
Cell Populations Expressing Stemness-Associated Markers in Lung Adenocarcinoma

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lung Adenocarcinoma Tissue Samples
2.2. Lung Adenocarcinoma-Derived Primary Cell Lines
2.3. Histochemical and Immunohistochemical Staining
2.4. Image Capture and Analysis
2.5. In Situ Hybridization
2.6. Cell Counting and Statistical Analysis
2.7. RT-qPCR and Statistical Analysis
2.8. Western Blotting
2.9. In Vitro Initial Tumorsphere Formation Assays
3. Results
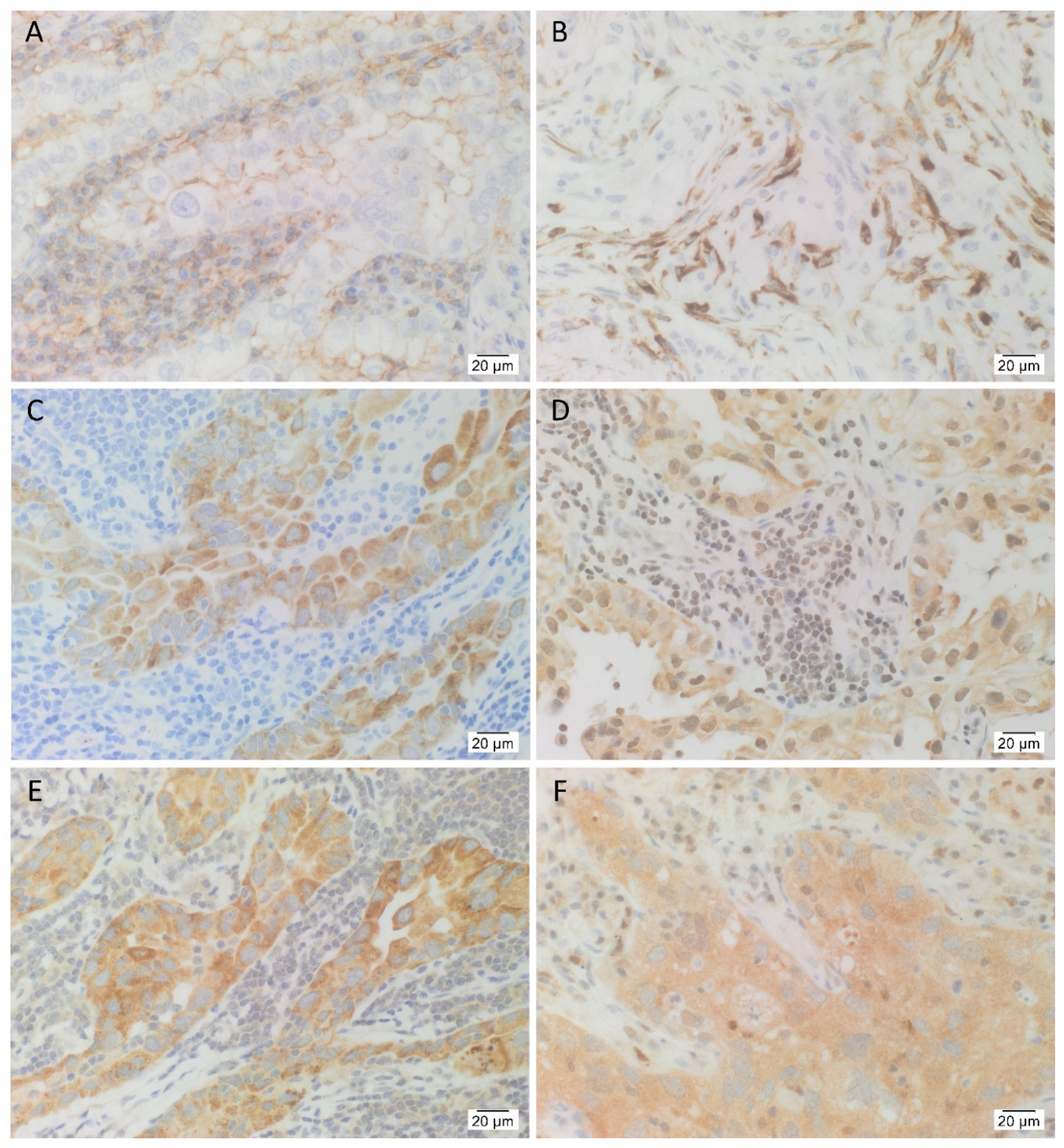
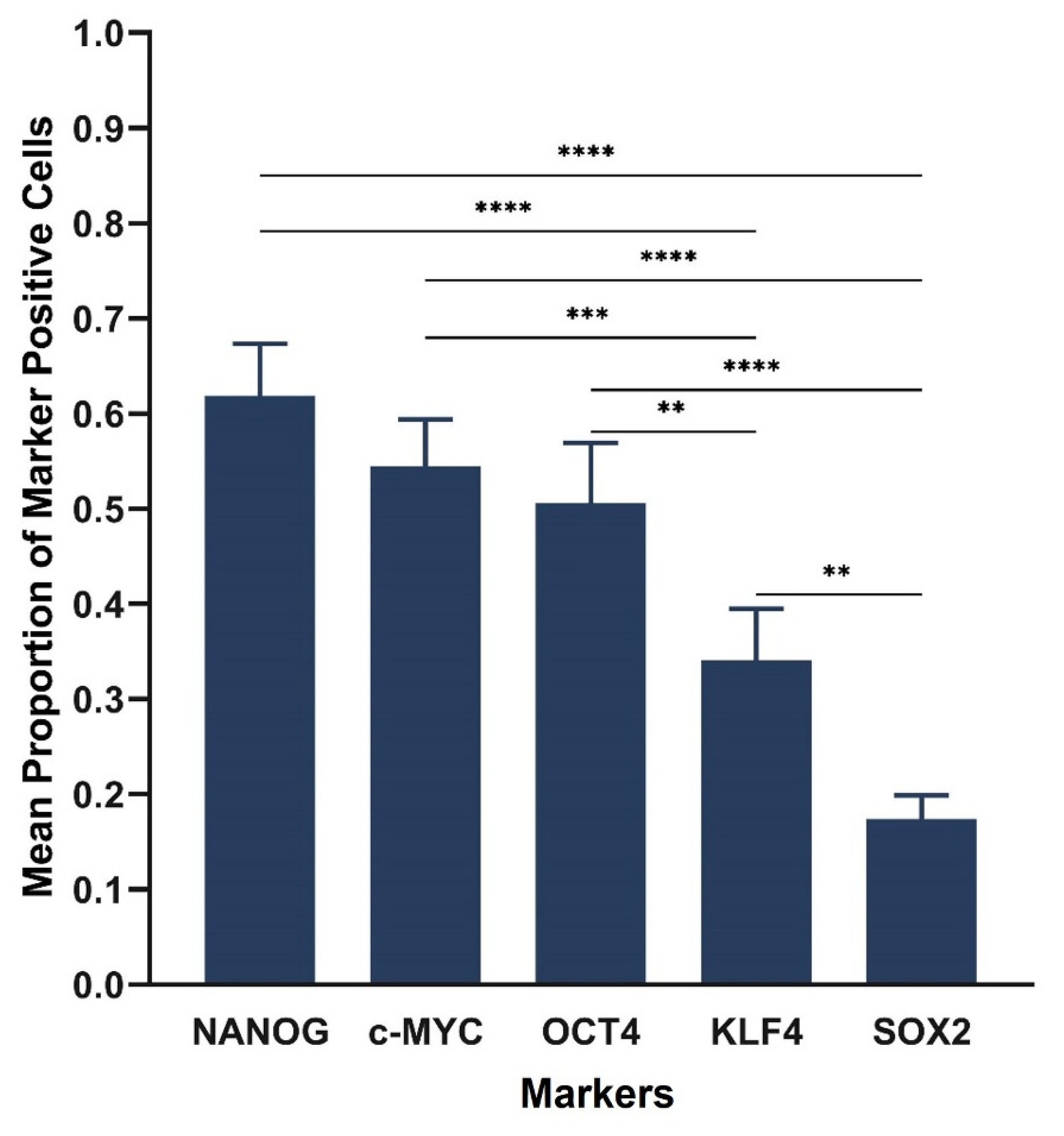
3.1. IHC Staining Demonstrated the Presence the Cancer Stem Cell Marker CD44, and the Five Stemness-Associated Markers in Lung Adenocarcinoma Tissues
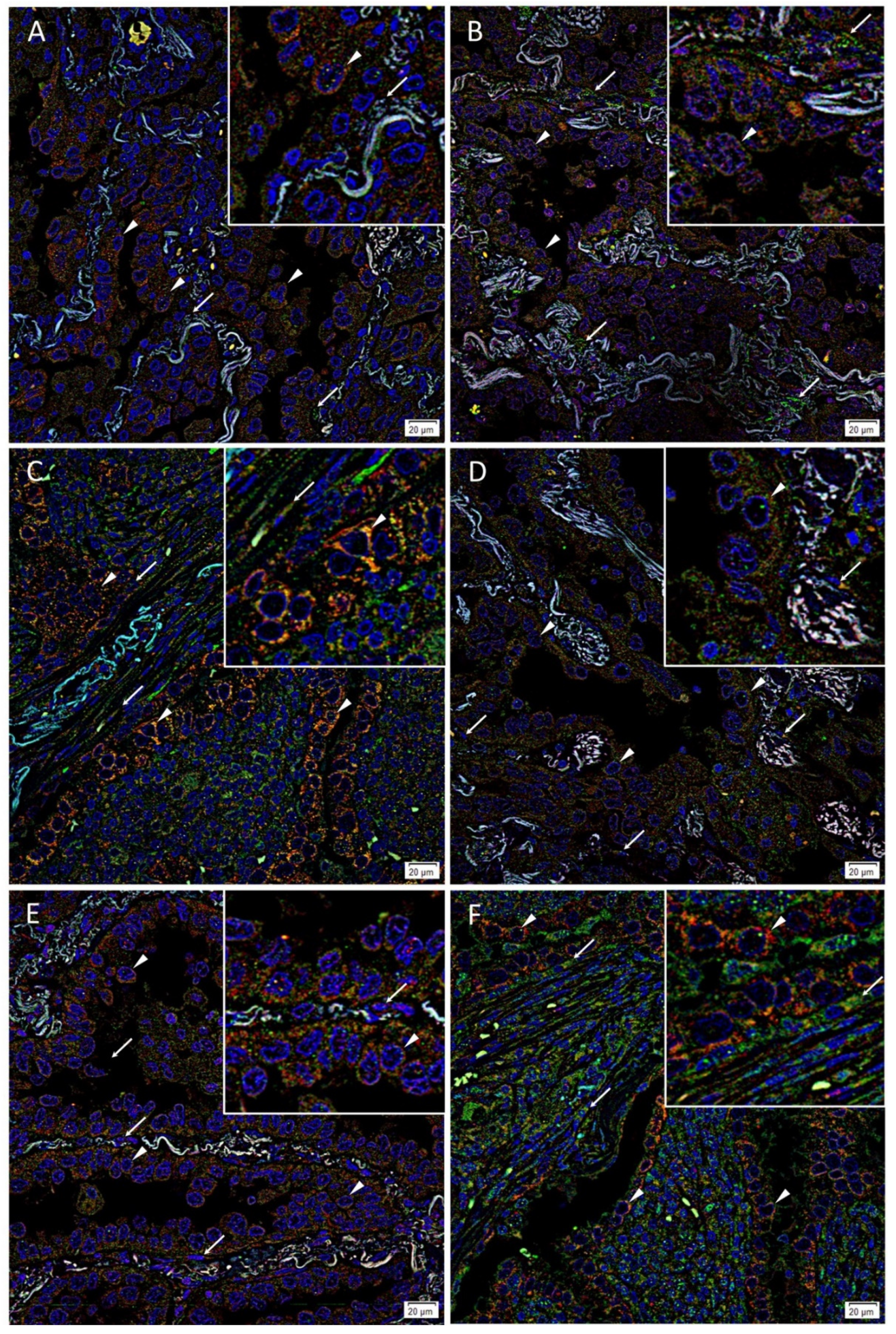
3.2. Immunofluorescence Staining Showed Co-Expression of OCT4 and c-MYC with NANOG, SOX2 and KLF4 by Tumor Gland Cells, and Exclusive Expression of OCT4 and c-MYC by Stromal Cells in Lung Adenocarcinoma Tissues
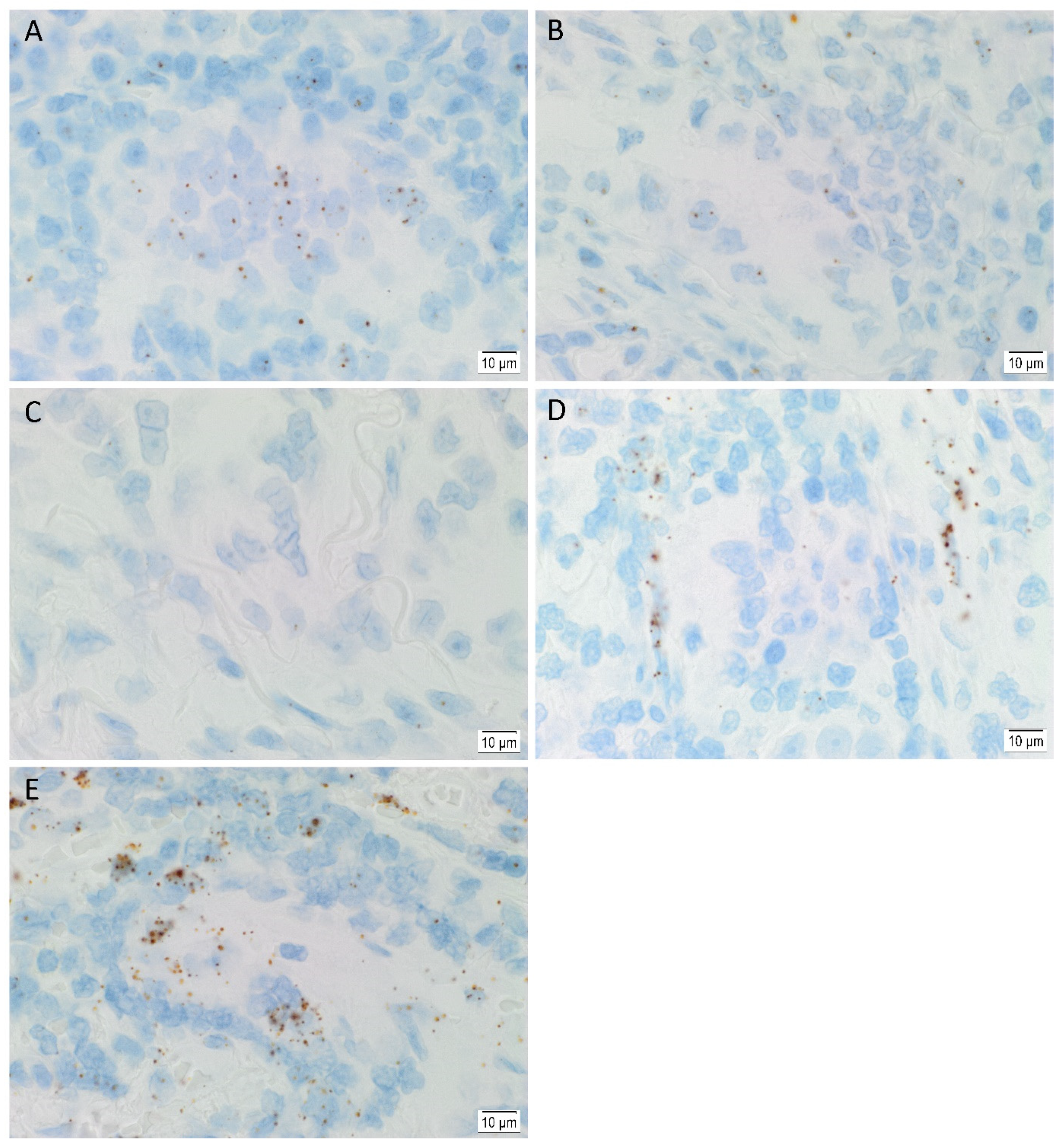
3.3. In Situ Hybridization Showed mRNA Expression of All Five Stemness-Associated Markers in Lung Adenocarcinoma Tissues
3.4. RT-qPCR Demonstrated mRNA Expression of All Stemness-Associated Markers, except for SOX2 in Lung Adenocarcinoma-Derived Primary Cell Lines
3.5. Western Blotting Demonstrated the Presence of All Five Stemness-Associated Markers, except for NANOG in Lung Adenocarcinoma-Derived Primary Cell Lines
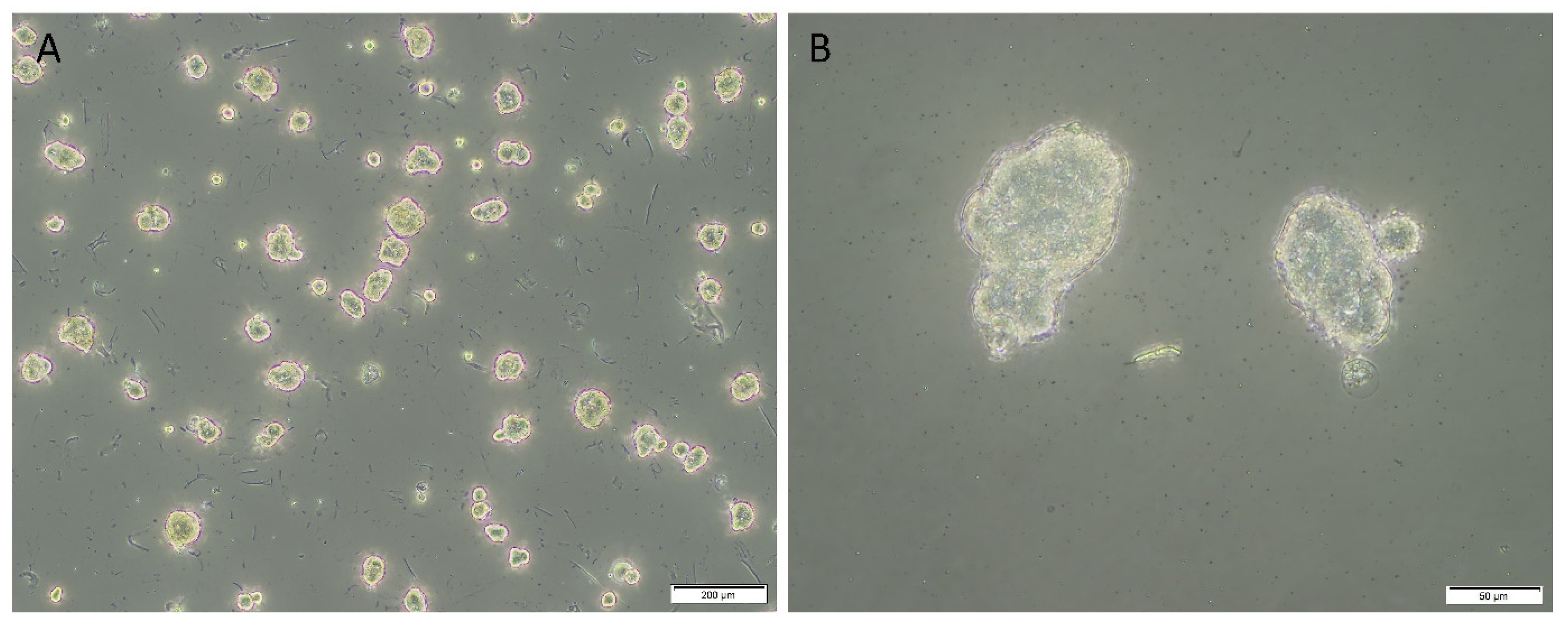
3.6. In Vitro Initial Tumorsphere Formation Assays Demonstrated Evidence of Tumorspheres across Four Lung Adenocarcinoma-Derived Primary Cell Lines
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Patents
References
- Bray, F.; Me, J.F.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Zappa, C.; Mousa, S.A. Non-small cell lung cancer: Current treatment and future advances. Transl. Lung Cancer Res. 2016, 5, 288–300. [Google Scholar] [CrossRef] [Green Version]
