How Diet-Induced Changes in the “Gut-Liver” Axis Affect Chronic Liver Disease Outcome?


1
Department of Nutritional Sciences and Dietetics, International Hellenic University (IHU), 57400 Thessaloniki, Greece
2
Department of Biology, School of Medicine, National and Kapodistrian University of Athens, 157 72 Athens, Greece
3
Center for New Biotechnologies and Precision Medicine, School of Medicine, National and Kapodistrian University of Athens, 106 79 Athens, Greece
4
Center of Basic Research, Biomedical Research Foundation of the Academy of Athens, 115 27 Athens, Greece
*
Author to whom correspondence should be addressed.
Livers 2021, 1(1), 40-48; https://0-doi-org.brum.beds.ac.uk/10.3390/livers1010004
Submission received: 22 December 2020
/
Revised: 12 February 2021
/
Accepted: 15 February 2021
/
Published: 23 February 2021
Abstract
:Hepatocellular carcinoma (HCC) occurs in patients with chronic liver damage, inflammation and cirrhosis. The facilitators involved in increasing the HCC risk in the damaged liver are yet to be discovered. Diet and lifestyle have a profound effect on the liver inflammation and HCC. The term “gut liver axis” describes the bidirectional relationship between the liver and the gut, which are both anatomically and functionally related. Chronic liver damage is characterised by increased intestinal permeability that allows the translocation of various components and metabolites from the gut microbiota to the liver, resulting in liver inflammation and fibrosis. In this review, we discuss how diet-induced changes in gut microbiome composition, such as lipopolysaccharide and lipoteichoic acid, and its metabolites, such as bile acids, play a role in the pathogenesis of liver fibrosis and HCC.
1. Introduction
Hepatocellular carcinoma (HCC) is the most common primary cancer in adults representing third of all cancer-related deaths [1]. Patients with chronic liver disease (CLD), inflammation and cirrhosis have an 80% risk to develop HCC. The factors that facilitate HCC onset and development remain to be discovered. We do know from epidemiological and observational data that a healthy diet can offer protection for a number of different cancer kinds, whereas an unbalanced diet augments the risk for cancer. The majority of studies on the association of diet and HCC pathogenesis agree that there is a protective role of vegetables, fruit, cereals and fish in HCC risk [2,3].
Diet exerts a powerful influence on gut microbial composition that may impact liver disease. Humans are thought to have evolved a symbiotic relationship with gut microbiota; however, the molecular mechanisms underpinning this mutually benefiting relationship remain poorly understood. DNA-based analysis has enabled the identification of the microbes that reside in gut microbiome, but understanding their precise function is still a challenge. For example, gut microbiota produces low molecular weight substances, including metabolic derivatives that can regulate biochemical and molecular pathways.
The notion that cirrhosis positively correlates with intestinal dysbiosis was proposed by Sherlock and his team in the 1950s. They reported that there was an overload of coliform bacteria in the ileum of cirrhotic patients [4]. Today we know for a fact that chronic liver inflammation induces intestinal permeability followed by the translocation of gut bacteria, their components and/or metabolites, which are causing the characteristic complications of advanced liver disease stages [5,6].
In chronic liver inflammation, new-forming tumour cells and the resident liver cells and/or the tumour-forming cells are both regulated by inflammatory signals to survive. Due to the lack of comprehensive and prospective studies on chronic liver injury in humans, we have underestimated the contribution of inflammatory pathways induced from gut microbial components to HCC. The present review discusses how dietary patterns affect the liver-gut axis relationship through microbial alterations in the chronically inflamed liver and HCC development.
2. Diet Has a Direct Effect on the Gut-Liver Axis
An increasing body of evidence agrees that the dietary intake influences HCC outcome. A number of studies have reported an inverse correlation between milk [7,8], tea [9], fiber and whole grains [10], white meat [3,11], fruits and vegetables [12,13], and fish [14] and HCC progress. On the contrary, increased consumption of red meat [3,11] positively correlates with HCC risk; whereas studies on HCC risk and egg and rice consumption have reached conflicting results [9,15,16,17] (Table 1). Discrepancies between studies from different countries could be attributed to differences in the cultural dietary practices, for example, the type of oil used in cooking.
Animal meat provides a rich source of monounsaturated and saturated fatty acids and heme iron [3]. Red meat intake is known to increase the risk of HCC [3]. This association was anticipated because meat has high levels of carcinogens, such as N-nitroso compounds (NOC), heterocyclic amines (HCA) and the heme iron. During cooking, carcinogens are generated from iron metabolism together and reactive oxygen species are also releasing [3]. Iron catalysis to lipid peroxidation can cause hepatocyte and DNA damage in tissues [22]. Notably, there is a high HCC occurrence within patients with hemochromatosis, an iron overload hereditary disease [3]. Interestingly, polymorphisms in the N-acetyltransferase 2 (NAT2) gene, which is involved in the detoxification of carcinogenic aromatic amines, has been also shown to influence the association of red meat and liver cancer [24].
Evidence has shown that fish (lean and fatty), clams, and crustaceans play a protective role against HCC [3,23,25]. Fish is rich in n-3 polyunsaturated fatty acids (n-3 PUFA), which positively correlate with a low HCC risk, but have been shown to be inversely associated with high CLD risk [19,20,26]. In addition to fish, white meat contains high N3 PUFA levels and low levels of cholesterol and saturated fat levels [20]. N-3 PUFA contributes to HCC prevention because it has anti-inflammatory activities as it inhibits the production of IL-1 and TNF [27]. Further studies are required to identify the precise molecular pathways responsible for the chemoprotective role of n-3 PUFA on HCC development.
