of genes encoding cutinase reduced virulence in Colletotrichum truncatum, C. gloeosporioides, Botryosphaeria dothidea and Arthrinium phaeospermum [40,45,71,72]. Using enzyme inhibitors to inhibit cutinase activity can also prevent Fusarium solani pisi from infecting plants [73]. We also found that SsCut1 knock-out transformants have no difference in growth rate, colony morphology, infection cushions formation, sclerotia formation, and oxalic acid production compared with wild-type strain (Figure 4 and Figure S3). However, the virulence and cutinase activity of the SsCut1 knock-out transformants were significantly reduced, while there was no obvious difference between complementary transformants and wild-type strain (Figure 5 and Figure 6). Considering the high-level expression of SsCut1 in the S. sclerotiorum infection process, our results suggest that the SsCut1 has cutinase activity and is necessary for full virulence of S. sclerotiorum. There are reports that knocking out genes encoding cutinase has no obvious effect on the virulence of some plant pathogenic fungi [46,47,48,49], due to redundancy of gene functions for existence of multiple genes encoding cutinase in the genome. There are also eight cutinase genes in S. sclerotiorum [54], which is why the virulence of S. sclerotiorum only decreased after SsCut1 knock-out, but not completely lost.
During the interaction between plants and pathogenic fungi, cutinase can not only degrade the cuticle of plants, but also be recognized by plants as a PAMP (Pathogen-associated molecular patterns), thus activating the immune response [40,52,53,56,74]. For example, overexpression of the cutinase gene CUTE (cutinase-expressing) in Arabidopsis can improve the resistance of plants to B. cinerea [35]. Overexpression of a cutinase gene from B. dothidea (Bdo_10846) in N. benthamiana induced ROS burst and callose deposition, and also enhanced N. benthamiana resistance to B. cinerea [40]. The SsCut protein of S. sclerotiorum can also cause cell death in soybean and rapeseed, and induce resistance to S. sclerotiorum and B. cinerea in these plants [56]. In this study, we found that the expression of SsCut1 in N. benthamiana induced ROS burst, and upregulated expression of host pathogenesis-related genes (Figure 7A,B). However, when the wild-type strain 1980 was inoculated, the disease lesions on the plants expressing SsCut1 did not increase compared with the empty vector-expression plants, indicating that the resistance of these plants to S. sclerotiorum did not change significantly (Figure 7C,D). This may be due to the strong virulence of S. sclerotiorum or the existence of multiple cutinase in the genome of S. sclerotiorum. SsCut1 and other cutinase in S. sclerotiorum may also be recognized by plants during infection, thus inducing plant immune response. For example, the expression of SsCut also can trigger plant immune response [56], so the immune response induced by SsCut1 expression in plants may not be enough to affect the virulence of S. sclerotiorum. Interestingly, the virulence of the SsCut1 knock-out mutant on SsCut1-expressing plants recovered to the level of wild-type strain 1980, indicating that SsCut1 is important to the virulence of S. sclerotiorum and plays an important role in plants during infection.
5. Conclusions
In summary, our study revealed that SsCut1 is important for the cutinase activity and full virulence of S. sclerotiorum, and plays important roles in triggering plant defense responses. Here, we identified a conserved cutinase gene, SsCut1 from S. sclerotiorum. SsCut1 was found to be expressed highly in the early stage of S. sclerotiorum infection. SsCut1 had secretory activity and was mainly located in the plant cell wall. Knocking out SsCut1 reduced the virulence and cutinase activity of S. sclerotiorum, but had no effect on the growth rate, colony morphology, oxalic acid production, infection cushion formation and sclerotial development. Expression of SsCut1 in plants can trigger defense responses, such as induced ROS burst and upregulate the expression of pathogenesis-related genes. However, it also enhances plant susceptibility to SsCut1 gene knock-out mutants. These findings improve our understanding of the roles of cutinase gene during the interaction of S. sclerotiorum with its host plants, and also offer clues to further reveal the mechanisms of these genes in infection.