- Travis, W.D.; Brambilla, E.; Nicholson, A.G.; Yatabe, Y.; Austin, J.H.; Beasley, M.B.; Chirieac, L.R.; Dacic, S.; Duhig, E.; Flieder, D.B.; et al. The 2015 World Health Organization Classification of lung tumors: Impact of genetic, clinical and radiologic advances since the 2004 classification. J. Thorac. Oncol. 2015, 10, 1243–1260. [Google Scholar] [CrossRef] [Green Version]
- Wakelee, H.A.; Chang, E.T.; Gomez, S.L.; Keegan, T.H.M.; Feskanich, D.; Clarke, C.A.; Holmberg, L.; Yong, L.C.; Kolonel, L.N.; Gould, M.K.; et al. Lung cancer incidence in never smokers. J. Clin. Oncol. 2007, 25, 472–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gazdar, A.F.; Minna, J.D. Cigarettes, sex, and lung adenocarcinoma. J. Nat. 1997, 89, 1563–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alamgeer, M.; Peacock, C.D.; Matsui, W.; Ganju, V.; Watkins, D.N. Cancer stem cells in lung cancer: Evidence and controversies. Respirology 2013, 18, 757–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, E.L.-H.; Fiscus, R.R.; Tung, J.W.; Tin, V.P.-C.; Cheng, L.C.; Sihoe, A.D.-L.; Fink, L.M.; Ma, Y.; Wong, M.P. Non-small cell lung cancer cells expressing CD44 are enriched for stem cell-like properties. PLoS ONE 2010, 5, e14062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterlacci, W.; Savic, S.; Fiegl, M.; Obermann, E.; Tzankov, A. Putative stem cell markers in non–small-cell lung cancer: A Clinicopathologic characterization. J. Thorac. Oncol. 2014, 9, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.-Y.; Huang, C.-S.; Yang, Y.-P.; Liu, C.-Y.; Liu, Y.-Y.; Wu, W.-W.; Lu, K.-H.; Chen, K.-H.; Chang, Y.-L.; Lee, S.-D.; et al. The subpopulation of CD44-positive cells promoted tumorigenicity and metastatic ability in lung adenocarcinoma. J. Chin. Med. Assoc. 2019, 82, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Gao, Y.; Cui, Y.; Zhang, T.; Cui, R.; Jiang, Y.; Shi, J. Overexpression of CD44 is associated with the occurrence and migration of non-small cell lung cancer. Mol. Med. Rep. 2016, 14, 3159–3167. [Google Scholar] [CrossRef] [Green Version]
