Social, Reproductive and Contextual Influences on Fecal Glucocorticoid Metabolites in Captive Yangtze Finless Porpoises (Neophocaena asiaeorientalis asiaeorientalis) and Bottlenose Dolphins (Tursiops truncatus)

Abstract
:1. Introduction
2. Material and Methods
2.1. Subjects, Housing and Group Composition
2.2. Data Collection
2.2.1. Fecal Sample Collection
2.2.2. Contextual Events and Situations Associated with Fecal Samples
2.2.3. Sample Processing and GCMs Assay
2.3. Statistical Analyses
3. Results
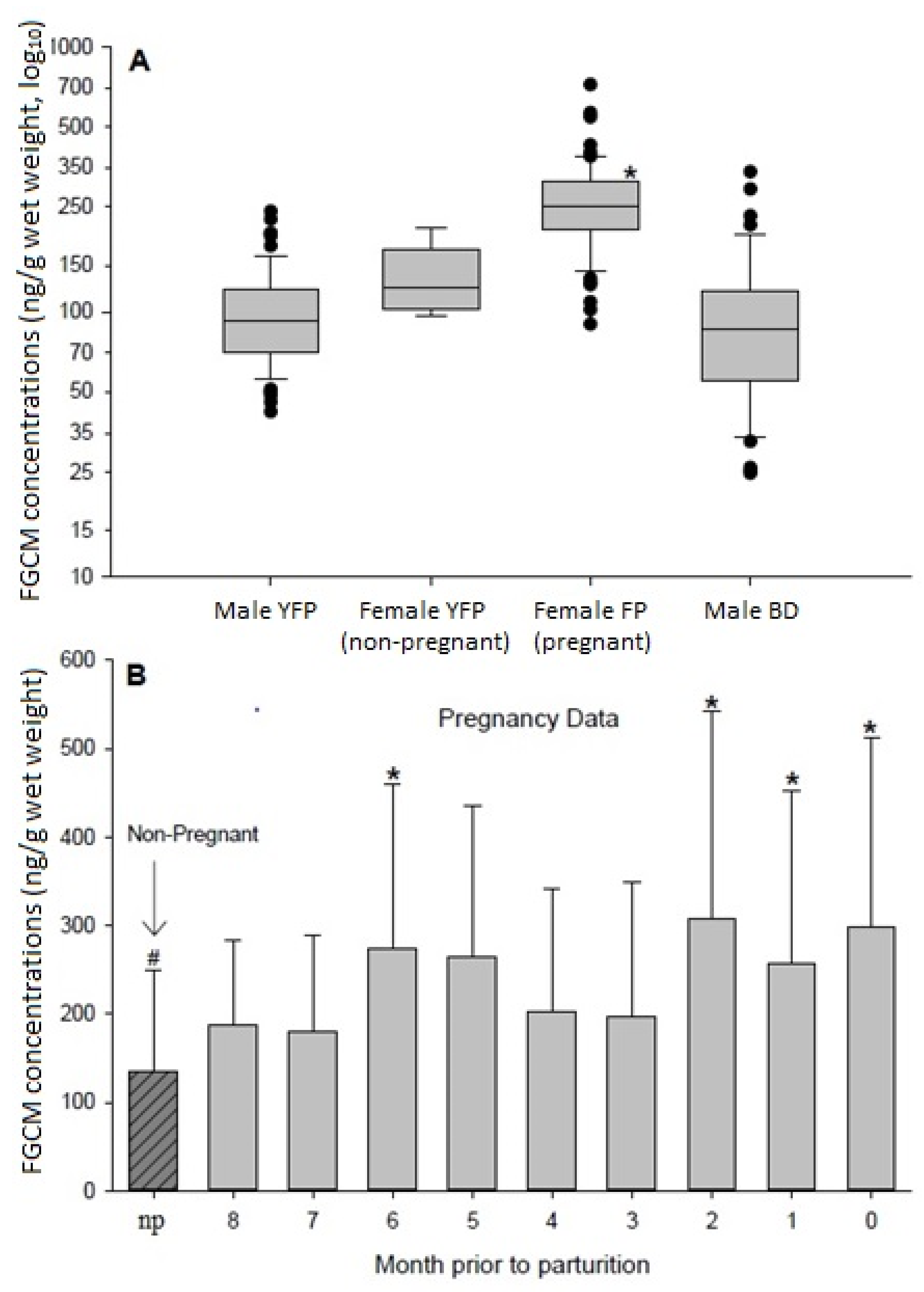
3.1. FGCM Concentration Characteristics and Physiological Validations
3.2. FGCM Concentrations and Contextual Data
4. Discussion
4.1. FGCM Concentrations Biological Validation
4.2. FGCM Concentrations and Contextual Data
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Levine, S.; Ursin, H. Stress—Neurobiology and neuroendocrinology. In What Is Stress? Brown, M.R., Koob, G.F., Rivier, C., Eds.; Marcel Dekker: New York, NY, USA, 1991. [Google Scholar]
- Reeder, D.; Kramer, K. Stress in free-ranging mammals: Integrating physiology, ecology, and natural history. J. Mammal. 2005, 86, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Romero, L.M.; Dickens, M.J.; Cyr, N.E. The reactive scope model—A new model integrating homeostasis allostasis and stress. Horm. Behav. 2009, 55, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, D.S.; Woods, S.C. Clarifying the roles of homeostasis and allostasis in physiologic regulation. Psychol. Rev. 2014, 121, 225–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomson, C.A.; Geraci, J.R. Cortisol aldosterone and leucocytes in the stress response of bottlenose dolphins Tursiops truncatus. Can. J. Fish. Aquat. Sci. 1986, 43, 1010–1016. [Google Scholar] [CrossRef]
- Champagne, C.D.; Kellar, N.M.; Trego, M.L.; Brendan, D.; Rudy, B.; Wasser, S.K.; Booth, R.K.; Crocker, D.E.; Houser, D.S. Comprehensive Endocrine Response to Acute Stress in the Bottlenose Dolphin from Serum Blubber and Feces. Gen. Comp. Endocrinol. 2018, 266, 178–193. [Google Scholar] [CrossRef]
- Steinman, K.J.; Robeck, T.R.; Fetter, G.A.; Schmitt, T.L.; Osborn, S.; DiRocco, S.; Nollens, H.H.; O’Brien, J.K. Circulating and excreted corticosteroids and metabolites, hematological, and serum chemistry parameters in the killer whale (Orcinus orca) before and after a biological stress response. Front. Mar. Sci. 2020, 6, 830. [Google Scholar] [CrossRef]
- Dhabhar, F.S.; McEwen, B.S. Acute stress enhances while chronic stress suppresses immune function in vivo: A potential role for leukocyte trafficking. BrainBehav. Immun. 1997, 11, 286–306. [Google Scholar] [CrossRef] [Green Version]
- Dobson, H.; Smith, R.F. What is stress and how does it affect reproduction? Anim. Reprod. Sci. 2000, 60–61, 743–752. [Google Scholar] [CrossRef]
