
Structures and Divergent Mechanisms in Capsid Maturation and Stabilization Following Genome Packaging of Human Cytomegalovirus and Herpesviruses

 , , and
, , and
Abstract
:1. Introduction
2. Assembly of Herpesvirus Capsids
2.1. The Major Capsid Protein (MCP)
2.2. The Smallest Capsid Protein (SCP)
2.3. External Scaffolding Proteins: Triplex Proteins
2.4. Maturational Protease and Internal Scaffolding Protein from the Fusion Protein
2.5. The Portal Protein
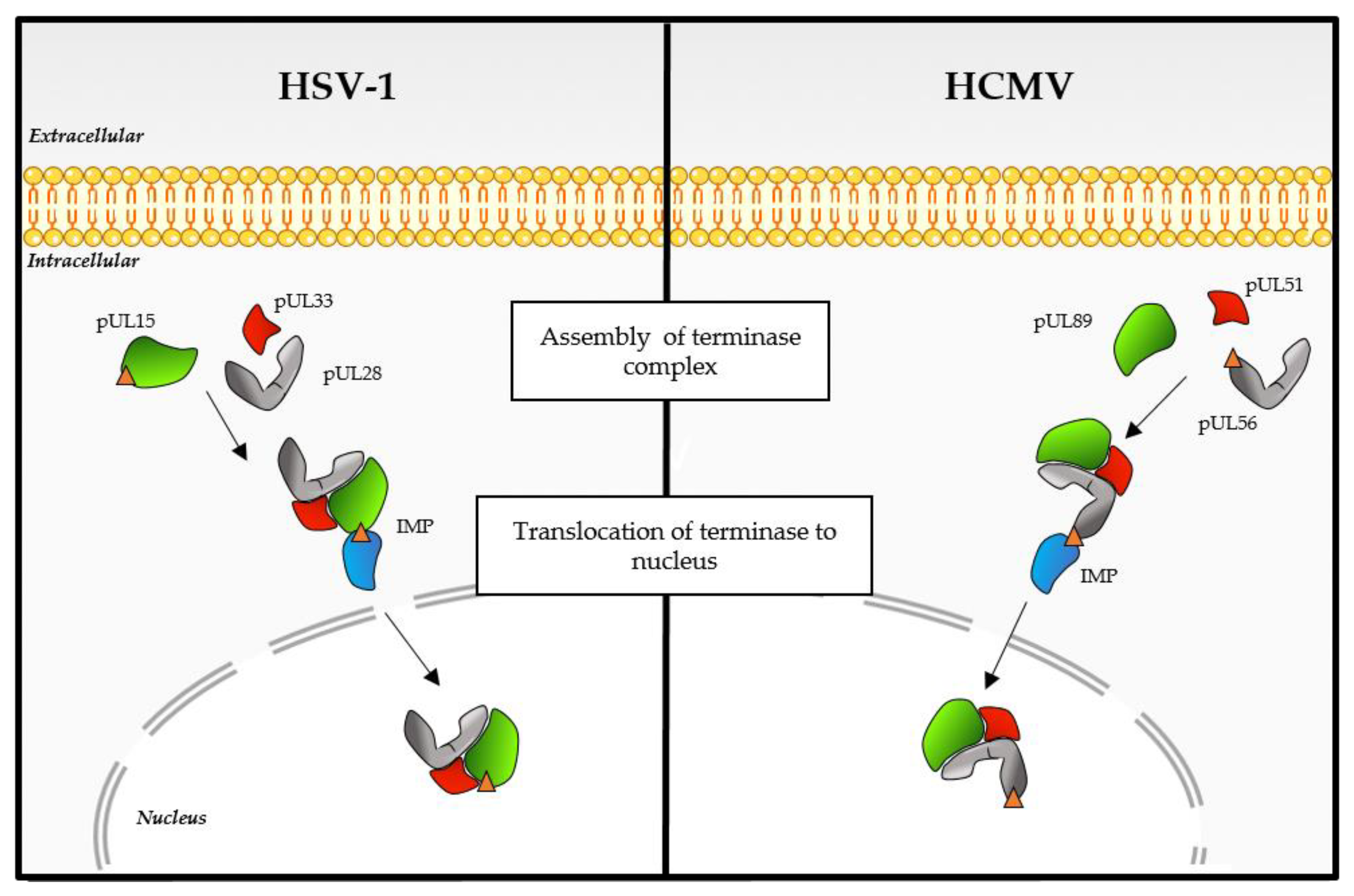
3. Viral DNA Cleavage/Packaging
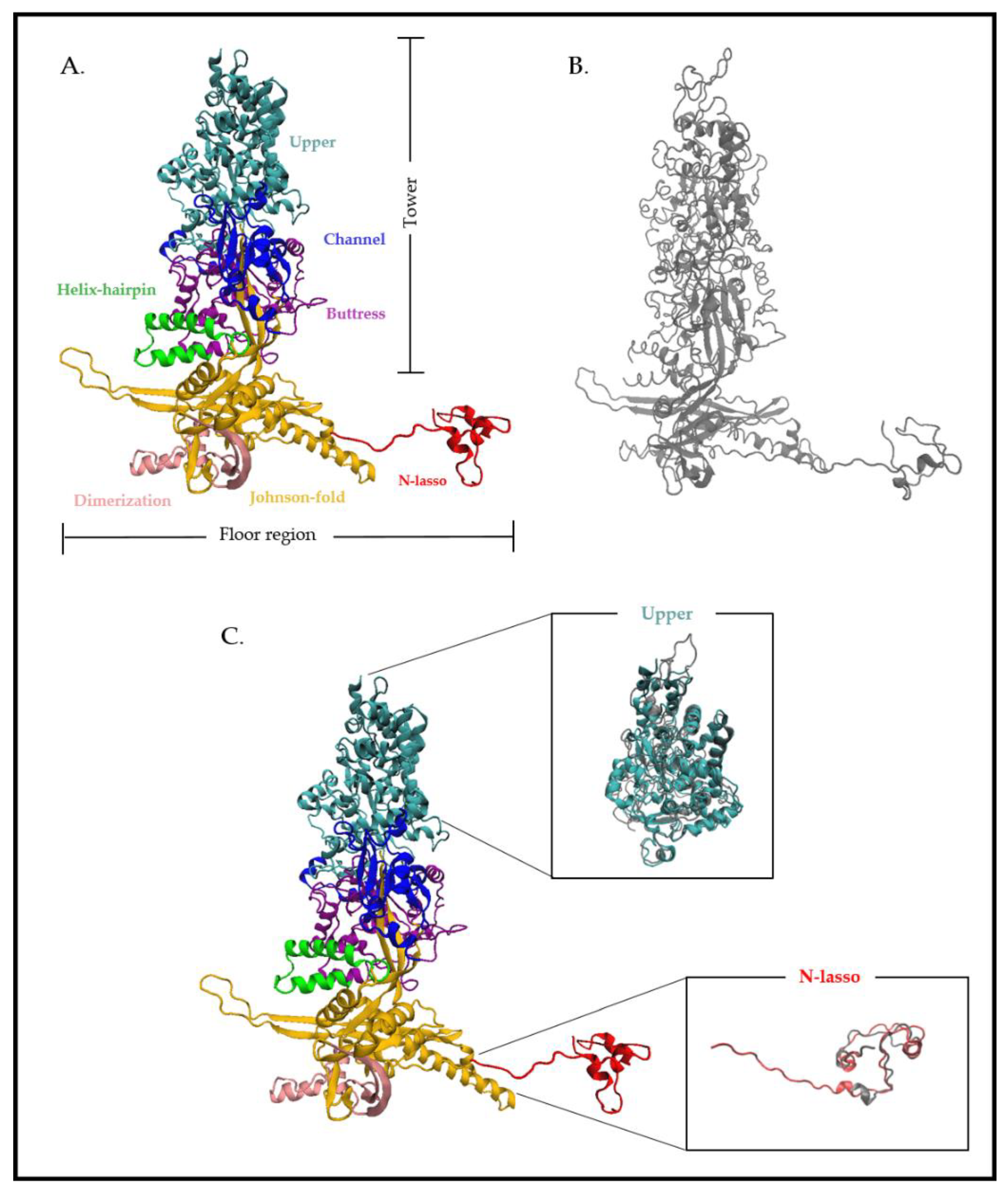
3.1. The Terminase Large Subunit
3.2. The Terminase Small Subunit
3.3. Additional Proteins Contribute to the HCMV DNA Cleavage/Packaging Process
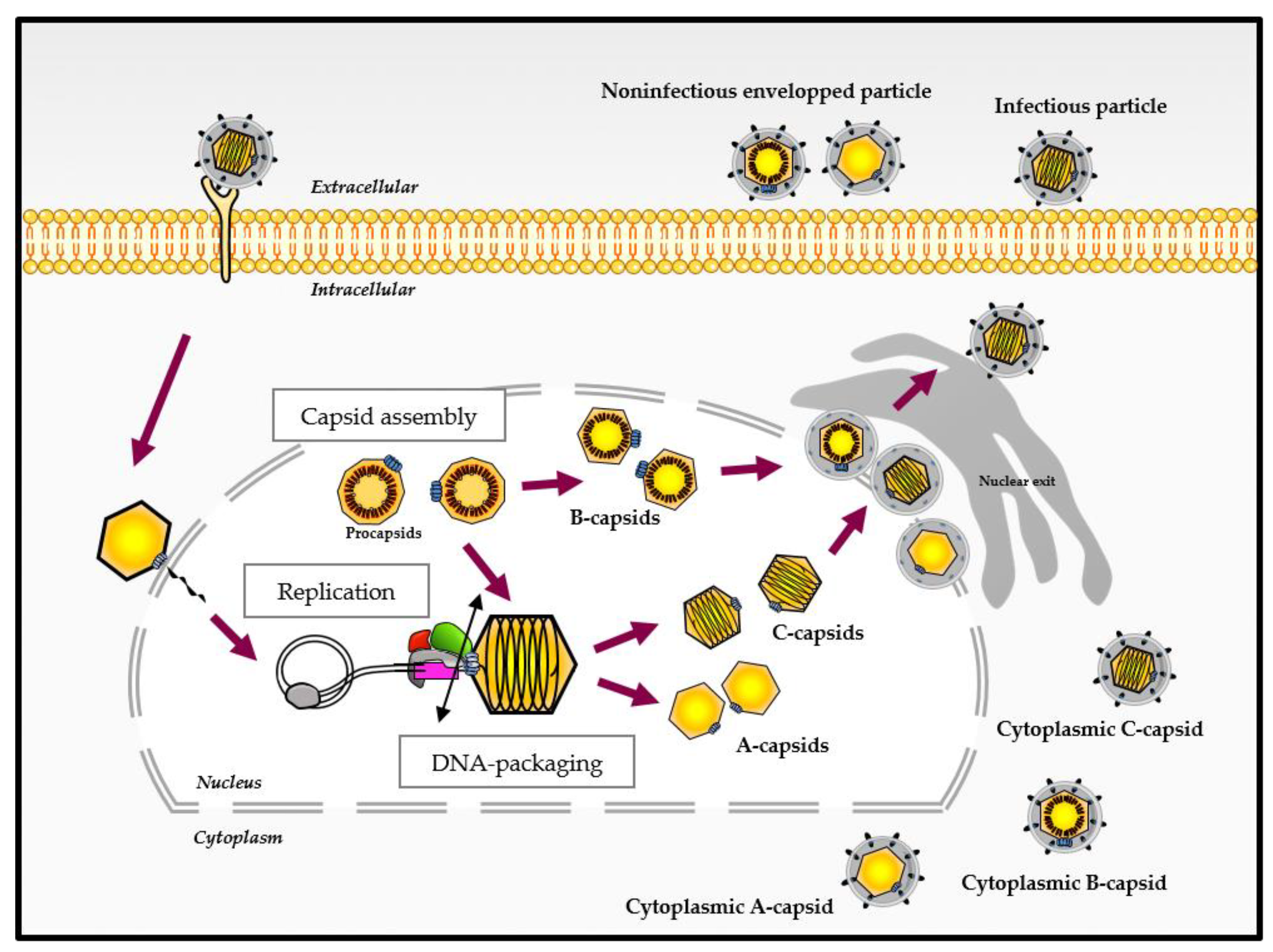
4. Different Capsid Forms Are Present in the Host Cell: A-, B- and C-Capsids
5. Inhibitors of Terminase and Capsid Proteins for the Treatment of Herpesvirus Infections
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Leruez-Ville, M.; Ville, Y. Fetal Cytomegalovirus Infection. Best Pract. Res. Clin. Obstet. Gynaecol. 2017, 38, 97–107. [Google Scholar] [CrossRef]
- Lurain, N.S.; Chou, S. Antiviral Drug Resistance of Human Cytomegalovirus. Clin. Microbiol. Rev. 2010, 23, 689–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Chaer, F.; Shah, D.P.; Chemaly, R.F. How I Treat Resistant Cytomegalovirus Infection in Hematopoietic Cell Transplantation Recipients. Blood 2016, 128, 2624–2636. [Google Scholar] [CrossRef] [Green Version]
- Andouard, D.; Mazeron, M.-C.; Ligat, G.; Couvreux, A.; Pouteil-Noble, C.; Cahen, R.; Yasdanpanah, Y.; Deering, M.; Viget, N.; Alain, S.; et al. Contrasting Effect of New HCMV PUL54 Mutations on Antiviral Drug Susceptibility: Benefits and Limits of 3D Analysis. Antiviral Res. 2016, 129, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Ligat, G.; Cazal, R.; Hantz, S.; Alain, S. The Human Cytomegalovirus Terminase Complex as an Antiviral Target: A Close-up View. FEMS Microbiol. Rev. 2018, 42, 137–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ligat, G.; Muller, C.; Alain, S.; Hantz, S. The terminase complex, a relevant target for the treatment of HCMV infection. Med. Sci. 2020, 36, 367–375. [Google Scholar] [CrossRef]
- Dai, X.; Zhou, Z.H. Structure of the Herpes Simplex Virus 1 Capsid with Associated Tegument Protein Complexes. Science 2018, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Liu, C.; Peng, R.; Zhang, F.-K.; Tong, Z.; Liu, S.; Shi, Y.; Zhao, Z.; Zeng, W.-B.; Gao, G.F.; et al. Cryo-EM Structure of the Varicella-Zoster Virus A-Capsid. Nat. Commun. 2020, 11, 4795. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Jih, J.; Jiang, J.; Zhou, Z.H. Atomic Structure of the Human Cytomegalovirus Capsid with Its Securing Tegument Layer of Pp150. Science 2017, 356. [Google Scholar] [CrossRef] [Green Version]