- Eramo, A.; Lotti, F.; Sette, G.; Pilozzi, E.; Biffoni, M.; Di Virgilio, A.; Conticello, C.; Ruco, L.; Peschle, C.; De Maria, R. Identification and expansion of the tumorigenic lung cancer stem cell population. Cell Death Differ. 2007, 15, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-C.; Hsu, H.-S.; Chen, Y.-W.; Tsai, T.-H.; How, C.-K.; Wang, C.-Y.; Hung, S.-C.; Chang, Y.-L.; Tsai, M.-L.; Lee, Y.-Y.; et al. Oct-4 expression maintained cancer stem-like properties in lung cancer-derived CD133-positive cells. PLoS ONE 2008, 3, e2637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertolini, G.; Roz, L.; Perego, P.; Tortoreto, M.; Fontanella, E.; Gatti, L.; Pratesi, G.; Fabbri, A.; Andriani, F.; Tinelli, S.; et al. Highly tumorigenic lung cancer CD133+ cells display stem-like features and are spared by cisplatin treatment. Proc. Natl. Acad. Sci. USA 2009, 106, 16281–16286. [Google Scholar] [CrossRef] [Green Version]
- Tirino, V.; Camerlingo, R.; Franco, R.; Malanga, D.; La Rocca, A.; Viglietto, G.; Rocco, G.; Pirozzi, G. The role of CD133 in the identification and characterisation of tumour-initiating cells in non-small-cell lung cancer. Eur. J. Cardio-Thoracic Surg. 2009, 36, 446–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, F.; Qiu, Q.; Khanna, A.; Todd, N.W.; Deepak, J.; Xing, L.; Wang, H.; Liu, Z.; Su, Y.; Stass, S.A.; et al. Aldehyde dehydrogenase 1 Is a tumor stem cell-associated marker in lung cancer. Mol. Cancer Res. 2009, 7, 330–338. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Walcher, L.; Kistenmacher, A.-K.; Suo, H.; Kitte, R.; Dluczek, S.; Strauß, A.; Blaudszun, A.