The association of fruit and vegetable consumption with HCC progression has been largely confirmed since Negri et al. and his team reported it in the 1990s [28]. High amounts of leafy vegetables in the diet, such as spinach and parsley, have been shown to offer protection against HCC [15]. Fruits and vegetables have high levels of minerals, vitamins, fibers, and flavonoids. Numerous studies demonstrated in vitro that flavonoids have anti-tumour effects, whereas studies using animal experiments showed that they do so by regulating pathways in metastasis, proliferation and angiogenesis [3,21]. Moreover, the protective role of tea in HCC development has been shown to the polyphenols acting as antioxidants, especially in the case of green tea [9].
Dietary intake of fiber from cereals inversely associates with HCC risk [3]. Fibers protect against HCC development, because they reduce appetite and energy input and therefore contribute directly to a balanced body weight and to the maintenance of postprandial glucose and circulating lipid levels [3]. Interestingly, a study using a rat experimental model showed that consumption of dry beans significantly increased the gene expression of six stress and toxicity-related hepatic genes (Cyp3a11, Cyp7a1, Fmo1, Gstm1, Mif and Ugt1a6) indicating that they play a protective role in HCC [29].
Dietary glycemic load (GL) measures the amount of food required to raise blood glucose levels, and it, therefore, represents the degree by which carbohydrate intake can surge blood glucose levels, i.e., the effect of the diet on glycemia. GL positively correlates with liver cancer independently from the presence of diabetes mellitus. The mechanisms involved are suggested to be through high insulin levels and resistance as well as glucose intolerance [2]. There is an urge to conduct intense research on the clinical features, dietary intake, microbiota composition and epidemiology of HCC in different populations.
3. The Gut-Liver Axis
Gut microbiome is crucial for the host’s survival as it metabolizes ingested food that could not be processed by the host otherwise. Bioinformatics, metabolomics and next-generation sequencing have led to the assessment of the intestinal composition by identifying the DNA sequences of the 16S ribosomal RNA gene and other microbial genes. In addition, the functional relevance of the intestinal microbial community shows that gut microbial components and their metabolites influence greatly human pathology [30].
Anatomically and functionally, the liver and the intestine are connected. The liver, however, is not directly in contact with gut bacteria, but it is the first organ where gut bacteria are translocating via the systemic circulation. Therefore, gut microbes and metabolites influence also organs distant from the intestine via portal circulation [30].
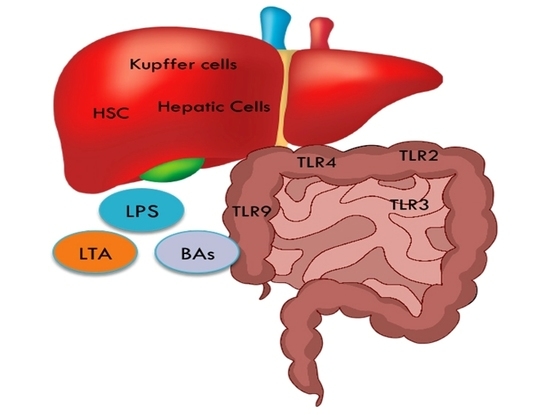
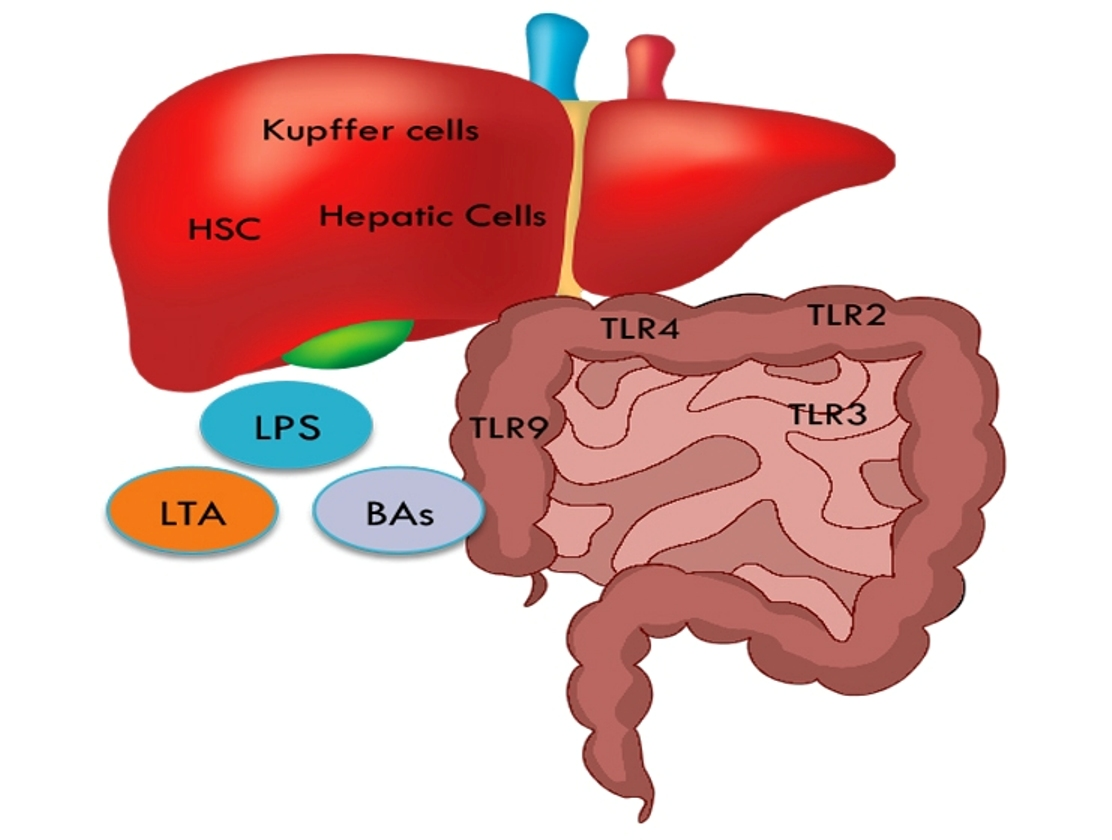
The “gut liver axis” is defined as the bidirectional relationship between the liver and the intestinal microbiome and involves signaling pathways induced by genetic, dietary and environmental elements. The intestinal barrier function can be compromised by disproportionate consumption of tissue damaging foods, like high-fat diet (HFD) components. As a result, the levels of intestinal microbes, termed pathogen-associated molecular patterns (PAMPs), surge and are allowed to be transferred to the liver. PAMPs translocation induces inflammatory responses via the Toll-like receptors (TLRs) and can lead to CLD by promoting inflammation and fibrosis [31]. For each TLR, there is an assumed corresponding gut bacterial product that acts as a ligand (Table 2). Furthermore, a compromised intestinal barrier allows the release of bacterial products in the portal circulation and the liver cells are the first to be exposed to. Continuous damage to the liver can lead to further transportation of bacterial components, which is characteristic of CLD and contributes to cirrhosis and HCC.