- Bhella, D.; Rixon, F.J.; Dargan, D.J. Cryomicroscopy of Human Cytomegalovirus Virions Reveals More Densely Packed Genomic DNA than in Herpes Simplex Virus Type 1. J. Mol. Biol. 2000, 295, 155–161. [Google Scholar] [CrossRef]
- Baines, J.D. Herpes Simplex Virus Capsid Assembly and DNA Packaging: A Present and Future Antiviral Drug Target. Trends Microbiol. 2011, 19, 606–613. [Google Scholar] [CrossRef]
- Chee, M.; Rudolph, S.A.; Plachter, B.; Barrell, B.; Jahn, G. Identification of the Major Capsid Protein Gene of Human Cytomegalovirus. J. Virol. 1989, 63, 1345–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, X.; Gong, D.; Lim, H.; Jih, J.; Wu, T.-T.; Sun, R.; Zhou, Z.H. Structure and Mutagenesis Reveal Essential Capsid Protein Interactions for KSHV Replication. Nature 2018, 553, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Arvin, A.; Arvin, A.M. Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2007; ISBN 9781280916946. [Google Scholar]
- Booy, F.P.; Trus, B.L.; Newcomb, W.W.; Brown, J.C.; Conway, J.F.; Steven, A.C. Finding a Needle in a Haystack: Detection of a Small Protein (the 12-KDa VP26) in a Large Complex (the 200-MDa Capsid of Herpes Simplex Virus). Proc. Natl. Acad. Sci. USA 1994, 91, 5652–5656. [Google Scholar] [CrossRef] [Green Version]
- Desai, P.; DeLuca, N.A.; Person, S. Herpes Simplex Virus Type 1 VP26 Is Not Essential for Replication in Cell Culture but Influences Production of Infectious Virus in the Nervous System of Infected Mice. Virology 1998, 247, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Varnum, S.M.; Streblow, D.N.; Monroe, M.E.; Smith, P.; Auberry, K.J.; Paša-Tolic, L.; Wang, D.; Rodland, K.; Wiley, S.; Britt, W.; et al. Identification of Proteins in Human Cytomegalovirus (HCMV) Particles: The HCMV Proteome. J. Virol. 2004, 78, 7. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.Y.; Roizman, B. The Herpes Simplex Virus 1 Gene Encoding a Protease Also Contains within Its Coding Domain the Gene Encoding the More Abundant Substrate. J. Virol. 1991, 65, 5149–5156. [Google Scholar] [CrossRef] [Green Version]