-R.; Yevsa, T.; Fricke, S.; Kossatz-Boehlert, U. Cancer stem cells—Origins and biomarkers: Perspectives for targeted personalized therapies. Front. Immunol. 2020, 11, 1280. [Google Scholar] [CrossRef]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Zeineddine, D.; Hammoud, A.A.; Mortada, M.; Boeuf, H. The Oct4 protein: More than a magic stemness marker. Am. J. Stem Cells 2014, 3, 74–82. [Google Scholar]
- Zhang, W.C.S. Sox2, a key factor in the regulation of pluripotency and neural differentiation. World J. Stem Cells 2014, 6, 305–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fadous-Khalifé, M.C.; Aloulou, N.; Jalbout, M.; Hadchity, J.; Aftimos, G.; Paris, F.; Hadchity, E. Krüppel-like factor 4: A new potential biomarker of lung cancer. Mol. Clin. Oncol. 2016, 5, 35–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, D.M.; Thomas, S.D.; Islam, A.; Muench, D.; Sedoris, K. c-Myc and cancer metabolism. Clin. Cancer Res. 2012, 18, 5546–5553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Schaijik, B.; Davis, P.F.; Wickremesekera, A.C.; Tan, S.T.; Itinteang, T. Subcellular localisation of the stem cell markers OCT4, SOX2, NANOG, KLF4 and c-MYC in cancer: A review. J. Clin. Pathol. 2017, 71, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, J.; Xu, Z.; Ahmad, A.; Li, E.; Wang, Y.; Qin, S.; Wang, Q. Expression of Sox2 and Oct4 and their clinical significance in human non-small-cell lung cancer. Int. J. Mol. Sci. 2012, 13, 7663–7675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Yu, H.; Wang, X.; Zeng, J.; Li, D.; Lu, J.; Wang, C.; Wang, J.; Wei, J.; Jiang, M.; et al. Expression of seven stem-cell-associated markers in human airway biopsy specimens obtained via fiberoptic bronchoscopy. J. Exp. Clin. Cancer Res. 2013, 32, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-L.; Jia, L.-L.; Shi, M.-M.; Li, X.; Li, Z.-H.; Li, H.-F.; Wang, E.-H.; Jia, X.-S. Downregulation of KPNA2 in non-small-cell lung cancer is associated with Oct4 expression. J. Transl. Med. 2013, 11, 232. [Google Scholar] [CrossRef] [Green Version]