- Boonstra, R. Coping with changing northern environments: The role of the stress axis in birds and mammals. Int. Comp. Biol. 2004, 44, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Charbonnel, N.; Chaval, Y.; Berthier, K.; Deter, J.; Morand, S.; Palme, R.; Cosson, J.F. Stress and demographic decline: A potential effect mediated by impairment of reproduction and immune function in cyclic vole populations. Physiol. Biochem. Zool. 2007, 81, 63–73. [Google Scholar] [CrossRef]
- Dhabhar, F.S. Effects of stress on immune function: The good the bad and the beautiful. Immun. Res. 2014, 58, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Mumby, H.S.; Mar, K.U.; Thitaram, C.; Courtiol, A.; Towiboon, P.; Min-Oo, Z.; Htut-Aung, Y.; Brown, J.L.; Lummaa, V. Stress and body condition are associated with climate and demography in Asian elephants. Cons. Physiol. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shave, J.R.; Derocher, A.E.; Cherry, S.G.; Thiemann, G.W. Chronic stress and body condition of wolf-killed prey in prince albert national park saskatchewan. Conserv. Physiol. 2019, 7, coz037. [Google Scholar] [CrossRef] [PubMed]
- Mostl, E.; Palme, R. Hormones as indicators of stress. Domest. Anim. Endocrinol. 2002, 23, 67–74. [Google Scholar] [CrossRef]
- Sheriff, M.; Dantzer, B.; Delehanty, B.; Palme, R.; Boonstra, R. Measuring stress in wildlife: Techniques for quantifying glucocorticoids. Oecologia 2011, 166, 869–887. [Google Scholar] [CrossRef] [PubMed]
- Broom, D.M. Quality of life means welfare: How is it related to other concepts and assessed? Anim. Welf. 2007, 16, 45–53. [Google Scholar]
- Busch, D.S.; Hayward, L.S. Stress in a conservation context: A discussion of glucocorticoid actions and how concentrations change with conservation-relevant variables. Biol. Conserv. Lett. 2009, 142, 2844–2853. [Google Scholar] [CrossRef]
- Cooke, S.J.; O’Connor, C.M. Making conservation physiology relevant to policy makers and conservation practitioners. Conserv. Lett. 2010, 3, 159–166. [Google Scholar] [CrossRef]
- Atkinson, S.; Crocker, D.; Houser, D.; Mashburn, K. Stress physiology in marine mammals: How well do they fit the terrestrial model? J. Comp. Physiol. B 2015, 185, 463–486. [Google Scholar] [CrossRef]
- St Aubin, D.J.; Geraci, J.R. Adaptive Changes in Hematologic and Plasma Chemical. Can. J. Fish. Aquat. Sci. 1989, 46, 796–803. [Google Scholar]
- Tryland, M.; Thoresen, S.I.; Kovacs, K.M.; Lydersen, C. Serum chemistry of free-ranging white whales (Delphinapterus leucas) in Svalbard. Vet. Clin. Pathol. 2006, 35, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, T.L.; St. Aubin, D.J.; Schaefer, A.M.; Dunn, J.L. Baseline diurnal variations and stress-induced changes of stress hormones in three captive beluga whales Delphinapterus leucas. Mar. Mamm. Sci. 2010, 26, 635–647. [Google Scholar]