- Wood, L.J.; Baxter, M.K.; Plafker, S.M.; Gibson, W. Human Cytomegalovirus Capsid Assembly Protein Precursor (PUL80.5) Interacts with Itself and with the Major Capsid Protein (PUL86) through Two Different Domains. J. Virol. 1997, 71, 179–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaudet-Miller, M.; Zhang, R.; Durkin, J.; Gibson, W.; Kwong, A.D.; Hong, Z. Virus-Specific Interaction between the Human Cytomegalovirus Major Capsid Protein and the C Terminus of the Assembly Protein Precursor. J. Virol. 1996, 70, 8081–8088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, W. Structure and Assembly of the Virion. Intervirology 1996, 39, 389–400. [Google Scholar] [CrossRef]
- Hong, Z.; Beaudet-Miller, M.; Durkin, J.; Zhang, R.; Kwong, A.D. Identification of a Minimal Hydrophobic Domain in the Herpes Simplex Virus Type 1 Scaffolding Protein Which Is Required for Interaction with the Major Capsid Protein. J. Virol. 1996, 70, 533–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardone, G.; Heymann, J.B.; Cheng, N.; Trus, B.L.; Steven, A.C. Procapsid Assembly, Maturation, Nuclear Exit: Dynamic Steps in the Production of Infectious Herpesvirions. In Viral Molecular Machines; Rossmann, M.G., Rao, V.B., Eds.; Springer: Boston, MA, USA, 2012; Volume 726, pp. 423–439. ISBN 9781461409793. [Google Scholar]
- Singer, G.P.; Newcomb, W.W.; Thomsen, D.R.; Homa, F.L.; Brown, J.C. Identification of a Region in the Herpes Simplex Virus Scaffolding Protein Required for Interaction with the Portal. J. Virol. 2005, 79, 132–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.H.; Macnab, S.J.; Jakana, J.; Scott, L.R.; Chiu, W.; Rixon, F.J. Identification of the Sites of Interaction between the Scaffold and Outer Shell in Herpes Simplex Virus-1 Capsids by Difference Electron Imaging. Proc. Natl. Acad. Sci. USA 1998, 95, 2778–2783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dittmer, A.; Drach, J.C.; Townsend, L.B.; Fischer, A.; Bogner, E. Interaction of the Putative Human Cytomegalovirus Portal Protein PUL104 with the Large Terminase Subunit PUL56 and Its Inhibition by Benzimidazole-d-Ribonucleosides. J. Virol. 2005, 79, 14660–14667. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-T.; Jih, J.; Dai, X.; Bi, G.-Q.; Zhou, Z.H. Cryo-EM Structures of Herpes Simplex Virus Type 1 Portal Vertex and Packaged Genome. Nature 2019, 570, 257–261. [Google Scholar] [CrossRef]
- Albright, B.S.; Nellissery, J.; Szczepaniak, R.; Weller, S.K. Disulfide Bond Formation in the Herpes Simplex Virus 1 UL6 Protein Is Required for Portal Ring Formation and Genome Encapsidation. J. Virol. 2011, 85, 8616–8624. [Google Scholar] [CrossRef] [Green Version]
- Nellissery, J.K.; Szczepaniak, R.; Lamberti, C.; Weller, S.K. A Putative Leucine Zipper within the Herpes Simplex Virus Type 1 UL6 Protein Is Required for Portal Ring Formation. J. Virol. 2007, 81, 8868–8877. [Google Scholar] [CrossRef] [Green Version]