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/jof8050526/s1, Figure S1: Exon and intron structure display of cutinase family genes of S. sclerotiorum. Exons and introns are indicated by red boxes and lines, respectively. The names of the S. sclerotiorum cutinase genes and intron-exon structures are indicated at the left and right sides, respectively. The number in the figures indicates the length of each intron and exon. Figure S2: Analysis of SsCut1 mutants. (A) Schematic diagram of the SsCut1 gene knock-out strategy. The different colored parts show the HptII cassette (white color parts), SsCut1 (black box parts) and flanked sequences (grey box parts) (B) PCR validation of the SsCut1 knock-out strains. Partial HptII fragment (lanes 3, 7, 11, 15); the upstream of SsCut1 overlapped Ptrp (lanes 1, 5, 9, 13); the downstream of SsCut1 overlapped trpC (lanes 2, 6, 10, 14); the full-length SsCut1 (lanes 4, 8, 12, 16). Lane M, DL5000 marker (Takara, Dalian, China). (C) Southern blot analysis of the SpeI-digested genomic DNAs from wild-type strain, ΔSsCut1-1, ΔSsCut1-3 and ΔSsCut1-7 strains. Probe was labelled with alkaline phosphatase. Lane M: DNA marker λ-HindIII DNA Ladder. (Takara, Dalian, China) (D) PCR validation of the SsCut1 complementary strains. The full-length SsCut1 (lanes 1, 4, 7, 10); partial HptII fragment (lanes 2, 5, 8, 11); partial G418 fragment (lanes 3, 6, 9, 12). Lane M, DL2000 marker (Takara, Dalian, China). Figure S3: Colony morphology of WT, ΔSsCut1-3, ΔSsCut1-3-C8 and Δ SsCut1-3-C12 strains. (A) Growth of wild-type strain, ΔSsCut1-3, ΔSsCut1-3-C8 and ΔSsCut1-3-C12 strains on PDA medium at 20 °C in complete darkness. Photographs were taken at 36 h post-inoculation (hpi). (B) Sclerotial primordium of wild-type strain, ΔSsCut1-3, ΔSsCut1-3-C8 and ΔSsCut1-3-C12 strains on PDA medium at 20 °C in complete darkness. Photographs were captured at 3 days post-inoculation (dpi). (C) Sclerotial formation of wild-type strain ΔSsCut1-3, ΔSsCut1-3-C8 and ΔSsCut1-3-C12 strains on PDA medium at 20 °C in complete darkness. Photographs were taken at 7 days post-inoculation (dpi). (D) Growth rate of all the strains growing on a PDA plate at 20 °C in complete darkness. (E) Comparation of sclerotia weight of strains growing on a PDA plate at 20 °C for 7 days in complete darkness. (F) Sclerotia number per plate of strains growing on a PDA plate at 20 °C for 7 days in complete darkness. Three independent replications were performed for each treatment. Bars indicate ± SE. Statistical differentiation was evaluated by t-test. Different letters on a graph indicate significant differences, p < 0.01. Figure S4: All the knock-out mutants of SsCut1 showing reduced virulence on detached oilseed rape leaves. (A) Lesions formation on oilseed rapeseed leaves inoculated with wild-type strain and SsCut1 transformants, the photographs were taken at 24, 36, 48 h post-inoculation (hpi), respectively. (B) Statistical results of lesion area on oilseed rape leaves. Bars indicate ± SE. Statistical significance is indicated in the graph (one-way ANOVA): * p < 0.05, ** p < 0.01.
Author Contributions
Y.G. and J.C. designed the experiments. Y.G. performed the experiments and data analyses. D.J., Y.F., J.C., J.X., B.L., T.C. and Y.L. contributed reagents/materials/analysis tools. Y.G. wrote the original draft, J.C. and W.C. revised and editing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Nature Science Foundation of China (32130087, 32172368) and the earmarked fund of China Agriculture Research System (CARS-13).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boland, G.J.; Hall, R. Index of plant hosts of Sclerotinia sclerotiorum. Can. J. Plant. Pathol. 1994, 16, 93–108. [Google Scholar] [CrossRef]
- Guimarães, R.L.; Stotz, H.U. Oxalate production by Sclerotinia sclerotiorum deregulates guard cells during Infection. Plant. Physiol. 2004, 136, 3703–3711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Issam, S.M.; Mohamed, G.; Dominique, L.M.; Thierry, M.; Farid, L.; Nejib, M. A β-Glucosidase from Sclerotinia sclerotiorum biochemical characterization and use in oligosaccharide synthesis. Appl. Biochem. Biotechnol. 2004, 112, 63–78. [Google Scholar] [CrossRef]