Dapito et al., have shown that HCC was dramatically reduced at the last stages in a mouse model by gut sterilization after antibiotic administration [6]. Moreover, in almost 80% of murine, HCCs were prevented by a complete decontamination of the intestine suggesting that the intestinal microbiota is a potential target for HCC prevention [6].
4. How Does A HCC Protective Diet Influence the Gut Microbiome?
The majority of the studies agree that consumption of vegetables, fruits, cereals, nuts, legumes, modest consumption of fish and dairy, and decreased intake of red and/or processed meat and sugar has a chemoprotective role in HCC development. The closest to such a diet is the Mediterranean, which is rich in fiber and unsaturated fats and is known to be a balanced and healthy regime [32]. The Mediterranean diet contributes to cancer prevention by modulating gut microbiota and by reducing metabolic endoxermia [26]. The latter is defined as a twofold surge of the circulating levels of bacterial endotoxins generated in the gut, such as Lipoteichoic acid (LTA) and Lipopolysaccharide (LPS) [33].
Firm adherence to a Mediterranean diet is significantly associated to a vegan diet and microbiome composition and function [34]. Vegetarian and vegan diets were found to enrich fiber-degrading Prevotella and Lachnospira bacteria in the gut, compared to an omnivorous diet [34]. In addition, short-chain fatty acids (SCFA) levels significantly correlate with Mediterranean diet patterns [34]. Clinical and observational studies, in which total food interventions were used, showed that increased walnut or almond consumption, rich in unsaturated fats and fiber, elevates some Gram-positive bacteria such as the Clostridia genera including Oscillospira, Lachnospira, Faecalibacterium, Roseburia and Clostridium, while it reduces other bacteria species such as Dorea, Ruminococcus, and Bifidobacterium [33].
On the contrary, a diet that contains low fiber levels and surged saturated fat levels can contribute to metabolic endotoxemia via alterations in the composition of the gut microbiome, the products of bacterial fermentation, gut barrier permeability and function, and circulating levels of enterohepatic bile acids [33]. Undoubtedly, there is a need to conduct large-scale prospective clinical trials to understand the processes by which a Mediterranean diet can protect against HCC.
5. Bacterial Components and Metabolites
5.1. LPS
LPS is a component of the outer membrane of Gram-negative bacteria, and LTA is a cell membrane constituent of Gram-positive bacteria. Both LTA and LPS can induce liver inflammation and fibrosis through their interaction with TLRs [6]. Studies using mouse models demonstrated that LPS hepatic translocation is crucial in the development of liver cirrhosis and liver cancer. Seki et al. showed that continuous signaling from LPS-induced TLR4 activation promotes fibrosis, while increased LPS levels have been reported in the portal vein of cirrhotic patients [35,36].
A HFD has been shown to dramatically accelerate oncogenes or chemical carcinogen-induced HCC in the mouse [37,38]. Excess dietary fats enhance intestinal permeability facilitating the transportation of LPS and other bacterial fragments to the liver through the enterohepatic system [39]. In turn, LPS activates TLR4 expressed in resident hepatocytes, liver macrophages (Kupffer cells) and hepatic stellate cells and triggers intracellular NF-κB and MAPK cascades that orchestrate pro-inflammatory and proliferative responses6. CD14, a co-factor of TLR4, is upregulated in fatty liver, indicating that chronic adherence to a high-fat diet may sensitize Kupffer cells to even low LPS signals [40]. In addition, TLR4 specific innate immunity activates the TGF-β signal in hepatic stellate cells by inhibiting TGF-β expression by downregulating a decoy receptor for TGF-β, named Bmp and activin membrane-bound inhibitor (BAMBI), hence facilitating promoting liver fibrosis 36 (Figure 1).
Further evidence to support that the LPS-TLR4 interactions contribute to liver cirrhosis comes from a gut microbiota profiling analysis that showed increased levels of Gram-negative bacteria, in particular, Prevotella and Veillonella species, in cirrhotic patients [25]. Interestingly, the same group further demonstrated in a mouse model that constant exposure to LPS promoted hepatocarcinogenesis [6]. LPS-TLR4 interactions do so by inducing proliferative and anti-apoptotic signals as well as by increasing epiregulin and hepatocyte growth factor in local hepatocytes [6].
LPS-TLR4 signaling pathways can induce liver inflammation, and therefore, contribute to liver cirrhosis and hepatocarcinogenesis. Furthermore, CD14, a known compound of TLR4 that can bind to LPS, is highly expressed in fatty liver and the LPS-TLR4 signaling is activated [6]. This also indicates that enduring exposure to even low LPS concentrations may result in fatty liver related to chronic hepatitis. Considering these functions of the TLR4 pathway in liver inflammation, the potential role of TLR4 antagonists, like E5564 and TAK-242, in HCC therapy has been suggested.