- Bogner, E.; Radsak, K.; Stinski, M.F. The Gene Product of Human Cytomegalovirus Open Reading Frame UL56 Binds the Pac Motif and Has Specific Nuclease Activity. J. Virol. 1998, 72, 2259–2264. [Google Scholar] [CrossRef] [Green Version]
- Theiß, J.; Sung, M.W.; Holzenburg, A.; Bogner, E. Full-Length Human Cytomegalovirus Terminase PUL89 Adopts a Two-Domain Structure Specific for DNA Packaging. PLoS Pathog. 2019, 15, e1008175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borst, E.M.; Kleine-Albers, J.; Gabaev, I.; Babic, M.; Wagner, K.; Binz, A.; Degenhardt, I.; Kalesse, M.; Jonjic, S.; Bauerfeind, R.; et al. The Human Cytomegalovirus UL51 Protein Is Essential for Viral Genome Cleavage-Packaging and Interacts with the Terminase Subunits PUL56 and PUL89. J. Virol. 2013, 87, 1720–1732. [Google Scholar] [CrossRef] [Green Version]
- Borst, E.M.; Wagner, K.; Binz, A.; Sodeik, B.; Messerle, M. The Essential Human Cytomegalovirus Gene UL52 Is Required for Cleavage-Packaging of the Viral Genome. J. Virol. 2008, 82, 2065–2078. [Google Scholar] [CrossRef] [Green Version]
- Borst, E.M.; Bauerfeind, R.; Binz, A.; Stephan, T.M.; Neuber, S.; Wagner, K.; Steinbrück, L.; Sodeik, B.; Lenac Roviš, T.; Jonjić, S.; et al. The Essential Human Cytomegalovirus Proteins PUL77 and PUL93 Are Structural Components Necessary for Viral Genome Encapsidation. J. Virol. 2016. [Google Scholar] [CrossRef] [Green Version]
- DeRussy, B.M.; Tandon, R. Human Cytomegalovirus PUL93 Is Required for Viral Genome Cleavage and Packaging. J. Virol. 2015. [Google Scholar] [CrossRef] [Green Version]
- Köppen-Rung, P.; Dittmer, A.; Bogner, E. Intracellular Distribution of Capsid-Associated PUL77 of Human Cytomegalovirus and Interactions with Packaging Proteins and PUL93. J. Virol. 2016, 90, 5876–5885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
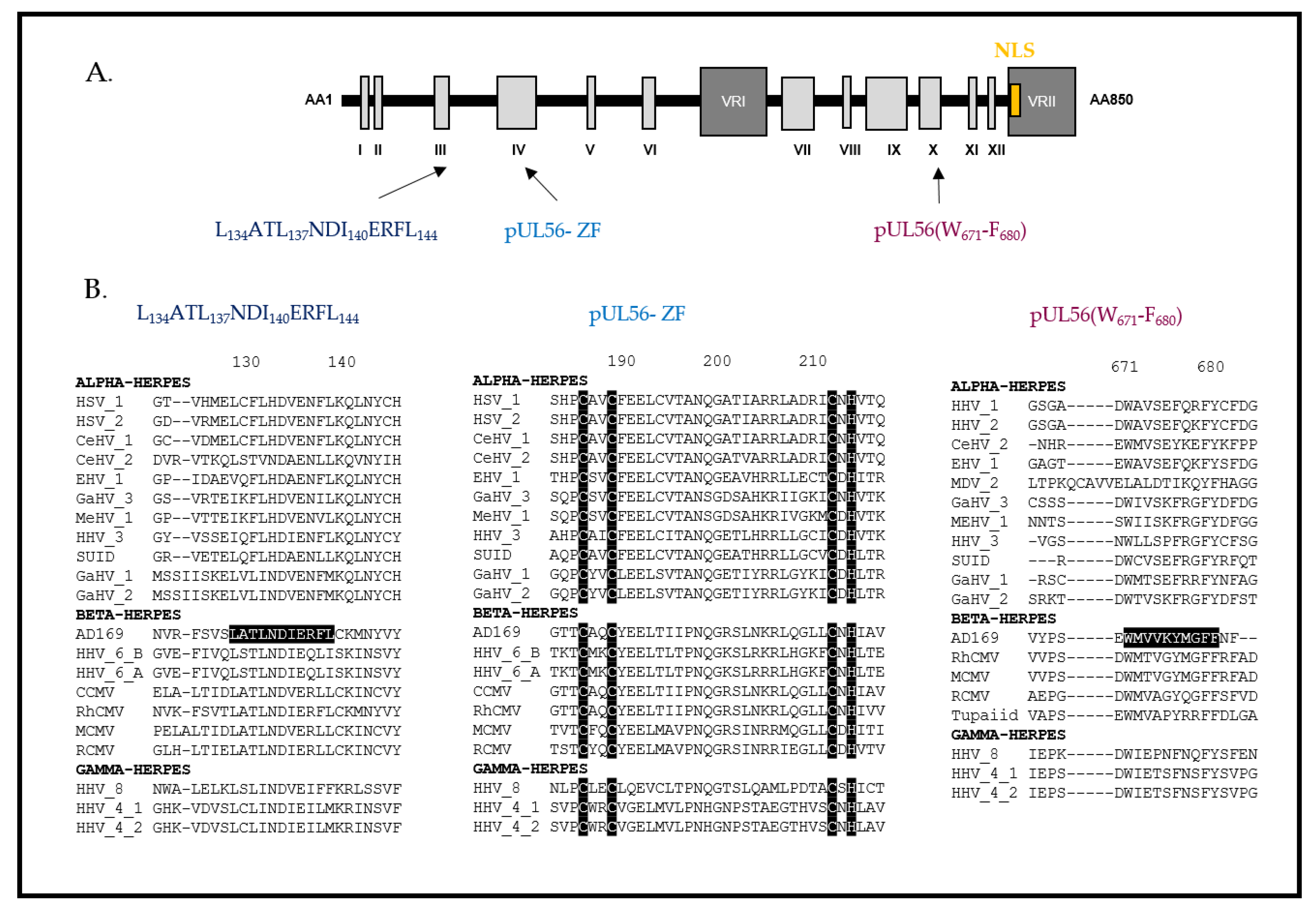
- Champier, G.; Couvreux, A.; Hantz, S.; Rametti, A.; Mazeron, M.-C.; Bouaziz, S.; Denis, F.; Alain, S. Putative Functional Domains of Human Cytomegalovirus PUL56 Involved in Dimerization and Benzimidazole D-Ribonucleoside Activity. Antivir. Ther. 2008, 13, 643–654. [Google Scholar]
- Bogner, E.; Reschke, M.; Reis, B.; Mockenhaupt, T.; Radsak, K. Identification of the Gene Product Encoded by ORF UL56 of the Human Cytomegalovirus Genome. Virology 1993, 196, 290–293. [Google Scholar] [CrossRef]
- Addison, C.; Rixon, F.J.; Preston, V.G. Herpes Simplex Virus Type 1 UL28 Gene Product Is Important for the Formation of Mature Capsids. J. Gen. Virol. 1990, 71 Pt 10, 2377–2384. [Google Scholar] [CrossRef]