- Chiou, S.-H.; Wang, M.-L.; Chou, Y.-T.; Chen, C.-J.; Hong, C.-F.; Hsieh, W.-J.; Chang, H.-T.; Chen, Y.-S.; Lin, T.-W.; Hsu, H.-S.; et al. Coexpression of Oct4 and Nanog enhances malignancy in lung adenocarcinoma by inducing cancer stem cell–like properties and epithelial–mesenchymal transdifferentiation. Cancer Res. 2010, 70, 10433–10444. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Ma, C.; Wang, Z.; Liu, Z.; Liu, H.; Wang, T. Nanog, a novel prognostic marker for lung cancer. Surg. Oncol. 2013, 22, 224–229. [Google Scholar] [CrossRef]
- Li, X.-Q.; Yang, X.-L.; Zhang, G.; Wu, S.-P.; Deng, X.-B.; Xiao, S.-J.; Liu, Q.-Z.; Yao, K.-T.; Xiao, G.-H. Nuclear β-catenin accumulation is associated with increased expression of Nanog protein and predicts poor prognosis of non-small cell lung cancer. J. Transl. Med. 2013, 11, 114. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhang, J.; Kang, H.; Sun, G.; Wang, B.; Wang, Y.; Yang, M. Significance of stem cell marker Nanog gene in the diagnosis and prognosis of lung cancer. Oncol. Lett. 2016, 12, 2507–2510. [Google Scholar] [CrossRef] [Green Version]
- Chang, B.; Park, M.J.; Choi, S.I.; In, K.H.; Kim, C.H.; Lee, S.H. NANOG as an adverse predictive marker in advanced non–small cell lung cancer treated with platinum-based chemotherapy. Onco Targets Ther. 2017, 10, 4625–4633. [Google Scholar] [CrossRef] [Green Version]
- Nakatsugawa, M.; Takahashi, A.; Hirohashi, Y.; Torigoe, T.; Inoda, S.; Murase, M.; Asanuma, H.; Tamura, Y.; Morita, R.; Michifuri, Y.; et al. SOX2 is overexpressed in stem-like cells of human lung adenocarcinoma and augments the tumorigenicity. Lab. Investig. 2011, 91, 1796–1804. [Google Scholar] [CrossRef]