5.2. LTA
A HFD induces dysbiosis including enrichment of C. Sordellii from Clostridium cluster XI that produce deoxycholic acid (DCA) [37,38]. LTA is also detected in the livers of these mice following administration of a chemical carcinogen [37]. Together, LTA and DCA stimulate a senescence secretory program in liver stellate cells that includes IL-6, CXCL9 and PGE2, which is critical for hepatocarcinogenesis [37]. Mechanistically, LTA and DCA cooperate to activate TLR2-mediated signaling which, presumably through a TPL2-MEK-ERK cascade [41], leads to the expression of cyclooxygenase 2 (COX2), the enzyme that catalyses a compelling step in PGE2 synthesis. PGE2 in turn suppresses liver resident CD8+ T lymphocytes [37] and most likely other immune cell types, thereby creating an immunosuppressive environment conducing to cancer (Figure 1). A recent study has further implicated DCA in HCC progression through the activation of the mTOR pathway [42].
5.3. Bile Acids
Dysbiosis-associated gut microbes facilitate the breakdown of primary bile acids (PBAs) into secondary bile acids (SBAs) that circulate back to the liver. PBAs and SBAs have different effects on sinusoidal endothelial cells that reside in the liver forming the lining of liver blood capillaries that receive the blood coming from the gut. Unlike PBAs, SBAs suppress the endothelial cell expression of CXCL16 which serves as the ligand of CXCR6 expressed in Natural Killer T (NKT) lymphocytes [43]. As a result, hepatic NKT cell numbers decrease, causing reduced anti-tumor immunity that accelerates HCC [43] (Figure 1).
The relevance of these experimental observations to human HCC remains to be shown. Notably, however, obesity increases predisposition to several types of malignancy, including liver cancer. Tantalizing associations also support the link between gut dysbiosis and HCC. For example, the levels of the PBA cheno-deoxycholic acid in HCC patients associate with CXCL16 synthesis, whereas an inverse correlation was observed with the SBA glycolithocholate (GLCA) [43]. Additionally, COX-2 and PGE2 over-expression are detected in hepatic stellate cells of human non-cirrhotic NASH [37].
6. Epigenetic Control of Treg Cells through SCFAs
Gut microbial products may impact liver physiology also via epigenetic mechanisms that, however, remain largely elusive. A HFD causes alterations in gene methylation patterns in the mouse liver, some of which persist following normalization of diet [44]. Notably, the Odoribacter splanchnicus species that belong to Odoribacter genera, were reported to persist even upon reversal of a HFD, along with the hypomethylation of the Apoa4 gene [44]. Odoribacter produces butyrate, which is known to act as a histone deacetylate inhibitor and is thus likely to impact methylation patterns in the liver. Notably, SCFAs, like acetic and butyric acids as well as gut microbiota products, can inhibit inflammation via the activation of regulatory T cells by epigenetic control [43].
7. Conclusions
HCC is the foremost cause of death in CLD and compensated cirrhosis patients. The development of more effective therapy and preventive methods are paramount. Increasing evidence shows that gut microbiota composition contributes significantly to the progression of liver fibrosis from chronic inflammation. We know that the underlying mechanisms involve the translocation of microbiota components and metabolites into the liver due to an impaired intestinal barrier. More specifically, the gut microbiota components and metabolites, such as LPS, LTA, BAs, fatty acids as well as other unknown microbial metabolites and toxins, affect the development of HCC.
The role of diet-induced changes in gut microbiome composition in liver disease is largely underestimated. Longitudinal clinical studies on the effects of a diet similar to a Mediterranean diet on gut microbiota, metabolic endotoxemia and liver inflammation are necessary to determine efficacious dietary strategies for reducing the risk for HCC development. There is a requirement to elaborate a dietary pattern that can be integrated in an interdisciplinary approach of HCC management.
Author Contributions
Writing—Original Draft Preparation, L.K.; writing—review and editing, L.K., E.V. and A.G.E.; Conceptualization: L.K. and A.G.E.; supervision and critical revision, L.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
Chronic Liver Disease (CLD); Cyclooxygenase 2 (COX2); Deoxycholic acid (DCA); Glycemic load (GL); Glycolithocholate (GLCA); Hepatocellular carcinoma (HCC); High fat diet (HFD); Heterocyclic amines (HCA); Lipopolysaccharide (LPS); Lipoteichoic acid (LTA); N-nitroso compounds (NOC); N-acetyltransferase 2 (NAT2); Natural Killer T (NKT); Pathogen-Associated Molecular Patterns (PAMPs); Primary Bile acids (PBAs); Secondary bile acids (SBAs); Short Chain Fatty Acids (SCFA); Toll-like receptors (TLRs).