- Giesen, K.; Radsak, K.; Bogner, E. The Potential Terminase Subunit of Human Cytomegalovirus, PUL56, Is Translocated into the Nucleus by Its Own Nuclear Localization Signal and Interacts with Importin α. J. Gen. Virol. 2000, 81, 2231–2244. [Google Scholar] [CrossRef]
- Adelman, K.; Salmon, B.; Baines, J.D. Herpes Simplex Virus DNA Packaging Sequences Adopt Novel Structures That Are Specifically Recognized by a Component of the Cleavage and Packaging Machinery. Proc. Natl. Acad. Sci. USA 2001, 98, 3086–3091. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.-S.; Bogner, E. ATPase Activity of the Terminase Subunit PUL56 of Human Cytomegalovirus. J. Biol. Chem. 2002, 277, 6943–6948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholz, B.; Rechter, S.; Drach, J.C.; Townsend, L.B.; Bogner, E. Identification of the ATP-Binding Site in the Terminase Subunit PUL56 of Human Cytomegalovirus. Nucleic Acids Res. 2003, 31, 1426–1433. [Google Scholar] [CrossRef] [Green Version]
- Thoma, C.; Borst, E.; Messerle, M.; Rieger, M.; Hwang, J.-S.; Bogner, E. Identification of the Interaction Domain of the Small Terminase Subunit PUL89 with the Large Subunit PUL56 of Human Cytomegalovirus. Biochemistry 2006, 45, 8855–8863. [Google Scholar] [CrossRef]
- Ligat, G.; Jacquet, C.; Chou, S.; Couvreux, A.; Alain, S.; Hantz, S. Identification of a Short Sequence in the HCMV Terminase PUL56 Essential for Interaction with PUL89 Subunit. Sci. Rep. 2017, 7, 8796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ligat, G.; Couvreux, A.; Cazal, R.; Alain, S.; Hantz, S. Highlighting of a LAGLIDADG and a Zing Finger Motifs Located in the PUL56 Sequence Crucial for HCMV Replication. Viruses 2019, 11, 1093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Champier, G.; Hantz, S.; Couvreux, A.; Stuppfler, S.; Mazeron, M.-C.; Bouaziz, S.; Denis, F.; Alain, S. New Functional Domains of Human Cytomegalovirus PUL89 Predicted by Sequence Analysis and Three-Dimensional Modelling of the Catalytic Site DEXDc. Antivir. Ther. 2007, 12, 217–232. [Google Scholar]
- Leffers, G.; Rao, V.B. Biochemical Characterization of an ATPase Activity Associated with the Large Packaging Subunit Gp17 from Bacteriophage T4. J. Biol. Chem. 2000, 275, 37127–37136. [Google Scholar] [CrossRef] [Green Version]
- Scheffczik, H. The Terminase Subunits PUL56 and PUL89 of Human Cytomegalovirus Are DNA-Metabolizing Proteins with Toroidal Structure. Nucleic Acids Res. 2002, 30, 1695–1703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankhala, R.S.; Lokareddy, R.K.; Cingolani, G. Divergent Evolution of Nuclear Localization Signal Sequences in Herpesvirus Terminase Subunits. J. Biol. Chem. 2016, 291, 11420–11433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuber, S.; Wagner, K.; Goldne, T.; Lischka, P.; Steinbrueck, L.; Messerle, M.; Borst, E.M. Mutual Interplay between the Human Cytomegalovirus Terminase Subunits PUL51, PUL56, and PUL89 Promotes Terminase Complex Formation. J. Virol. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neuber, S.; Wagner, K.; Messerle, M.; Borst, E.M. The C-Terminal Part of the Human Cytomegalovirus Terminase Subunit PUL51 Is Central for Terminase Complex Assembly. J. Gen. Virol. 2017. [Google Scholar] [CrossRef] [Green Version]
- Albright, B.S.; Kosinski, A.; Szczepaniak, R.; Cook, E.A.; Stow, N.D.; Conway, J.F.; Weller, S.K. The Putative Herpes Simplex Virus 1 Chaperone Protein UL32 Modulates Disulfide Bond Formation during Infection. J. Virol. 2015, 89, 443–453. [Google Scholar] [CrossRef] [Green Version]
- Meissner, C.S.; Köppen-Rung, P.; Dittmer, A.; Lapp, S.; Bogner, E. A “Coiled-Coil” Motif Is Important for Oligomerization and DNA Binding Properties of Human Cytomegalovirus Protein UL77. PLoS ONE 2011, 6, e25115. [Google Scholar] [CrossRef] [PubMed]
- Meissner, C.S.; Suffner, S.; Schauflinger, M.; von Einem, J.; Bogner, E. A Leucine Zipper Motif of a Tegument Protein Triggers Final Envelopment of Human Cytomegalovirus. J. Virol. 2012, 86, 3370–3382. [Google Scholar] [CrossRef] [Green Version]
- Cockrell, S.K.; Huffman, J.B.; Toropova, K.; Conway, J.F.; Homa, F.L. Residues of the UL25 Protein of Herpes Simplex Virus That Are Required for Its Stable Interaction with Capsids. J. Virol. 2011, 85, 4875–4887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeRussy, B.M.; Boland, M.T.; Tandon, R. Human Cytomegalovirus PUL93 Links Nucleocapsid Maturation and Nuclear Egress. J. Virol. 2016. [Google Scholar] [CrossRef] [Green Version]
- Huet, A.; Huffman, J.B.; Conway, J.F.; Homa, F.L. Role of the Herpes Simplex Virus CVSC Proteins at the Capsid Portal Vertex. J. Virol. 2020, 94, e01534-20. [Google Scholar] [CrossRef] [PubMed]
- Heming, J.D.; Conway, J.F.; Homa, F.L. Herpesvirus Capsid Assembly and DNA Packaging. Adv. Anat. Embryol. Cell Biol. 2017, 223, 119–142. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, W.; Li, Z.; Kumar, V.; Alvarez-Cabrera, A.L.; Leibovitch, E.C.; Cui, Y.; Mei, Y.; Bi, G.-Q.; Jacobson, S.; et al. Atomic Structure of the Human Herpesvirus 6B Capsid and Capsid-Associated Tegument Complexes. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Tandon, R.; Mocarski, E.S.; Conway, J.F. The A, B, Cs of Herpesvirus Capsids. Viruses 2015, 7, 899–914. [Google Scholar] [CrossRef]