- Ellouze, O.E.; Loukil, S.; Marzouki, M.N. Cloning and molecular characterization of a new fungal xylanase gene from Sclerotinia sclerotiorum S2. BMB Rep. 2011, 44, 653–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, B.; Kabbage, M.; Kim, H.J.; Britt, R.; Dickman, M.B. Tipping the balance: Sclerotinia sclerotiorum secreted oxalic acid suppresses host defenses by manipulating the host redox environment. PLoS Pathog. 2011, 7, e1002107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabbage, M.; Williams, B.; Dickman, M.B. Cell death control: The interplay of apoptosis and autophagy in the pathogenicity of Sclerotinia sclerotiorum. PLoS Pathog. 2013, 9, e1003287. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Xie, J.; Cheng, J.; Li, G.; Yi, X.; Jiang, D.; Fu, Y. Novel secretory protein Ss-Caf1 of the plant-pathogenic fungus Sclerotinia sclerotiorum is required for host penetration and normal sclerotial development. Mol. Plant-Microbe Interact. 2014, 27, 40–55. [Google Scholar] [CrossRef] [Green Version]
- Lyu, X.; Shen, C.; Fu, Y.; Xie, J.; Jiang, D.; Li, G.; Cheng, J. A small secreted virulence-related protein is essential for the necrotrophic interactions of Sclerotinia sclerotiorum with its host plants. PLoS Pathog. 2016, 12, e1005435. [Google Scholar] [CrossRef]
- Yang, G.; Tang, L.; Gong, Y.; Xie, J.; Fu, Y.; Jiang, D.; Li, G.; Collinge, D.B.; Chen, W.; Cheng, J. A cerato-platanin protein SsCP1 targets plant PR1 and contributes to virulence of Sclerotinia sclerotiorum. New Phytol. 2018, 217, 739–755. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Yang, G.; Ma, M.; Liu, X.; Li, B.; Xie, J.; Fu, Y.; Chen, T.; Yu, Y.; Chen, W.; et al. An effector of a necrotrophic fungal pathogen targets the calcium-sensing receptor in chloroplasts to inhibit host resistance. Mol. Plant Pathol. 2020, 21, 686–701. [Google Scholar] [CrossRef] [Green Version]
- Bolton, M.D.; Thomma, B.P.; Nelson, B.D. Sclerotinia sclerotiorum (Lib.) de Bary: Biology and molecular traits of a cosmopolitan pathogen. Mol. Plant Pathol. 2006, 7, 1–16. [Google Scholar] [CrossRef]
- Clarkson, J.P.; Phelps, K.; Whipps, J.M.; Young, C.S.; Smith, J.A.; Watling, M. Forecasting sclerotinia disease on lettuce: A predictive model for carpogenic germination of Sclerotinia sclerotiorum sclerotia. Phytopathology 2007, 97, 621–631. [Google Scholar] [CrossRef] [Green Version]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 6 April 2022).
- Kunst, L.; Samuels, L. Plant cuticles shine: Advances in wax biosynthesis and export. Curr. Opin. Plant Biol. 2009, 12, 721–727. [Google Scholar] [CrossRef]
- Kong, L.; Liu, Y.; Zhi, P.; Wang, X.; Xu, B.; Gong, Z.; Chang, C. Origins and Evolution of Cuticle Biosynthetic Machinery in Land Plants. Plant Physiol. 2020, 184, 1998–2010. [Google Scholar] [CrossRef]
- Lewandowska, M.; Keyl, A.; Feussner, I. Wax biosynthesis in response to danger: Its regulation upon abiotic and biotic stress. New Phytol. 2020, 227, 698–713. [Google Scholar] [CrossRef] [Green Version]
- Arya, G.C.; Sarkar, S.; Manasherova, E.; Aharoni, A.; Cohen, H. The Plant Cuticle: An Ancient Guardian Barrier Set Against Long-Standing Rivals. Front. Plant Sci. 2021, 12, 663165. [Google Scholar] [CrossRef]
- Bhanot, V.; Fadanavis, S.V.; Panwar, J. Revisiting the architecture, biosynthesis and functional aspects of the plant cuticle: There is more scope. Environ. Exp. Bot. 2021, 183, 104364. [Google Scholar] [CrossRef]
- Arya, G.C.; Cohen, H. The multifaceted roles of fungal cutinases during infection. J. Fungi 2022, 8, 199. [Google Scholar] [CrossRef]
- Stepinski, D.; Kwiatkowska, M.; Wojtczak, A.; Polit, J.T.; Dominguez, E.; Heredia, A.; Poplonska, K. The role of cutinsomes in plant cuticle formation. Cells 2020, 9, 1778. [Google Scholar] [CrossRef]
- Renault, H.; Alber, A.; Horst, N.A.; Basilio Lopes, A.; Fich, E.A.; Kriegshauser, L.; Wiedemann, G.; Ullmann, P.; Herrgott, L.; Erhardt, M.; et al. A phenol-enriched cuticle is ancestral to lignin evolution in land plants. Nat. Commun. 2017, 8, 14713. [Google Scholar] [CrossRef]
- Muller, C.; Riederer, M. Plant surface properties in chemical ecology. J. Chem. Ecol. 2005, 31, 2621–2651. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, E.; Cuartero, J.; Heredia, A. An overview on plant cuticle biomechanics. Plant Sci. 2011, 181, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Yeats, T.H.; Rose, J.K. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serrano, M.; Coluccia, F.; Torres, M.; L’Haridon, F.; Métraux, J.-P. The cuticle and plant defense to pathogens. Front. Plant Sci. 2014, 5, 274. [Google Scholar] [CrossRef] [Green Version]