- Xiang, R.; Liao, D.; Cheng, T.; Zhou, H.; Shi, Q.; Chuang, T.S.; Markowitz, D.; A Reisfeld, R.; Luo, Y. Downregulation of transcription factor SOX2 in cancer stem cells suppresses growth and metastasis of lung cancer. Br. J. Cancer 2011, 104, 1410–1417. [Google Scholar] [CrossRef]
- Chen, S.; Xu, Y.; Chen, Y.; Li, X.; Mou, W.; Wang, L.; Liu, Y.; Reisfeld, R.A.; Xiang, R.; Lv, D.; et al. SOX2 gene regulates the transcriptional network of oncogenes and affects tumorigenesis of human lung cancer cells. PLoS ONE 2012, 7, e36326. [Google Scholar] [CrossRef] [Green Version]
- Sholl, L.M.; Barletta, J.A.; Yeap, B.Y.; Chirieac, L.R.; Hornick, J.L. Sox2 protein expression is an independent poor prognostic indicator in stage I lung adenocarcinoma. Am. J. Surg. Pathol. 2010, 34, 1193–1198. [Google Scholar] [CrossRef]
- Gomez-Casal, R.; Bhattacharya, C.; Ganesh, N.; Bailey, L.; Basse, P.; Gibson, M.; Epperly, M.; Levina, V. Non-small cell lung cancer cells survived ionizing radiation treatment display cancer stem cell and epithelial-mesenchymal transition phenotypes. Mol. Cancer 2013, 12, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, W.; Hofstetter, W.L.; Li, H.; Zhou, Y.; He, Y.; Pataer, A.; Wang, L.; Xie, K.; Swisher, S.G.; Fang, B. putative tumor-suppressive function of Krüppel-Like Factor 4 in primary lung carcinoma. Clin. Cancer Res. 2009, 15, 5688–5695. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Huang, L.; Gu, J.; Wu, J.; Ou, W.; Feng, J.; Liu, B.; Cui, X.; Zhou, Y. Restoration of KLF4 inhibits invasion and metastases of lung adenocarcinoma through suppressing MMP2. J. Cancer 2017, 8, 3480–3489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Li, X.; Peng, K.-Z.; Gao, T.; Cui, Y.; Ma, N.; Zhou, Y.; Hou, G. Subcellular localization of Klf4 in non-small cell lung cancer and its clinical significance. Biomed. Pharmacother. 2018, 99, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Sławek, S.; Szmyt, K.; Fularz, M.; Dziudzia, J.; Boruczkowski, M.; Sikora, J.; Kaczmarek, M. Pluripotency transcription factors in lung cancer—A review. Tumor Biol. 2015, 37, 4241–4249. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.M.S.; Itinteang, T.; A Chudakova, D.; Dunne, J.C.; Marsh, R.; Brasch, H.D.; Davis, P.F.; Tan, S.T. Characterisation of lymphocyte subpopulations in infantile haemangioma. J. Clin. Pathol. 2015, 68, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.; Brasch, H.D.; Van Schaijik, B.; Armstrong, J.R.; Marsh, R.W.; Davis, P.F.; Tan, S.T.; Itinteang, T. Expression and localization of cathepsins B, D, and G in Dupuytren’s disease. Plast. Reconstr. Surg. Glob. Open 2018, 6, e1686. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-H.; Yu, C.-C.; Wang, B.-Y.; Chang, W.-W. Tumorsphere as an effective in vitro platform for screening anti-cancer stem cell drugs. Oncotarget 2015, 7, 1215–1226. [Google Scholar] [CrossRef] [Green Version]
- Ebradshaw, A.; Ewickremesekera, A.; Tan, S.T.; Epeng, L.; Davis, P.F.; Itinteang, T. Cancer stem cell hierarchy in glioblastoma multiforme. Front. Surg. 2016, 3, 21. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.H.; Featherston, T.; Tan, S.T.; Chibnall, A.M.; Brasch, H.D.; Davis, P.F.; Itinteang, T. Characterization of cancer stem cells in moderately differentiated buccal mucosal squamous cell carcinoma. Front. Surg. 2016, 3, 46. [Google Scholar] [CrossRef]
- Pomares, A.H.; De-Maya-Girones, J.D.; Calabuig-Fariñas, S.; Lucas-Dominguez, R.; Martínez, A.; Pardo-Sánchez, J.M.; Alonso, S.; Blasco, A.; Guijarro, R.; Martorell, M.; et al. Lung tumorspheres reveal cancer stem cell-like properties and a score with prognostic impact in resected non-small-cell lung cancer. Cell Death Dis. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Bradshaw, A.; Wickremesekera, A.; Brasch, H.D.; Chibnall, A.M.; Davis, P.F.; Tan, S.T.; Itinteang, T. Cancer stem cells in glioblastoma multiforme. Front. Surg. 2016, 3, 48. [Google Scholar] [CrossRef] [Green Version]