- Spoon, T.R.; Romano, T.A. Neuroimmunological response of beluga whales (Delphinapterus leucas) to translocation and a novel social environment. Brain Behav. Immun. 2012, 26, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, M.; Tobayama, T.; Katsumata, E.; Yoskioka, M.; Aida, K. Serum Cortisol Concentrations in Captive Killer Whale and Bottlenose Dolphin. Fish. Sci. 1998, 64, 643–647. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, M.; Uchida, S.; Ueda, K.; Tobayama, T.; Katsumata, E.; Yoshioka, M.; Aida, K. Diurnal and annual changes in serum cortisol concentrations in Indo-Pacific bottlenose dolphins Tursiops aduncus and killer whales Orcinus orca. Gen. Comp. Endocrinol. 2003, 132, 427–433. [Google Scholar] [CrossRef]
- Wasser, S.K.; Lundin, J.I.; Ayres, K.; Seely, E.; Giles, D.; Balcomb, K.; Hemplemann, J.; Parsons, K.; Booth, R. Population growth is limited by nutritional impacts on pregnancy success in Southern Resident killer whales (Orcinus orca). PLoS ONE 2017, 12, e0179824. [Google Scholar] [CrossRef] [PubMed]
- Ayres, K.L.; Booth, R.K.; Hemplemann, J.A.; Koski, K.I.; Emmons, C.K.; Baird, R.W.; Balcomb-Bartok, K.; Hanson, M.B.; Ford, M.J.; Wasser, S.K. Distinguishing the impacts inadequate prey and vessel on an endangered killer whale (Orcinus orca) population. PLoS ONE 2012, 7, e36842. [Google Scholar] [CrossRef] [Green Version]
- Robeck, T.; Steinman, K.; O’Brien, J. Characterization and longitudinal monitoring of serum androgens and glucocorticoids during normal pregnancy in the killer whale (Orcinus orca). Gen. Comp. Endocrinol. 2017, 247, 116–129. [Google Scholar] [CrossRef]
- St. Aubin, D.J.; Ridgway, S.H.; Wells, R.S.; Rhinehart, H.L. Dolphin thyroid and adrenal hormones: Circulating concentrations in wild and semi domesticated Tursiops truncatus and influence of sex age and season. Mar. Mamm. Sci. 1996, 12, 1–13. [Google Scholar] [CrossRef]
- Ortiz, R.M.; Worthy, G.A.J. Effects of capture on adrenal steroid and vasopressin. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2000, 125, 317–324. [Google Scholar] [CrossRef]
- Noda, K.; Aoki, M.; Akiyoshi, H.; Asaki, H.; Ogata, T.; Yamauchi, K.; Shimada, T.; Ohashi, F. Effect of bovine lactoferrin on the immune responses of captive bottlenosed dolphins (Tursiops truncatus) being transported over long distances. Vet. Rec. 2006, 159, 885–888. [Google Scholar] [PubMed]
- Pedernra-Romano, C.; Valdez, R.A.; Singh, S.; Chiappa, X.; Romano, M.C.; Galindo, F. Salivary cortisol in captive dolphins (Tursiops truncates): A non-invasive technique. Anim. Welf. 2006, 1, 359–362. [Google Scholar]
- Houser, D.S.; Yeates, L.C.; Crocker, D.E. Cold Stress Induces an Adrenocortical Response in Bottlenose Dolphins (Tursiops truncatus). J. Zoo Wildl. Med. 2011, 42, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Luna Blasio, A.; Valdez Pérez, R.; Romano Pardo, M.; Galindo, M.F. Maintenance behaviour and cortisol concentrations in bottlenose dolphins (Tursiops truncatus) in closed and open facilities. Vet. Mex. 2012, 43, 103–112. [Google Scholar]