References
- Giannitrapani, L.; Zerbo, M.; Amodeo, S.; Pipitone, E.; Galia, M.; Li Cavoli, T.V.; Minissale, M.G.; Licata, A.; Schiavone, C.; Brancatelli, G.; et al. The Changing Epidemiology of Hepatocellular Carcinoma: Experience of a Single Center. Biomed Res. Int. 2020, 2020. [Google Scholar] [CrossRef] [Green Version]
- Asgharpour, A.; Cazanave, S.C.; Pacana, T.; Seneshaw, M.; Vincent, R.; Banini, B.A.; Kumar, D.P.; Daita, K.; Min, H.K.; Mirshahi, F.; et al. A Diet-Induced Animal Model of Non-Alcoholic Fatty Liver Disease and Hepatocellular Cancer. J. Hepatol. 2016, 65, 579–588. [Google Scholar] [CrossRef] [Green Version]
- Koumbi, L. Dietary Factors Can Protect against Liver Cancer Development. World J. Hepatol. 2017, 9, 119–125. [Google Scholar] [CrossRef]
- Gorham, J.; Gleeson, M. Cirrhosis and Dysbiosis: New Insights from next-Generation Sequencing. Hepatology 2016, 63, 336–338. [Google Scholar] [CrossRef]
- Raj, A.S.; Shanahan, E.R.; Tran, C.D.; Bhat, P.; Fletcher, L.M.; Vesey, D.A.; Morrison, M.; Holtmann, G.; Macdonald, G.A. Dysbiosis of the Duodenal Mucosal Microbiota Is Associated with Increased Small Intestinal Permeability in Chronic Liver Disease. Clin. Transl. Gastroenterol. 2019, 10. [Google Scholar] [CrossRef]
- Dapito, D.H.; Mencin, A.; Gwak, G.Y.; Pradere, J.P.; Jang, M.K.; Mederacke, I.; Caviglia, J.M.; Khiabanian, H.; Adeyemi, A.; Bataller, R.; et al. Promotion of Hepatocellular Carcinoma by the Intestinal Microbiota and TLR4. Cancer Cell 2012, 21, 504–516. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Sui, J.; Ma, Y.; Simon, T.G.; Chong, D.; Meyerhardt, J.A.; Willett, W.C.; Giovannucci, E.L.; Chan, A.T.; Zhang, X. A Prospective Study of Dairy Product Intake and the Risk of Hepatocellular Carcinoma in U.S. Men and Women. Int. J. Cancer 2020, 146, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Godos, J.; Tieri, M.; Ghelfi, F.; Titta, L.; Marventano, S.; Lafranconi, A.; Gambera, A.; Alonzo, E.; Sciacca, S.; Buscemi, S.; et al. Dairy Foods and Health: An Umbrella Review of Observational Studies. Int. J. Food Sci. Nutr. 2020, 71, 138–151. [Google Scholar] [CrossRef]
- Talamini, R.; Polesel, J.; Montella, M.; Dal Maso, L.; Crispo, A.; Tommasi, L.G.; Izzo, F.; Crovatto, M.; La Vecchia, C.; Franceschi, S. Food Groups and Risk of Hepatocellular Carcinoma: A Multicenter Case-Control Study in Italy. Int. J. Cancer 2006, 119, 2916–2921. [Google Scholar] [CrossRef] [PubMed]
- Tieri, M.; Ghelfi, F.; Vitale, M.; Vetrani, C.; Marventano, S.; Lafranconi, A.; Godos, J.; Titta, L.; Gambera, A.; Alonzo, E.; et al. Whole Grain Consumption and Human Health: An Umbrella Review of Observational Studies. Int. J. Food Sci. Nutr. 2020, 71, 668–677. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Yang, Y.; Liu, J.; Lu, K.; Tang, Z.; Liu, P.; Liu, L.; Zhu, Y. Systematic Review with Meta-Analysis: Meat Consumption and the Risk of Hepatocellular Carcinoma. Aliment. Pharmacol. Ther. 2014, 39, 913–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwingshackl, L.; Schwedhelm, C.; Galbete, C.; Hoffmann, G. Adherence to Mediterranean Diet and Risk of Cancer: An Updated Systematic Review and Meta-Analysis. Nutrients 2017, 9, 1063. [Google Scholar] [CrossRef] [PubMed]
- Angelino, D.; Godos, J.; Ghelfi, F.; Tieri, M.; Titta, L.; Lafranconi, A.; Marventano, S.; Alonzo, E.; Gambera, A.; Sciacca, S.; et al. Fruit and Vegetable Consumption and Health Outcomes: An Umbrella Review of Observational Studies. Int. J. Food Sci. Nutr. 2019, 70, 652–667. [Google Scholar] [CrossRef]
- Gao, M.; Sun, K.; Guo, M.; Gao, H.; Liu, K.; Yang, C.; Li, S.; Liu, N. Fish Consumption and N-3 Polyunsaturated Fatty Acids, and Risk of Hepatocellular Carcinoma: Systematic Review and Meta-Analysis. Cancer Causes Control 2015, 26, 367–376. [Google Scholar] [CrossRef] [PubMed]