- Townsend, L.B.; Turk, S.R.; Nassiri, M.R.; Drach, J.C. Design, Synthesis, and Antiviral Activity of Certain 2,5,6-Trihalo-1-(.beta.-D-ribofuranosyl) benzimidazoles. J. Med. Chem. 1995, 38, 4098–4105. [Google Scholar] [CrossRef]
- Emery, V.C.; Hassan-Walker, A.F. Focus on New Drugs in Development Against Human Cytomegalovirus. Drugs 2002, 62, 1853–1858. [Google Scholar] [CrossRef]
- Underwood, M.R.; Ferris, R.G.; Selleseth, D.W.; Davis, M.G.; Drach, J.C.; Townsend, L.B.; Biron, K.K.; Boyd, F.L. Mechanism of Action of the Ribopyranoside Benzimidazole GW275175X against Human Cytomegalovirus. Antimicrob. Agents Chemother. 2004, 48, 1647–1651. [Google Scholar] [CrossRef] [Green Version]
- Reefschlaeger, J. Novel Non-Nucleoside Inhibitors of Cytomegaloviruses (BAY 38-4766): In Vitro and in Vivo Antiviral Activity and Mechanism of Action. J. Antimicrob. Chemother. 2001, 48, 757–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kornfeind, E.M.; Visalli, R.J. Human Herpesvirus Portal Proteins: Structure, Function, and Antiviral Prospects. Rev. Med. Virol. 2018, 28, e1972. [Google Scholar] [CrossRef] [PubMed]
- Chemaly, R.F.; Ullmann, A.J.; Stoelben, S.; Richard, M.P.; Bornhäuser, M.; Groth, C.; Einsele, H.; Silverman, M.; Mullane, K.M.; Brown, J.; et al. Letermovir for Cytomegalovirus Prophylaxis in Hematopoietic-Cell Transplantation. N. Engl. J. Med. 2014, 370, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Marty, F.M.; Ljungman, P.; Chemaly, R.F.; Maertens, J.; Dadwal, S.S.; Duarte, R.F.; Haider, S.; Ullmann, A.J.; Katayama, Y.; Brown, J.; et al. Letermovir Prophylaxis for Cytomegalovirus in Hematopoietic-Cell Transplantation. N. Engl. J. Med. 2017. [Google Scholar] [CrossRef] [PubMed]
- Goldner, T.; Hempel, C.; Ruebsamen-Schaeff, H.; Zimmermann, H.; Lischka, P. Geno- and Phenotypic Characterization of Human Cytomegalovirus Mutants Selected in Vitro after Letermovir (AIC246) Exposure. Antimicrob. Agents Chemother. 2014, 58, 610–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, S. Rapid in Vitro Evolution of Human Cytomegalovirus UL56 Mutations That Confer Letermovir Resistance. Antimicrob. Agents Chemother. 2015. [Google Scholar] [CrossRef] [Green Version]
- Chou, S. A Third Component of the Human Cytomegalovirus Terminase Complex Is Involved in Letermovir Resistance. Antiviral Res. 2017, 148, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Chou, S. Comparison of Cytomegalovirus Terminase Gene Mutations Selected after Exposure to Three Distinct Inhibitor Compounds. Antimicrob. Agents Chemother. 2017. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
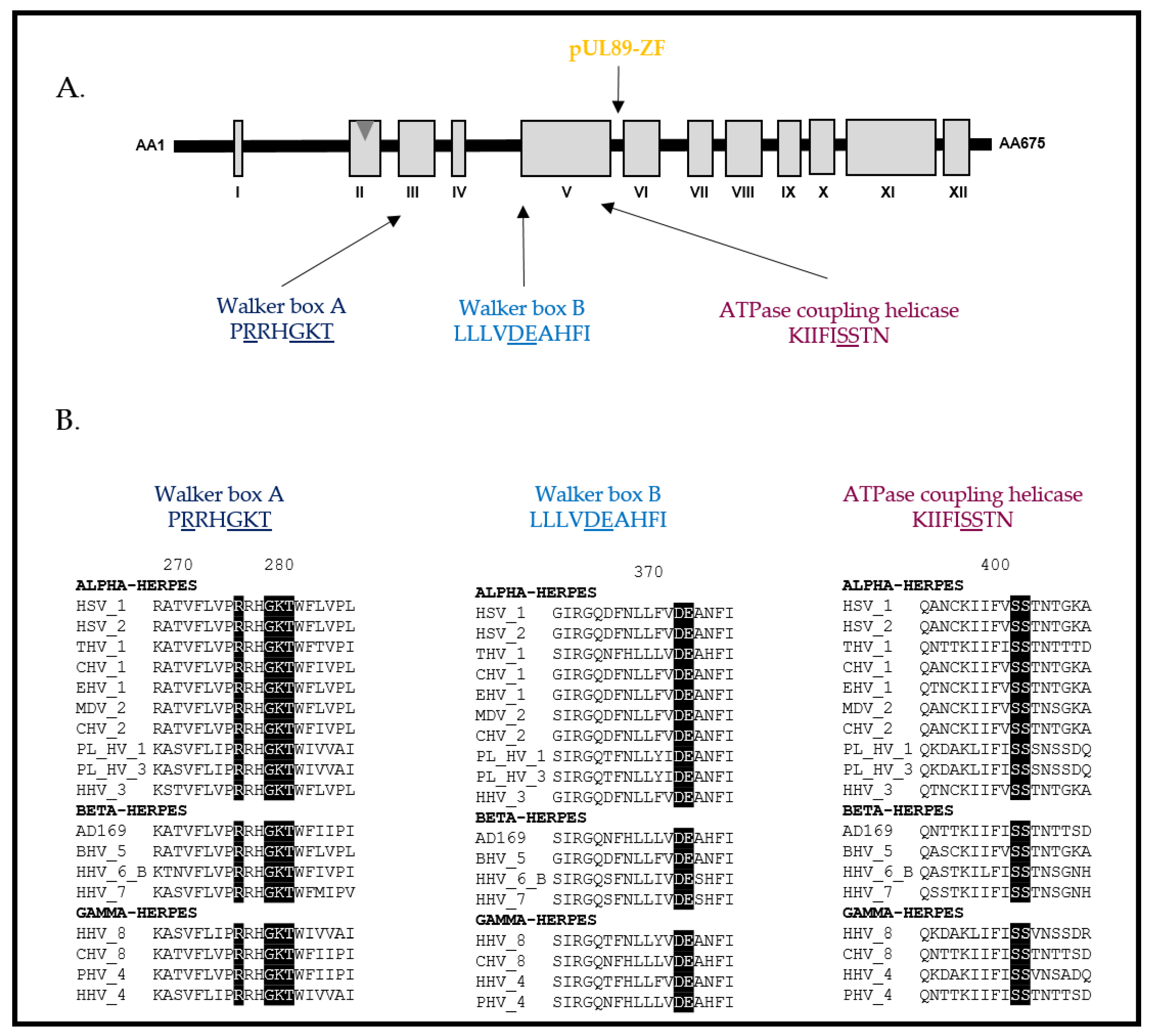{kind=link}
| α Herpesviruses | β Herpesviruses | γ Herpesviruses | Notes | ||||