- Hao, Y.; Zhao, Q.; Wu, H.; Chen, D.; Gong, C.; Li, L.; Wang, D. Physiological responses to capture and handling of free-ranging male Yangtze finless porpoises (Neophocaena phocaenoides asiaeorientalis). Mar. Freshw. Behav. Physiol. 2009, 42, 315–327. [Google Scholar] [CrossRef]
- Morton, D.J.; Anderson, E.; Foggin, C.M.; Kock, M.D.; Tiran, E.P. Plasma cortisol as an indicator of stress due to capture and translocation in wildlife species. Vet. Rec. 1995, 136, 60–63. [Google Scholar] [CrossRef]
- Pankhurst, N.; Dedualj, M. Effects of capture and recovery on plasma levels of cortisol, lactate and gonadal steroids in a natural population of rainbow trout. J. Fish Biol. 2005, 45, 1013–1025. [Google Scholar] [CrossRef]
- Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 2019, 199, 229–243. [Google Scholar] [CrossRef]
- Hunt, K.E.; Rolland, R.M.; Kraus, S.D.; Wasser, S.K. Analysis of fecal glucocorticoids in the North Atlantic right whale (Eubalaena glacialis). Gen. Comp. Endocrinol. 2006, 148, 260–272. [Google Scholar] [CrossRef]
- Biancani, B.; Da Dalt, L.; Gallina, G.; Capolongo, F.; Gabai, G. Fecal cortisol radioimmunoassay to monitor adrenal gland activity in the bottlenose dolphin (Tursiops truncatus) under human care. Mar. Mamm. Sci. 2017, 3, 1014–1034. [Google Scholar] [CrossRef]
- Rolland, R.M.; Parks, S.E.; Hunt, K.E.; Castellote, M.; Corkeron, P.J.; Nowacek, D.P.; Wasser, S.K.; Kraus, S.D. Evidence that ship noise increases stress in right whales. Proc. R. Soc. B 2012, B 279, 2363–2368. [Google Scholar] [CrossRef]
- Hunt, K.E.; Robbins, J.; Buck, C.L.; Bérubé, M.; Rolland. R., M. Evaluation of fecal hormones for noninvasive research on reproduction and stress in humpback whales (Megaptera novaeangliae). Gen. Comp. Endocrinol. 2019, 280, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Rolland, R.M.; McLellan, W.A.; Moore, M.J.; Harms, C.A.; Burgess, E.A.; Hunt, K.E. Fecal glucocorticoids and anthropogenic injury and mortality in North Atlantic right whales Eubalaena glacialis. Endanger. Species Res. 2017, 34, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Copland, M.D.; Needham, D.J. Hematological and biochemical changes associated with transport of dolphins (Tursiops truncatus). In Proceedings of the AAAM Conferences, Hong Kong, 18–22 May 1992. [Google Scholar]
- Cockrem, J.F. Individual variation in glucocorticoid stress responses in animals. Gen. Comp. Endocrinol. 2013, 181, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Waples, K.A.; Gales, N.J. Evaluating and minimising social stress in the care of captive bottlenose dolphins (Tursiops aduncus). Zoo Biol. 2002, 21, 5–26. [Google Scholar] [CrossRef]
- Brando, S.; Broom, D.M.; Acasuso-Rivero, C.; Clark, F. Optimal marine mammal welfare under human care: Current efforts and future directions. Behav. Proc. 2018, 156, 16–36. [Google Scholar] [CrossRef] [PubMed]
- Breuner, C.W.; Delehanty, B.; Boonstra, R. Evaluating stress in natural populations of vertebrates: Total cort is not good enough. Funct. Ecol. 2013, 27, 24–36. [Google Scholar] [CrossRef]