- Shawon, M.A.-A.; Yousuf, M.A.K.; Raheem, E.; Ahmed, S.; Dipti, T.T.; Hoque, M.R.; Taniguchi, H.; Karim, M.R. Epidemiology, Clinical Features, and Impact of Food Habits on the Risk of Hepatocellular Carcinoma: A Case-Control Study in Bangladesh. PLoS ONE 2020, 15, e0232121. [Google Scholar] [CrossRef]
- Marventano, S.; Godos, J.; Tieri, M.; Ghelfi, F.; Titta, L.; Lafranconi, A.; Gambera, A.; Alonzo, E.; Sciacca, S.; Buscemi, S.; et al. Egg Consumption and Human Health: An Umbrella Review of Observational Studies. Int. J. Food Sci. Nutr. 2020, 71, 325–331. [Google Scholar] [CrossRef]
- Fedirko, V.; Lukanova, A.; Bamia, C.; Trichopolou, A.; Trepo, E.; Nöthlings, U.; Schlesinger, S.; Aleksandrova, K.; Boffetta, P.; Tjønneland, A.; et al. Glycemic Index, Glycemic Load, Dietary Carbohydrate, and Dietary Fiber Intake and Risk of Liver and Biliary Tract Cancers in S. Ann. Oncol. 2013, 24, 543–553. [Google Scholar] [CrossRef]
- Ma, Y.; Yang, W.; Li, T.; Liu, Y.; Simon, T.G.; Sui, J.; Wu, K.; Giovannucci, E.L.; Chan, A.T.; Zhang, X. Meat Intake and Risk of Hepatocellular Carcinoma in Two Large US Prospective Cohorts of Women and Men. Int. J. Epidemiol. 2019, 48, 1863–1871. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.S.; Chern, H.D.; Wu, J.C.; Chao, Y.; Huang, Y.H.; Chang, F.Y.; Lee, S.D. Polymorphism of the N-Acetyltransferase 2 Gene, Red Meat Intake, and the Susceptibility of Hepatocellular Carcinoma. Am. J. Gastroenterol. 2003, 98, 1417–1422. [Google Scholar] [CrossRef]
- Fedirko, V.; Trichopolou, A.; Bamia, C.; Duarte-Salles, T.; Trepo, E.; Aleksandrova, K.; Nöthlings, U.; Lukanova, A.; Lagiou, P.; Boffetta, P.; et al. Consumption of Fish and Meats and Risk of Hepatocellular Carcinoma: The European Prospective Investigation into Cancer and Nutrition (EPIC). Ann. Oncol. 2013, 24, 2166–2173. [Google Scholar] [CrossRef] [PubMed]
- Qin, N.; Yang, F.; Li, A.; Prifti, E.; Chen, Y.; Shao, L.; Guo, J.; Le Chatelier, E.; Yao, J.; Wu, L.; et al. Alterations of the Human Gut Microbiome in Liver Cirrhosis. Nature 2014, 513, 59–64. [Google Scholar] [CrossRef]
- Huang, R.-X.; Duan, Y.-Y.; Hu, J.-A. Fish Intake and Risk of Liver Cancer: A Meta-Analysis. PLoS ONE 2015, 10, e0096102. [Google Scholar] [CrossRef]
- Freedman, N.D.; Cross, A.J.; McGlynn, K.A.; Abnet, C.C.; Park, Y.; Hollenbeck, A.R.; Schatzkin, A.; Everhart, J.E.; Sinha, R. Association of Meat and Fat Intake with Liver Disease and Hepatocellular Carcinoma in the NIH-AARP Cohort. J. Natl. Cancer Inst. 2010, 102, 1354–1365. [Google Scholar] [CrossRef] [Green Version]
- George, E.S.; Sood, S.; Broughton, A.; Cogan, G.; Hickey, M.; Chan, W.S.; Sudan, S.; Nicoll, A.J. The Association between Diet and Hepatocellular Carcinoma: A Systematic Review. Nutrients 2021, 13, 172. [Google Scholar] [CrossRef] [PubMed]
- Endres, S.; Ghorbani, R.; Kelley, V.E.; Georgilis, K.; Lonnemann, G.; van der Meer, J.W.M.; Cannon, J.G.; Rogers, T.S.; Klempner, M.S.; Weber, P.C.; et al. The Effect of Dietary Supplementation with n—3 Polyunsaturated Fatty Acids on the Synthesis of Interleukin-1 and Tumor Necrosis Factor by Mononuclear Cells. N. Engl. J. Med. 1989, 320, 265–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Negri, E.; La Vecchia, C.; Franceschi, S.; D’Avanzo, B.; Parazzini, F. Vegetable and Fruit Consumption and Cancer Risk. Int. J. Cancer 1991, 48, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Zamora-Ros, R.; Knaze, V.; Luján-Barroso, L.; Romieu, I.; Scalbert, A.; Slimani, N.; Hjartåker, A.; Engeset, D.; Skeie, G.; Overvad, K.; et al. Differences in Dietary Intakes, Food Sources and Determinants of Total Flavonoids between Mediterranean and Non-Mediterranean Countries Participating in the European Prospective Investigation into Cancer and Nutrition (EPIC) Study. Br. J. Nutr. 2013, 109, 1498–1507. [Google Scholar] [CrossRef] [PubMed]