- Ziv, C.; Zhao, Z.; Gao, Y.G.; Xia, Y. Multifunctional roles of plant cuticle during plant-pathogen interactions. Front. Plant Sci. 2018, 9, 1088. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Choi, D.; Suh, M.C. Cuticle ultrastructure, cuticular lipid composition, and gene expression in hypoxia-stressed Arabidopsis stems and leaves. Plant Cell Rep. 2017, 36, 815–827. [Google Scholar] [CrossRef]
- Philippe, G.; Sorensen, I.; Jiao, C.; Sun, X.; Fei, Z.; Domozych, D.S.; Rose, J.K. Cutin and suberin: Assembly and origins of specialized lipidic cell wall scaffolds. Curr. Opin. Plant Biol. 2020, 55, 11–20. [Google Scholar] [CrossRef]
- Yang, S.L.; Tran, N.; Tsai, M.Y.; Ho, C.K. Misregulation of MYB16 expression causes stomatal cluster formation by disrupting polarity during asymmetric cell divisions. Plant Cell 2022, 34, 455–476. [Google Scholar] [CrossRef]
- Raffaele, S.; Leger, A.; Roby, D. Very long chain fatty acid and lipid signaling in the response of plants to pathogens. Plant Signal. Behav. 2009, 4, 94–99. [Google Scholar] [CrossRef] [Green Version]
- Javelle, M.; Vernoud, V.; Rogowsky, P.M.; Ingram, G.C. Epidermis: The formation and functions of a fundamental plant tissue. New Phytol. 2011, 189, 17–39. [Google Scholar] [CrossRef]
- Aragon, W.; Reina-Pinto, J.J.; Serrano, M. The intimate talk between plants and microorganisms at the leaf surface. J. Exp. Bot. 2017, 68, 5339–5350. [Google Scholar] [CrossRef]
- Lim, G.-H.; Liu, H.; Yu, K.; Liu, R.; Shine, M.B.; Fernandez, J.; Burch-Smith, T.; Mobley, J.K.; McLetchie, N.; Kachroo, A.; et al. The plant cuticle regulates apoplastic transport of salicylic acid during systemic acquired resistance. Sci. Adv. 2020, 6, eaaz0478. [Google Scholar] [CrossRef]
- Philippe, G.; Geneix, N.; Petit, J.; Guillon, F.; Sandt, C.; Rothan, C.; Lahaye, M.; Marion, D.; Bakan, B. Assembly of tomato fruit cuticles: A cross-talk between the cutin polyester and cell wall polysaccharides. New Phytol. 2020, 226, 809–822. [Google Scholar] [CrossRef]
- Chassot, C.; Nawrath, C.; Metraux, J.P. Cuticular defects lead to full immunity to a major plant pathogen. Plant J. 2007, 49, 972–980. [Google Scholar] [CrossRef] [Green Version]
- Skamnioti, P.; Gurr, S.J. Magnaporthe grisea cutinase2 mediates appressorium differentiation and host penetration and is required for full virulence. Plant Cell 2007, 19, 2674–2689. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Su, L.; Chen, J.; Wu, J. Cutinase: Characteristics, preparation, and application. Biotechnol. Adv. 2013, 31, 1754–1767. [Google Scholar] [CrossRef]
- Fu, H.; Chung, K.R.; Gai, Y.; Mao, L.; Li, H. The basal transcription factor II H subunit Tfb5 is required for stress response and pathogenicity in the tangerine pathotype of Alternaria alternata. Mol. Plant Pathol. 2020, 21, 1337–1352. [Google Scholar] [CrossRef]
- Villafana, R.T.; Rampersad, S.N. Diversity, structure, and synteny of the cutinase gene of Colletotrichum species. Ecol. Evol. 2020, 10, 1425–1443. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.Z.; Zhu, X.Q.; Fan, J.; Guo, L.Y. The cutinase Bdo_10846 play an important role in the virulence of Botryosphaeria dothidea and in inducing the wart symptom on apple plant. Int. J. Mol. Sci. 2021, 22, 1910. [Google Scholar] [CrossRef]
- Longhi, S.; Cambillau, C. Structure-activity of cutinase, a small lipolytic enzyme. Biochim. Biophys. Acta 1999, 1441, 185–196. [Google Scholar] [CrossRef]
- Nikolaivits, E.; Kanelli, M.; Dimarogona, M.; Topakas, E. A middle-aged enzyme still in its prime: Recent advances in the field of cutinases. Catalysts 2018, 8, 612. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Ashby, A.M.; Johnstone, K. Molecular evidence that the extracellular cutinase Pbc1 is required for pathogenicity of Pyrenopeziza brassicae on oilseed rape. Mol. Plant-Microbe Interact. 2003, 16, 545–552. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Hou, J.; Wang, Y.; Jin, Y.; Borth, W.; Zhao, F.; Liu, Z.; Hu, J.; Zuo, Y. Genome-wide identification, classification and expression analysis in fungal-plant interactions of cutinase gene family and functional analysis of a putative ClCUT7 in Curvularia lunata. Mol. Genet. Genom. 2016, 291, 1105–1115. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, J.; Li, D.-W.; Zheng, L.; Huang, J. CglCUT1 gene required for cutinase activity and pathogenicity of Colletotrichum gloeosporioides causing anthracnose of Camellia oleifera. Eur. J. Plant Pathol. 2016, 147, 103–114. [Google Scholar] [CrossRef]