- MacDougall-Shackleton, S.A.; Bonier, F.; Romero, L.M.; Moore, I.T. Glucocorticoids and “Stress” are not synonymous. Integr. Org. Biol. 2019, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Breuner, C.W.; Patterson, S.H.; Hahn, T.P. In search of relationships between the acute adrenocortical response and fitness. Gen. Comp. Endocrinol. 2008, 157, 288–295. [Google Scholar] [CrossRef]
- Bonier, F.; Martin, P.R.; Moore, I.T.; Wingfield, J.C. Do baseline glucocorticoids predict fitness? Trends Ecol. Evol. 2009, 24, 634–642. [Google Scholar] [CrossRef]
- International Union for Conservation of Nature (IUCN 2013). IUCN Red List of Threatened Species. 2013. Available online: http://iucnredlist.org/ (accessed on 19 December 2018).
- Munro, C.J.; Lasley, B.L. Non-radiometric methods for immunoassay of steroid hormones. In Technology and Application In Polypeptide and Steroid Hormone Detection; Anderson, B.D., Haseltine, F.D., Eds.; A.R. Liss Inc.: New York, NY, USA, 1988; pp. 289–329. [Google Scholar]
- West, B.T.; Welch, K.B.; Galecki, A.T. Linear Mixed Models: In A Practical Guide Using Statistical Software, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 199–248. [Google Scholar]
- Judge, G.G.; Griffiths, W.E.; Hill, R.C.; Lütkepohl, H.; Lee, T.C. The Theory and Practice of Econometrics, 2nd ed.; Wiley: New York, NY, USA, 1985. [Google Scholar] [CrossRef]
- Akaike, H. Information theory and an extension of the maximum likelihood principle. In Selected Papers of Hirotugu Akaike; Parzen, E., Tanabe, K., Kitagawa, G., Eds.; Springer: New York, NY, USA, 1988. [Google Scholar] [CrossRef]
- Chang, Q.; Zhou, K.Y. The growth and reproduction of finless porpoise, Neophocaena phocaenoides, in the Yangtze River and Yellow/Bohai Sea. J. Nan. Norm. Univ. 1995, 18, 114–124. [Google Scholar]
- Champagne, C.D.; Kellar, N.M.; Crocker, D.E.; Wasser, S.K.; Booth, R.K.; Trego, M.L.; Houser, D.S. Blubber cortisol qualitatively reflects circulating cortisol concentrations in bottlenose dolphins. Mar. Mamm. Sci. 2017, 33, 134–153. [Google Scholar] [CrossRef]
- Steinman, K.J.; Robeck, T.R.; O’Brien, J.K. Characterization of estrogens testosterone and cortisol in normal bottlenose dolphin (Tursiops truncatus) pregnancy. Gen. Comp. Endocrinol. 2016, 226, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela-Molina, M.; Atkinson, S.; Mashburn, K.; Gendron, D.; Brownell, R. Fecal steroid hormones reveal reproductive state in female blue whales sampled in the Gulf of California, Mexico. Gen. Comp. Endocrinol. 2018, 261, 127–135. [Google Scholar] [CrossRef]
- Ash, R.W.; Heap, R.B. Oestrogen, progesterone and corticosteroid concentrations in peripheral plasma of sow during pregnancy, parturition, lactation and after weaning. J. Endocrionol. 1975, 64, 141–154. [Google Scholar] [CrossRef]