- Daniell, E.L.; Ryan, E.P.; Brick, M.A.; Thompson, H.J. Dietary Dry Bean Effects on Hepatic Expression of Stress and Toxicity-Related Genes in Rats. Br. J. Nutr. 2020, 108, S37–S45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtani, N.; Kawada, N. Role of the Gut-Liver Axis in Liver Inflammation, Fibrosis, and Cancer: A Special Focus on the Gut Microbiota Relationship. Hepatol. Commun. 2019, 3, 456–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiest, R.; Albillos, A.; Trauner, M.; Bajaj, J.S.; Jalan, R. Targeting the Gut-Liver Axis in Liver Disease. J. Hepatol. 2017, 67, 1084–1103. [Google Scholar] [CrossRef] [Green Version]
- Mentella, M.C.; Scaldaferri, F.; Ricci, C.; Gasbarrini, A.; Miggiano, G.A.D. Cancer and Mediterranean Diet: A Review. Nutrients 2019, 11, 2059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, M.A.; Holscher, H.D. Microbiome-Mediated Effects of the Mediterranean Diet on Inflammation. Adv. Nutr. 2018, 9, 93–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-Level Adherence to a Mediterranean Diet Beneficially Impacts the Gut Microbiota and Associated Metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef]
- Schwabe, R.F.; Luedde, T. Apoptosis and Necroptosis in the Liver: A Matter of Life and Death. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 738–752. [Google Scholar] [CrossRef] [PubMed]
- Seki, E.; De Minicis, S.; Österreicher, C.H.; Kluwe, J.; Osawa, Y.; Brenner, D.A.; Schwabe, R.F. TLR4 Enhances TGF-β Signaling and Hepatic Fibrosis. Nat. Med. 2007, 13, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Loo, T.M.; Kamachi, F.; Watanabe, Y.; Yoshimoto, S.; Kanda, H.; Arai, Y.; Nakajima-Takagi, Y.; Iwama, A.; Koga, T.; Sugimoto, Y.; et al. Gut Microbiota Promotes Obesity-Associated Liver Cancer through Pge2-Mediated Suppression of Antitumor Immunity. Cancer Discov. 2017, 7, 522–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimoto, S.; Loo, T.M.; Atarashi, K.; Kanda, H.; Sato, S.; Oyadomari, S.; Iwakura, Y.; Oshima, K.; Morita, H.; Hattori, M.; et al. Obesity-Induced Gut Microbial Metabolite Promotes Liver Cancer through Senescence Secretome. Nature 2013, 499, 97–101. [Google Scholar] [CrossRef]
- Rohr, M.W.; Narasimhulu, C.A.; Rudeski-Rohr, T.A.; Parthasarathy, S. Negative Effects of a High-Fat Diet on Intestinal Permeability: A Review. Adv. Nutr. 2020, 11, 77–91. [Google Scholar] [CrossRef] [Green Version]
- Imajo, K.; Fujita, K.; Yoneda, M.; Nozaki, Y.; Ogawa, Y.; Shinohara, Y.; Kato, S.; Mawatari, H.; Shibata, W.; Kitani, H.; et al. Hyperresponsivity to Low-Dose Endotoxin during Progression to Nonalcoholic Steatohepatitis Is Regulated by Leptin-Mediated Signaling. Cell Metab. 2012, 16, 44–54. [Google Scholar] [CrossRef] [Green Version]
- Eliopoulos, A.G.; Dumitru, C.D.; Wang, C.C.; Cho, J.; Tsichlis, P.N. Induction of COX-2 by LPS in Macrophages Is Regulated by Tpl2-Dependent CREB Activation Signals. EMBO J. 2002, 21, 4831–4840. [Google Scholar] [CrossRef] [Green Version]
- Yamada, S.; Takashina, Y.; Watanabe, M.; Nagamine, R.; Saito, Y.; Kamada, N.; Saito, H. Bile Acid Metabolism Regulated by the Gut Microbiota Promotes Non-Alcoholic Steatohepatitis-Associated Hepatocellular Carcinoma in Mice. Oncotarget 2018, 9, 9925–9939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, C.; Han, M.; Heinrich, B.; Fu, Q.; Zhang, Q.; Sandhu, M.; Agdashian, D.; Terabe, M.; Berzofsky, J.A.; Fako, V.; et al. Gut Microbiome–Mediated Bile Acid Metabolism Regulates Liver Cancer via NKT Cells. Science 2018, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Worsley, O.; Yang, E.; Purbojati, R.W.; Liang, A.L.; Tan, W.; Moses, D.I.D.; Hartono, S.; Fan, V.; Lim, T.K.H.; et al. Persistent Changes in Liver Methylation and Microbiome Composition Following Reversal of Diet-Induced Non-Alcoholic-Fatty Liver Disease. Cell. Mol. Life Sci. 2019, 76, 4341–4354. [Google Scholar] [CrossRef] [PubMed]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T.; et al. Commensal Microbe-Derived Butyrate Induces the Differentiation of Colonic Regulatory T Cells. Nature 2013, 504, 446–450. [Google Scholar] [CrossRef]
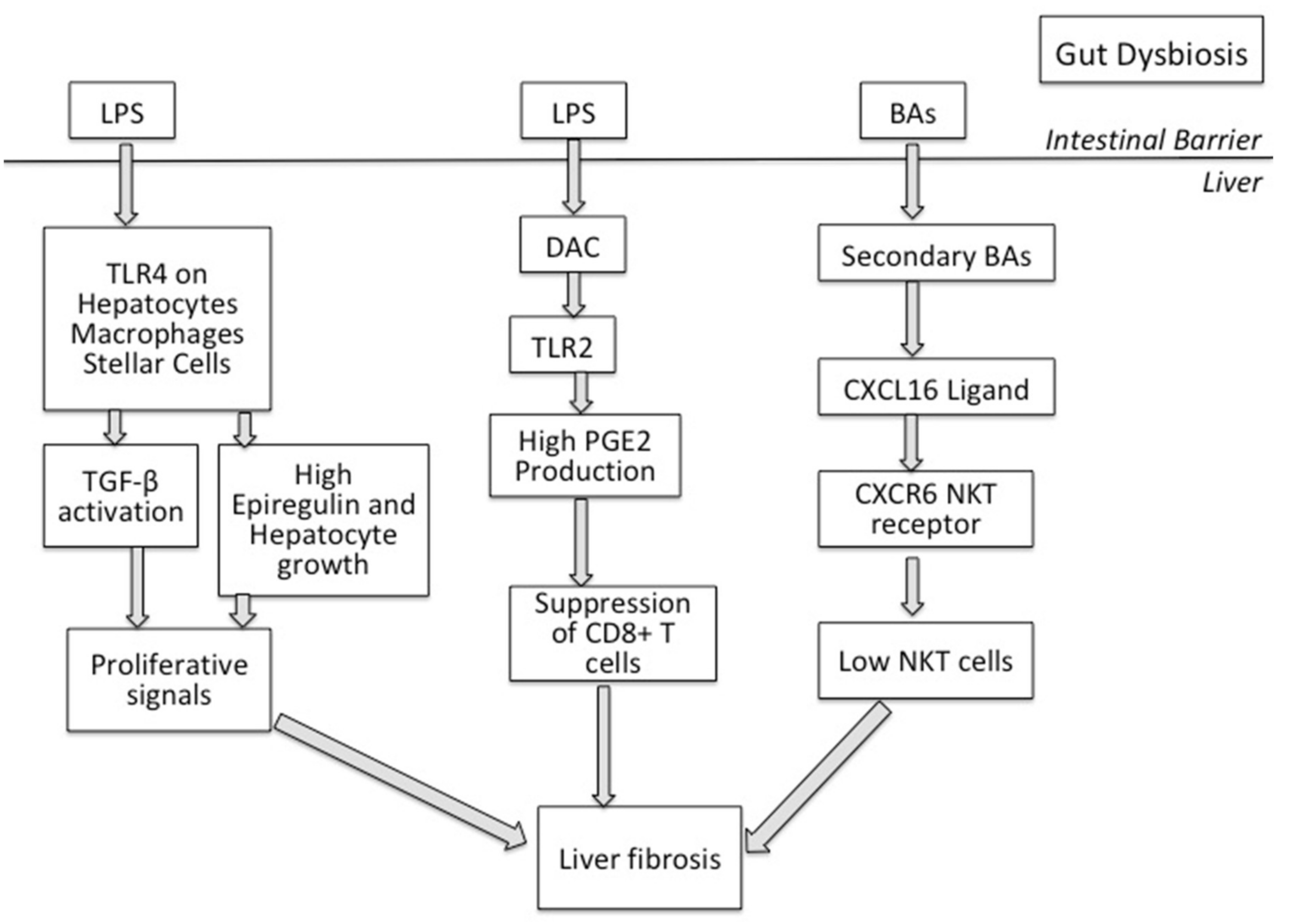
Figure 1.