|---|---|---|---|---|---|---|---|
| HSV-1 | VZV | HCMV | HHV-6 | HHV-8 | EBV | ||
| Genes encoding capsid proteins | UL19 | ORF40 | UL86 | U57 | ORF25 | BcLF1 | MCP: component of hexons and pentons |
| UL18 | ORF41 | UL85 | U56 | ORF26 | BDLF1 | Triplex dimer linking hexons and pentons together | |
| UL38 | ORF20 | UL46 | U29 | ORF62 | BORF1 | Triplex monomer linking hexons and pentons together | |
| UL35 | ORF23 | UL48a | U32 | ORF65 | BFRF3 | SCP: located on tips of hexons | |
| UL26 | ORF33 | UL80 | U53 | ORF17 | BVRF2 | Maturational protease: generates mature forms of scaffolding proteins | |
| UL26.5 | ORF33.5 | UL80.5 | U53.5 | ORF17.5 | BdRF1 | Scaffolding protein removed from capsid during DNA packaging | |
| UL6 | ORF54 | UL104 | U76 | ORF43 | BBRF1 | Portal protein: forms a dodecametic ring at one of the twelve capsid vertexes, complexed with terminase subunit | |
| Genes encoding DNA-packaging proteins | UL28 | ORF30 | UL56 | U40 | ORF7 | BALF3 | Subunit of terminase: “pac” site-specific binding, capsid-associated |
| UL15 | ORF42/45 | UL89 | U66 | ORF29 | BGRF1/BDRF1 | Subunit of terminase: ATPase subunit, putative nuclease domain | |
| UL33 | ORF25 | UL51 | U35 | ORF67A | BFRF1A | Interacts with terminase and mediates their correct subnuclear localization | |
| UL32 | ORF26 | UL52 | U36 | ORF68 | BFLF1 | Putative protein involved in capsid localization in the nucleus | |
| UL25 | ORF34 | UL77 | U50 | ORF19 | BVRF1 | Forms an asymmetric dimer and interacts with pUL17 and the nuclear egress complex | |
| UL17 | ORF43 | UL93 | U64 | ORF32 | BGLF1 | Anchors terminase to the capsid and is involved in determining the site of the first cleavage reaction | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muller, C.; Alain, S.; Baumert, T.F.; Ligat, G.; Hantz, S. Structures and Divergent Mechanisms in Capsid Maturation and Stabilization Following Genome Packaging of Human Cytomegalovirus and Herpesviruses. Life 2021, 11, 150. https://0-doi-org.brum.beds.ac.uk/10.3390/life11020150
Muller C, Alain S, Baumert TF, Ligat G, Hantz S. Structures and Divergent Mechanisms in Capsid Maturation and Stabilization Following Genome Packaging of Human Cytomegalovirus and Herpesviruses. Life. 2021; 11(2):150. https://0-doi-org.brum.beds.ac.uk/10.3390/life11020150
Chicago/Turabian StyleMuller, Clotilde, Sophie Alain, Thomas F. Baumert, Gaëtan Ligat, and Sébastien Hantz. 2021. "Structures and Divergent Mechanisms in Capsid Maturation and Stabilization Following Genome Packaging of Human Cytomegalovirus and Herpesviruses" Life 11, no. 2: 150. https://0-doi-org.brum.beds.ac.uk/10.3390/life11020150