- Stahl, D.J.; Schäfer, W. Cutinase is not required for fungal pathogenicity on pea. Plant Cell 1992, 4, 621–629. [Google Scholar]
- Crowhurst, R.N.; Binnie, S.J.; Bowen, J.K.; Hawthorne, B.T.; Plummer, K.M.; Rees-George, J.; Rikkerink, E.H.; Templeton, M.D. Effect of disruption of a cutinase gene (cutA) on virulence and tissue specificity of Fusarium solani f. sp. cucurbitae race 2 toward Cucurbita maxima and C. moschata. Mol. Plant-Microbe Interact. 1997, 10, 355–368. [Google Scholar] [CrossRef] [Green Version]
- Kan, J.A.v.; Klooster, J.W.v.t.; Wagemakers, C.A.; Dee, D.C.; Vlugt-Bergmans, C.J.v.d. Cutinase A of Botrytis cinerea is expressed, but not essential, during penetration of gerbera and tomato. Mol. Plant-Microbe Interact. 1997, 10, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Li, P.; Liu, H.; Chen, X.; Huang, J.; Luo, C.; Li, G.; Hsiang, T.; Collinge, D.B.; Zheng, L. A novel transcription factor UvCGBP1 regulates development and virulence of rice false smut fungus Ustilaginoidea virens. Virulence 2021, 12, 1563–1579. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Zhang, B.; Gai, Y.; Sun, X.; Chung, K.R.; Li, H. Cell-wall-degrading enzymes required for virulence in the host selective toxin-producing necrotroph Alternaria alternata of citrus. Front. Microbiol. 2019, 10, 2514. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-H.; Chiu, C.-M.; Roubtsova, T.; Chou, C.-M.; Bostock, R.M. Overexpression of a redox-regulated cutinase gene, MfCUT1, increases virulence of the brown rot pathogen Monilinia fructicola on Prunus spp. Mol. Plant-Microbe Interact. 2010, 23, 176–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gui, Y.J.; Zhang, W.Q.; Zhang, D.D.; Zhou, L.; Short, D.P.G.; Wang, J.; Ma, X.F.; Li, T.G.; Kong, Z.Q.; Wang, B.L.; et al. A Verticillium dahliae extracellular cutinase modulates plant immune responses. Mol. Plant-Microbe Interact. 2018, 31, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Rong, W.; Massart, S.; Zhang, Z. Genome-wide identification and expression analysis of cutinase gene family in Rhizoctonia cerealis and functional study of an active cutinase RcCUT1 in the fungal-wheat interaction. Front. Microbiol. 2018, 9, 1813. [Google Scholar] [CrossRef]
- Peyraud, R.; Mbengue, M.; Barbacci, A.; Raffaele, S. Intercellular cooperation in a fungal plant pathogen facilitates host colonization. Proc. Natl. Acad. Sci. USA 2019, 116, 3193–3201. [Google Scholar] [CrossRef] [Green Version]
- Dallal Bashi, Z.; Rimmer, S.R.; Khachatourians, G.G.; Hegedus, D.D. Factors governing the regulation of Sclerotinia sclerotiorum cutinase A and polygalacturonase 1 during different stages of infection. Can. J. Microbiol. 2012, 58, 605–616. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, Q.; Cao, S.; Zhao, T.; Chen, L.; Zhuang, P.; Zhou, X.; Gao, Z. A novel protein elicitor (SsCut) from Sclerotinia sclerotiorum induces multiple defense responses in plants. Plant Mol. Biol. 2014, 86, 495–511. [Google Scholar] [CrossRef]
- Godoy, G.; Steadman, J.; Dickman, M.; Dam, R. Use of mutants to demonstrate the role of oxalic acid in pathogenicity of Sclerotinia sclerotiorum on Phaseolus vulgaris. Physiol. Mol. Plant Pathol. 1990, 37, 179–191. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Allen, G.C.; Flores-Vergara, M.A.; Krasynanski, S.; Kumar, S.; Thompson, W.F. A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethylammonium bromide. Nat. Protoc. 2006, 1, 2320–2325. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jacobs, K.A.; Collins-Racie, L.A.; Colbert, M.; Duckett, M.; Golden-Fleet, M.; Kelleher, K.; Kriz, R.; LaVallie, E.R.; Merberg, D.; Spaulding, V.; et al. A genetic selection for isolating cDNAs encoding secreted proteins. Gene 1997, 198, 289–296. [Google Scholar] [CrossRef]
- Lee, S.J.; Kim, B.D.; Rose, J.K. Identification of eukaryotic secreted and cell surface proteins using the yeast secretion trap screen. Nat. Protoc. 2006, 1, 2439–2447. [Google Scholar] [CrossRef]