Gut dysbiosis allows the translocation of gut microbe products, like Lipopolysaccharide (LPS) LPS, Lipoteichoic acid (LTA) and bile acids (BAs). LPS translocates and interacts with the Tolle-like Receptor 4 (TLR4) expressed on hepatocytes, macrophages and stellar cells in the liver, inducing proliferative responses and increasing epiregulin and hepatocyte growth. LPS-TLR4 interaction activates TGF-β signaling in stellate cells and further promotes liver fibrosis. LTA and deoxycholic acid (DCA) interaction leads to TLR2 activation and increased PGE2 production that, in turn, suppress liver CD8+ T cells, inducing liver fibrosis. Secondary BAs in the liver suppress CXCL16 ligand, decreasing hepatic NKT numbers and hence accelerating liver fibrosis.
Figure 1.
Gut dysbiosis allows the translocation of gut microbe products, like Lipopolysaccharide (LPS) LPS, Lipoteichoic acid (LTA) and bile acids (BAs). LPS translocates and interacts with the Tolle-like Receptor 4 (TLR4) expressed on hepatocytes, macrophages and stellar cells in the liver, inducing proliferative responses and increasing epiregulin and hepatocyte growth. LPS-TLR4 interaction activates TGF-β signaling in stellate cells and further promotes liver fibrosis. LTA and deoxycholic acid (DCA) interaction leads to TLR2 activation and increased PGE2 production that, in turn, suppress liver CD8+ T cells, inducing liver fibrosis. Secondary BAs in the liver suppress CXCL16 ligand, decreasing hepatic NKT numbers and hence accelerating liver fibrosis.


{kind=link}
{kind=link}
Table 1.
The effect of dietary components in hepatocellular carcinoma risk.
| Dietary Food Intake Associated with HCC Risk | |||
|---|---|---|---|
| Dietary Components | Association Type with HCC Risk | Country of Study | References |
| Dietary Fiber | Positive | Europe Rat Model | Fedirko et al. 2013 [17] Daniel et al. 2020 [18] |
| Fish | Inverse | Meta-analysis Meta-analysis USA Western Europe | Gao et al. 2015 [14] Huang et al. 2015 [19] Freedman et al. 2010 [20] Fedirko et al. 2013 [17] |
| Fruits | Inverse | Meta-analysis Europe | Schwingshackl et al. 2017 [12] Zamora-Ros et al. 2013 [21] |
| Glycemic load | Positive | Europe | Fedriko et al. 2013 [17] |
| Dairy products | Inverse | Meta-analysis USA | Godos et al. 2020 [8] Yang et al. 2020 [7] |
| Red meat | Positive | USA Europe | Ma et al. 2019 [22] Fedriko et al. 2013 [17] |
| White Meat | Inverse | Meta-analyses Western Europe | Luo et al. 2014 [11] Fedirko et al. 2013 [17] |
| Tea | Inverse | Italy | Talamini et al. 2006 [23] |
| Vegetables | Inverse | Bangladesh | Shawon et al. 2020 [15] Zamora-Ros et al. 2013 [21] |
Table 2.
Toll–like receptors (TLR) and their corresponding ligands in the liver.
| Toll Like Receptors | Cellular Targets in the Liver |
|---|---|
| TLR2, TLR4 and TLR9 | Hepatic stellate cells (HSC) |
| TLR2, TLR3, TLR4 and TLR9 | Kupffer cells |
| TLR1-9 | Hepatic cells |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Koumbi, L.; Eliopoulos, A.G.; Vassilopoulou, E. How Diet-Induced Changes in the “Gut-Liver” Axis Affect Chronic Liver Disease Outcome? Livers 2021, 1, 40-48. https://0-doi-org.brum.beds.ac.uk/10.3390/livers1010004
AMA Style
Koumbi L, Eliopoulos AG, Vassilopoulou E. How Diet-Induced Changes in the “Gut-Liver” Axis Affect Chronic Liver Disease Outcome? Livers. 2021; 1(1):40-48. https://0-doi-org.brum.beds.ac.uk/10.3390/livers1010004
Chicago/Turabian StyleKoumbi, Lemonica, Aristides G. Eliopoulos, and Emilia Vassilopoulou. 2021. "How Diet-Induced Changes in the “Gut-Liver” Axis Affect Chronic Liver Disease Outcome?" Livers 1, no. 1: 40-48. https://0-doi-org.brum.beds.ac.uk/10.3390/livers1010004