- Bolt, R.J.; Van Weissenbruch, M.M.; Lafeber, H.N.; Delemarre-van de Waal, H.A. Glucocorticoids and lung development in the fetus and preterm infant. Pediatr. Pulmonol. 2001, 32, 76–91. [Google Scholar] [CrossRef]
- Bassett, J.M.; Thorburn, G.D. Foetal plasma corticosteroids and the initiation of parturition in the sheep. J. Endocrinol. 1969, 44, 285–286. [Google Scholar] [CrossRef]
- Liggins, G.C. The role of cortisol in preparing the fetus for birth. Reprod. Fertil. Dev. 1994, 6, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Challis, J.R.G.; Matthews, S.G.; Gibb, W.; Lye, S.J. Endocrine and paracrine regulation of birth at term and preterm. Endocrine Rev. 2000, 21, 514–550. [Google Scholar] [CrossRef]
- Hart, L.B.; Wells, R.S.; Kellar, N.; Balmer, B.C.; Hohn, A.A.; Lamb, S.V.; Rowles, T.; Zolman, E.S.; Schwacke, L.H. Adrenal hormones in common bottlenose dolphins (Tursiops truncatus): Influential factors and reference intervals. PLoS ONE 2015, 10, e0127432. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, J.K.; Steinman, K.J.; Fetter, G.A.; Robeck, T.R. Androgen and glucocorticoid production in the male killer whale (Orcinus orca): Influence of age, maturity and environmental factors. J. Androl. 2017, 5, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Orlov, M.V.; Mukhlya, A.M.; Kulikov, N.A. Hormonal indices in the bottle-nosed dolphin Tursiops truncatus in the norm and in the dynamics of experimental stress. Zhurnal Evolyutsionnoi Biokhimii Fiziol. 1988, 24, 557–563. [Google Scholar]
- Orlov, M.; Mukhlya, A.M.; Kuz’min, A.A. Hormonal and electrolyte changes in cetacean blood after capture and during experimental stress. J. Evol. Biochem. Physiol. 1991, 27, 197–205. [Google Scholar]
- Touma, C.; Palme, R. Measuring fecal glucocorticoid metabolites in mammals and birds: The importance of validation. Ann. N. Y. Acad. Sci. 2005, 1046, 54–74. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, Y.; Guo, L.; Gu, B.; Liu, H.; Hou, A.; Liu, X.; Sun, L.; Liu, D. Stereotypic Behavior and Fecal Cortisol Level in Captive Giant Pandas in Relation to Environmental Enrichment. Zoo Biol. 2006, 25, 445–459. [Google Scholar] [CrossRef]
- Delfour, F.; Beyer, H. Assessing the effectiveness of environmental enrichment in bottlenose dolphins (Tursiops truncatus). Zoo Biol. 2012, 31, 137–150. [Google Scholar]
- Serres, A.; Delfour, F. Environmental changes and anthropogenic factors modulate social play in captive bottlenose dolphins (Tursiops truncatus). Zoo Biol. 2017, 36, 99–111. [Google Scholar] [CrossRef]
- Serres, A.; Hao, Y.; Wang, D. Swimming features in captive odontocetes: Indicative of animals’ emotional state? Behav. Proc. 2020, 170, 103998. [Google Scholar] [CrossRef]
- Serres, A.; Hao, Y.; Wang, D. The frequency of solitary behaviours in captive odontocetes is modulated by environmental and social factors. Int. J. Comp. Psychol. 2020, 32. Available online: https://escholarship.org/uc/item/42h458vs (accessed on 1 August 2020).