- Catlett, N.L.; Lee, B.-N.; Yoder, O.C.; Turgeon, B.G. Split-marker recombination for efficient targeted deletion of fungal genes. Fungal Genet. Rep. 2003, 50, 9–11. [Google Scholar] [CrossRef] [Green Version]
- Rollins, J.A. The Sclerotinia sclerotiorum pac1 gene is required for sclerotial development and virulence. Mol. Plant-Microbe Interact. 2003, 16, 785–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Jiang, D.; Xie, J.; Cheng, J.; Li, G.; Yi, X.; Fu, Y. Ss-Sl2, a novel cell wall protein with PAN modules, is essential for sclerotial development and cellular integrity of Sclerotinia sclerotiorum. PLoS ONE 2012, 7, e34962. [Google Scholar] [CrossRef] [PubMed]
- Tomme, P.; Boraston, A.; McLean, B.; Kormos, J.; Creagh, A.L.; Sturch, K.; Gilkes, N.R.; Haynes, C.A.; Warren, R.A.; Kilburn, D.G. Characterization and affinity applications of cellulose-binding domains. J. Chromatogr. B Biomed. Sci. Appl. 1998, 715, 283–296. [Google Scholar] [CrossRef]
- Skamnioti, P.; Furlong, R.F.; Gurr, S.J. Evolutionary history of the ancient cutinase family in five filamentous Ascomycetes reveals differential gene duplications and losses and in Magnaporthe grisea shows evidence of sub- and neo-functionalization. New Phytol. 2008, 180, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.A.; Procopiou, J. The cuticles of citrus species composition of leaf and fruit waxes. J. Sci. Food Agric. 1975, 26, 1093–1101. [Google Scholar] [CrossRef]
- Bellincampi, D.; Cervone, F.; Lionetti, V. Plant cell wall dynamics and wall-related susceptibility in plant-pathogen interactions. Front. Plant Sci. 2014, 5, 228. [Google Scholar] [CrossRef] [Green Version]
- Laluk, K.; Mengiste, T. Necrotroph attacks on plants: Wanton destruction or covert extortion? Arab. Book 2010, 8, e0136. [Google Scholar] [CrossRef] [Green Version]
- Auyong, A.S.M. The role of cutinase and its impact on pathogenicity of Colletotrichum truncatum. J. Plant Pathol. Microbiol. 2015, 6, 3. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Yan, P.; Guan, M.; Han, S.; Qiao, T.; Lin, T.; Zhu, T.; Li, S. Comparative Transcriptomics and Gene Knockout Reveal Virulence Factors of Arthrinium phaeospermum in Bambusa pervariabilis x Dendrocalamopsis grandis. J. Fungi 2021, 7, 1001. [Google Scholar] [CrossRef]
- Maiti, I.B.; Kolattukudy, P.E. Prevention of fungal infection of plants by specific inhibition of cutinase. Science 1979, 205, 507–508. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Y.; Li, B.; Yang, X.; Dong, Y.; Qiu, D. A Verticillium dahliae pectate lyase induces plant immune responses and contributes to virulence. Front. Plant Sci. 2018, 9, 1271. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Analysis of the cutinase protein SsCut1. (A) The evolutionary relationship of SsCut1 and its homologs from other fungi determined with the maximum-likelihood algorithm. Branch lengths are proportional to the average probability of change for characters on that branch. The phylogeny was constructed with Mega 6.0 using the neighbor-joining method (parameters: 1000 bootstraps). (B) The amino acid sequence alignment of SsCut1, SsCut, BcCutA and BcCutB. The black stars indicate the conserved GYSQG catalytic site. The rectangle box indicates the conserved CBM1 domain.
Figure 1.
Analysis of the cutinase protein SsCut1. (A) The evolutionary relationship of SsCut1 and its homologs from other fungi determined with the maximum-likelihood algorithm. Branch lengths are proportional to the average probability of change for characters on that branch. The phylogeny was constructed with Mega 6.0 using the neighbor-joining method (parameters: 1000 bootstraps). (B) The amino acid sequence alignment of SsCut1, SsCut, BcCutA and BcCutB. The black stars indicate the conserved GYSQG catalytic site. The rectangle box indicates the conserved CBM1 domain.

Figure 2.
Expression patterns of SsCut1 of S. sclerotiorum at different stages of S. sclerotiorum. (A) Expression patterns of the SsCut1 in culture on PDA medium at 20 °C for 1–7 d (days). (B) Expression patterns of the SsCut1 during the infection of A. thaliana at 20 °C for 0–24 h (hours). The S. sclerotiorum β-tubulin gene was used as an internal control to normalize the data. Error bars represent the standard error (n = 3).
Figure 2.