- Roth, I.M.; Wickremesekera, A.C.; Wickremesekera, S.K.; Davis, P.F.; Tan, S.T. Therapeutic targeting of cancer stem cells via modulation of the renin-angiotensin system. Front. Oncol. 2019, 9, 745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, D.C.H.; Roth, I.M.; Wickremesekera, A.C.; Davis, P.F.; Kaye, A.H.; Mantamadiotis, T.; Stylli, S.S.; Tan, S.T. Therapeutic targeting of cancer stem cells in human glioblastoma by manipulating the renin-angiotensin system. Cells 2019, 8, 1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Initial Tumorsphere Size (µm) * | % of Initial Tumorspheres > 50 µm | Days for Initial Signs of Dark Centers | ||
|---|---|---|---|---|---|
| Mean | Median | Range | |||
| 1 | 63.81 | 66.19 | 43.52–85.28 | 80 | 3 |
| 2 | 29.06 | 27.01 | 20.91–43.02 | 0 | ns ** |
| 3 | 49.71 | 40.12 | 29.12–122.05 | 17.6 | 10 |
| 4 | 38.90 | 38.07 | 26.78–60.95 | 13 | ns ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paterson, C.; Kilmister, E.J.; Brasch, H.D.; Bockett, N.; Patel, J.; Paterson, E.; Purdie, G.; Galvin, S.; Davis, P.F.; Itinteang, T.; et al. Cell Populations Expressing Stemness-Associated Markers in Lung Adenocarcinoma. Life 2021, 11, 1106. https://0-doi-org.brum.beds.ac.uk/10.3390/life11101106
Paterson C, Kilmister EJ, Brasch HD, Bockett N, Patel J, Paterson E, Purdie G, Galvin S, Davis PF, Itinteang T, et al. Cell Populations Expressing Stemness-Associated Markers in Lung Adenocarcinoma. Life. 2021; 11(10):1106. https://0-doi-org.brum.beds.ac.uk/10.3390/life11101106
Chicago/Turabian StylePaterson, Claudia, Ethan J. Kilmister, Helen D. Brasch, Nicholas Bockett, Josie Patel, Erin Paterson, Gordon Purdie, Sean Galvin, Paul F. Davis, Tinte Itinteang, and et al. 2021. "Cell Populations Expressing Stemness-Associated Markers in Lung Adenocarcinoma" Life 11, no. 10: 1106. https://0-doi-org.brum.beds.ac.uk/10.3390/life11101106