- Mason, G.J. Species differences in responses to captivity: Stress, welfare and the comparative method. Trends Ecol. Evol. 2010, 25, 713–721. [Google Scholar] [CrossRef] [Green Version]
- Carlstead, K.; Brown, J.L. Relationships Between Patterns of Fecal Corticoid Excretion and Behavior Reproduction and Environmental Factors in Captive Black (Diceros bicornis) and White (Ceratotherium simum) Rhinoceros. Zoo Biol. 2005, 24, 215–232. [Google Scholar] [CrossRef]
- Pifarré, M.; Valdez, R.; González-Rebeles, C.; Vázquez Marta Romano, C.; Galindo, F. The effect of zoo visitors on the behaviour and fecal cortisol of the Mexican wolf (Canis lupus baileyi). Appl. Anim. Behav. Sci. 2012, 136, 57–66. [Google Scholar] [CrossRef]
- Rajagopal, T.; Archunan, G.; Sekar, M. Impact of Zoo Visitors on the Fecal Cortisol Concentrations and Behavior of an Endangered Species: Indian Blackbuck (Antelope cervicapra L.). J. Appl. Anim. Welf. Sci. 2011, 14, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.; Hosey, G.R. Interaction sequences between chimpanzees and human visitors at the zoo. Zoo Biol. 1995, 14, 431–444. [Google Scholar] [CrossRef]
- Hosey, G.R. Zoo animals and their human audiences: What is the visitor effect? Anim. Welf. 2000, 9, 343–357. [Google Scholar]
- Palme, R.; Fischer, P.; Schildorfer, H.; Ismael, M.N. Excretion of infused 14C-steroid hormones via faeces and urine in domestic livestock. Anim. Reprod. Sci. 1996, 43, 43–63. [Google Scholar] [CrossRef]
- Mostl, E.; Messmann, S.; Bagu, E.; Robia, C.; Palme, R. Measurement of Glucocorticoid Metabolite Concentrations in Faeces of Domestic Livestock. J. Vet. Med. A 1999, 46, 621–631. [Google Scholar]
- Ridgway, S.H. Homeostasis in the aquatic environment. In Mammals of the Sea: Biology and Medicine; Ridgway, S.H., Thomas, C.C., Eds.; Charles C. Thomas Publisher: Springfield, IL, USA, 1972; pp. 590–747. [Google Scholar]
- Fair, P.A.; Becker, P.R. Review of stress in marine mammals. J. Aquat. Ecosyst. Stress Recovery 2000, 7, 335–354. [Google Scholar] [CrossRef]
- St. Aubin, D.J.; Dierauf, L.A. Stress and marine mammals. In CRC Handbook Of Marine Mammal Medicine, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar] [CrossRef]
- Kuo, T.; McQueen, A.; Chen, T.C.; Wang, J.C. Regulation of Glucose Homeostasis by Glucocorticoids. Adv. Exp. Med. Biol. 2015, 872, 92–126. [Google Scholar]
- Whitham, J.C.; Wielebnowski, N.C. New directions for zoo animal welfare science. Appl. Anim. Behav. Sci. 2013, 147, 247–260. [Google Scholar] [CrossRef]


{kind=link}
| Name | Species | Sex | Age (Year) | Weight (kg) | Length (m) | N |
|---|---|---|---|---|---|---|
| MP1 | Yangtze finless porpoise a | M | 8 | NA | 1.57 | 44 |
| FP1 * | Yangtze finless porpoise a | F | 8 | NA | 1.45 | 70 |
| MP2 | Yangtze finless porpoise a | M | 14 | NA | 1.56 | 24 |
| FP2 * | Yangtze finless porpoise a | F | 11 | NA | 1.47 | 7 |
| MD1 | Bottlenose dolphin b | M | 13 | 280 | 2.74 | 4 |
| MD2 | Bottlenose dolphin b | M | 14 | 290 | 2.69 | 35 |
| MD3 | Bottlenose dolphin b | M | 13 | 260 | 2.70 | 10 |
| Species | Sampling Time | Season | Social Grouping | Enrichment | Visitors |
|---|---|---|---|---|---|
| YFP | Morning (8:00–11:00) | Winter (December–February) | All together | None | None |
| Early afternoon (14:00–15:30) | Spring (March–May) | Separated a | Toys c | Low d | |
| Late afternoon (16:00–17:30) | Summer (June–August) | Alone b | High d | ||
| Fall (September–November) | |||||