Expression patterns of SsCut1 of S. sclerotiorum at different stages of S. sclerotiorum. (A) Expression patterns of the SsCut1 in culture on PDA medium at 20 °C for 1–7 d (days). (B) Expression patterns of the SsCut1 during the infection of A. thaliana at 20 °C for 0–24 h (hours). The S. sclerotiorum β-tubulin gene was used as an internal control to normalize the data. Error bars represent the standard error (n = 3).

Figure 3.
SsCut1 has a functional signal peptide and is localized in the plant cell wall. (A) Validation of the secretion function of the SsCut1 signal peptide by the yeast secretion trap screen assay. Signal peptide of SsCut1 was fused in frame to the yeast invertase sequence in pSUC2 vector and expressed in YTK12 strains. The functional signal peptide of Avr1b was used as a positive control, while the YTK12 and pSUC2 empty plasmid was used as a negative control. (B) The invertase activity in TTC medium. TTC encounters raffinose breakdown products to produce triphenylformazan, which shows a red reaction to confirm that a functional signal peptide can cause sucrose invertase to be secreted. (C) Subcellular localization of SsCut1 in N. benthamiana epidermal cells. SsCut1-GFP localized in the plant cell wall. The fluorescence of GFP was monitored at 2 d post-agroinfiltration using confocal laser scanning microscopy. Bar = 20 µm.
Figure 3.
SsCut1 has a functional signal peptide and is localized in the plant cell wall. (A) Validation of the secretion function of the SsCut1 signal peptide by the yeast secretion trap screen assay. Signal peptide of SsCut1 was fused in frame to the yeast invertase sequence in pSUC2 vector and expressed in YTK12 strains. The functional signal peptide of Avr1b was used as a positive control, while the YTK12 and pSUC2 empty plasmid was used as a negative control. (B) The invertase activity in TTC medium. TTC encounters raffinose breakdown products to produce triphenylformazan, which shows a red reaction to confirm that a functional signal peptide can cause sucrose invertase to be secreted. (C) Subcellular localization of SsCut1 in N. benthamiana epidermal cells. SsCut1-GFP localized in the plant cell wall. The fluorescence of GFP was monitored at 2 d post-agroinfiltration using confocal laser scanning microscopy. Bar = 20 µm.

Figure 4.
The deletion of SsCut1 has no significant effect on oxalate production, hypha morphology and infection cushions formation. (A) Qualitative determination of acid produced by the wild-type strain and SsCut1 transformants on PDA medium containing 0.005% (w/v) bromophenol blue dye as a pH indicator. The presence of yellow indicates that acid was produced. Photographs were taken at 36 hpi. (B) In vitro hyphal development of the wild-type strain and SsCut1 transformants. All strains were cultured on PDA medium for 36 hpi. Hyphal tips were observed under a dissecting microscope. Bars = 500 µm. (C) Infection cushions formation of wild-type strain and SsCut1 transformants. Microscopic observation of infection cushions of wild-type strain and SsCut1 transformants on onion epidermal cell layer after staining with trypan blue. Photographs were taken at 14 hpi. Bar = 20 µm.
Figure 4.
The deletion of SsCut1 has no significant effect on oxalate production, hypha morphology and infection cushions formation. (A) Qualitative determination of acid produced by the wild-type strain and SsCut1 transformants on PDA medium containing 0.005% (w/v) bromophenol blue dye as a pH indicator. The presence of yellow indicates that acid was produced. Photographs were taken at 36 hpi. (B) In vitro hyphal development of the wild-type strain and SsCut1 transformants. All strains were cultured on PDA medium for 36 hpi. Hyphal tips were observed under a dissecting microscope. Bars = 500 µm. (C) Infection cushions formation of wild-type strain and SsCut1 transformants. Microscopic observation of infection cushions of wild-type strain and SsCut1 transformants on onion epidermal cell layer after staining with trypan blue. Photographs were taken at 14 hpi. Bar = 20 µm.

Figure 5.
SsCut1 knock-out mutants showing reduced virulence on the detached leaves of oilseed rape and Arabidopsis leaves. (A) Lesions formation on oilseed rape leaves inoculated with wild-type strain (1980) and SsCut1 transformants, the photographs were taken at 48 h post-inoculation (hpi). (B) Statistical results of lesion area on oilseed rape leaves. (C) Lesions formation on A. thaliana leaves inoculated with wild-type strain and SsCut1 transformants, the photographs were taken at 36 hpi. (D) Statistical results of lesion area on A. thaliana leaves. Bars indicate ± SE (n = 4). Statistical significance is indicated in the graph (one-way ANOVA): ** p < 0.01, **** p < 0.0001. The experiments were performed three times with similar results.
Figure 5.
SsCut1 knock-out mutants showing reduced virulence on the detached leaves of oilseed rape and Arabidopsis leaves. (A) Lesions formation on oilseed rape leaves inoculated with wild-type strain (1980) and SsCut1 transformants, the photographs were taken at 48 h post-inoculation (hpi). (B) Statistical results of lesion area on oilseed rape leaves. (C) Lesions formation on A. thaliana leaves inoculated with wild-type strain and SsCut1 transformants, the photographs were taken at 36 hpi. (D) Statistical results of lesion area on A. thaliana leaves. Bars indicate ± SE (n = 4). Statistical significance is indicated in the graph (one-way ANOVA): ** p < 0.01, **** p < 0.0001. The experiments were performed three times with similar results.

Figure 6.
Cutinase activity of wild-type strain and SsCut1 transformants. All strains were cultured on PDA medium. The hyphal of wild-type strain and SsCut1 transformants was collected at 36 hpi. Cutinase activity levels were examined with the enzyme-linked immunosorbent assay (ELISA) method. Bars indicate ± SE. Statistical significance is indicated in the graph (one-way ANOVA): ** p < 0.01, *** p < 0.001.
Figure 6.
Cutinase activity of wild-type strain and SsCut1 transformants. All strains were cultured on PDA medium. The hyphal of wild-type strain and SsCut1 transformants was collected at 36 hpi. Cutinase activity levels were examined with the enzyme-linked immunosorbent assay (ELISA) method. Bars indicate ± SE. Statistical significance is indicated in the graph (one-way ANOVA): ** p < 0.01, *** p < 0.001.

Figure 7.
SsCut1 triggers plant defense responses and plays a role in the Sclerotinia–plant interaction. (A) SsCut1 promote flg22-triggered reactive oxygen species burst. The N. benthamiana with SsCut1 and empty vector were treated with 100 µg/mL flg22. Bars indicate ± SE. Error bars represent the SE from ten biological replicates. (B) Induction of defense response genes by SsCut1. SsCut1 induces NbPR1 expression in N. benthamiana. Relative transcript accumulation of NbPR1, NbNPR1, NbPDF1.2 genes determined by RT-qPCR. The transcript level of NbEF1α in N. benthamiana was used to normalize the expression levels in different samples. Error bars represent the SE from three replicates. (C) Expression of SsCut1 in N. benthamiana increases plant susceptibility to the SsCut1 knock-out mutant ΔSsCut1-3. Leaves of N. benthamiana were agroinfiltrated with Agrobacterium tumefaciens containing empty vector or pCNF-SsCut1. The wild-type strain and SsCut1 transformants were inoculated 48 h after agroinfiltration. Photographs were taken at 48 hpi. (D) Statistical results of lesion area on N. benthamiana leaves. In this experiment, four independent replicates were performed. Bars indicate ± SE. Statistical significance is indicated in the graph (one-way ANOVA): *** p < 0.001.
Figure 7.
SsCut1 triggers plant defense responses and plays a role in the Sclerotinia–plant interaction. (A) SsCut1 promote flg22-triggered reactive oxygen species burst. The N. benthamiana with SsCut1 and empty vector were treated with 100 µg/mL flg22. Bars indicate ± SE. Error bars represent the SE from ten biological replicates. (B) Induction of defense response genes by SsCut1. SsCut1 induces NbPR1 expression in N. benthamiana. Relative transcript accumulation of NbPR1, NbNPR1, NbPDF1.2 genes determined by RT-qPCR. The transcript level of NbEF1α in N. benthamiana was used to normalize the expression levels in different samples. Error bars represent the SE from three replicates. (C) Expression of SsCut1 in N. benthamiana increases plant susceptibility to the SsCut1 knock-out mutant ΔSsCut1-3. Leaves of N. benthamiana were agroinfiltrated with Agrobacterium tumefaciens containing empty vector or pCNF-SsCut1. The wild-type strain and SsCut1 transformants were inoculated 48 h after agroinfiltration. Photographs were taken at 48 hpi. (D) Statistical results of lesion area on N. benthamiana leaves. In this experiment, four independent replicates were performed. Bars indicate ± SE. Statistical significance is indicated in the graph (one-way ANOVA): *** p < 0.001.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gong, Y.; Fu, Y.; Xie, J.; Li, B.; Chen, T.; Lin, Y.; Chen, W.; Jiang, D.; Cheng, J. Sclerotinia sclerotiorum SsCut1 Modulates Virulence and Cutinase Activity. J. Fungi 2022, 8, 526. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050526
AMA Style
Gong Y, Fu Y, Xie J, Li B, Chen T, Lin Y, Chen W, Jiang D, Cheng J. Sclerotinia sclerotiorum SsCut1 Modulates Virulence and Cutinase Activity. Journal of Fungi. 2022; 8(5):526. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050526
Chicago/Turabian StyleGong, Yingdi, Yanping Fu, Jiatao Xie, Bo Li, Tao Chen, Yang Lin, Weidong Chen, Daohong Jiang, and Jiasen Cheng. 2022. "Sclerotinia sclerotiorum SsCut1 Modulates Virulence and Cutinase Activity" Journal of Fungi 8, no. 5: 526. https://0-doi-org.brum.beds.ac.uk/10.3390/jof8050526
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.