| BD | Early afternoon (13:30–14:00) | Winter (December–February) | All together | None | Scale from 0 to 3 based on the public present during shows (0: park closed, “None”, 1: one to two rows full of visitors, “Few”, 2: three to six rows, “Moderate”, 3: more than six rows, “Many”) |
| Spring (March–May) | Separateda | Toysc | |||
| Summer (June–August) | |||||
| Fall (September–November) |
| Category | Condition | N | Marginal Mean FGCM | 95% CI |
|---|---|---|---|---|
| Stage of Pregnancy | Non-pregnant | 8 | 131 A | 107–161 |
| Early | 14 | 202 A, B | 127–321 | |
| Mid | 29 | 243 B | 206–286 | |
| Late | 24 | 253 B | 211–302 | |
| MPP | Non-pregnant | 8 | 135 A | 114–161 |
| 8 | 2 | 188 A, B | 95–372 | |
| 7 | 4 | 180 A, B | 109–298 | |
| 6 | 8 | 273 B | 186–403 | |
| 5 | 5 | 264 A, B | 172–406 | |
| 4 | 9 | 202 A, B | 139–293 | |
| 3 | 15 | 197 A, B | 152–255 | |
| 2 | 11 | 308 B | 234–403 | |
| 1 | 9 | 256 B | 195–341 | |
| 0 | 4 | 299 B | 213–341 |
| Category | Species | Condition | N | Marginal Mean | 95% CI |
|---|---|---|---|---|---|
| Status | YFP & BD | Pregnant Female | 67 | 236 A | 190–293 |
| Non-pregnant Female | 8 | 134 B | 93–194 | ||
| YFP Male | 69 | 100 B | 83–121 | ||
| BD Male | 49 | 89 B | 73–109 | ||
| Season | YFP | Winter | 55 | 165 A | 154–175 |
| Spring | 29 | 143 A,B | 125–164 | ||
| Summer | 5 | 111 A,B | 83–147 | ||
| Fall | 56 | 143B | 126–162 | ||
| Time of day | YFP | Morning | 15 | 176 A | 149–207 |
| Early afternoon | 94 | 141 B | 132–151 | ||
| Late afternoon | 36 | 152 A, B | 137–170 | ||
| Enrichment | YFP | Absent | 106 | 155 A | 146–165 |
| Present | 39 | 136 B | 123–150 | ||
| All together | 86 | 140 A | 130–150 | ||
| Social grouping | YFP | Separated | 33 | 153 B | 137–171 |
| Alone | 26 | 182 B | 159–209 | ||
| Vistors | YFP | None | 72 | 151 A | 140–162 |
| Few | 30 | 122 B | 109–138 | ||
| Many | 43 | 170 C | 154–188 | ||
| Season | BD | Winter | 20 | 99 | 82–120 |
| Spring | 12 | 82 | 62–109 | ||
| Summer | 11 | 72 | 54–95 | ||
| Enrichment | BD | Absent | 20 | 135 A | 109–166 |
| Present | 30 | 61 B | 52–73 | ||
| Visitors | BD | None | 15 | 70 A | 56–87 |
| Few | 14 | 113 B | 90–142 | ||
| Moderate | 5 | 64 A | 44–92 | ||
| Many | 11 | 99 A,B | 75–130 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serres, A.; Robeck, T.; Deng, X.; Steinman, K.; Hao, Y.; Wang, D. Social, Reproductive and Contextual Influences on Fecal Glucocorticoid Metabolites in Captive Yangtze Finless Porpoises (Neophocaena asiaeorientalis asiaeorientalis) and Bottlenose Dolphins (Tursiops truncatus). J. Zool. Bot. Gard. 2020, 1, 24-41. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg1010003
Serres A, Robeck T, Deng X, Steinman K, Hao Y, Wang D. Social, Reproductive and Contextual Influences on Fecal Glucocorticoid Metabolites in Captive Yangtze Finless Porpoises (Neophocaena asiaeorientalis asiaeorientalis) and Bottlenose Dolphins (Tursiops truncatus). Journal of Zoological and Botanical Gardens. 2020; 1(1):24-41. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg1010003
Chicago/Turabian StyleSerres, Agathe, Todd Robeck, Xiaojun Deng, Karen Steinman, Yujiang Hao, and Ding Wang. 2020. "Social, Reproductive and Contextual Influences on Fecal Glucocorticoid Metabolites in Captive Yangtze Finless Porpoises (Neophocaena asiaeorientalis asiaeorientalis) and Bottlenose Dolphins (Tursiops truncatus)" Journal of Zoological and Botanical Gardens 1, no. 1: 24-41. https://0-doi-org.brum.beds.ac.uk/10.3390/jzbg1010003