Accumulation, Biotransformation, Histopathology and Paralysis in the Pacific Calico Scallop Argopecten ventricosus by the Paralyzing Toxins of the Dinoflagellate Gymnodinium catenatum


Abstract
:1. Introduction
2. Results and Discussion
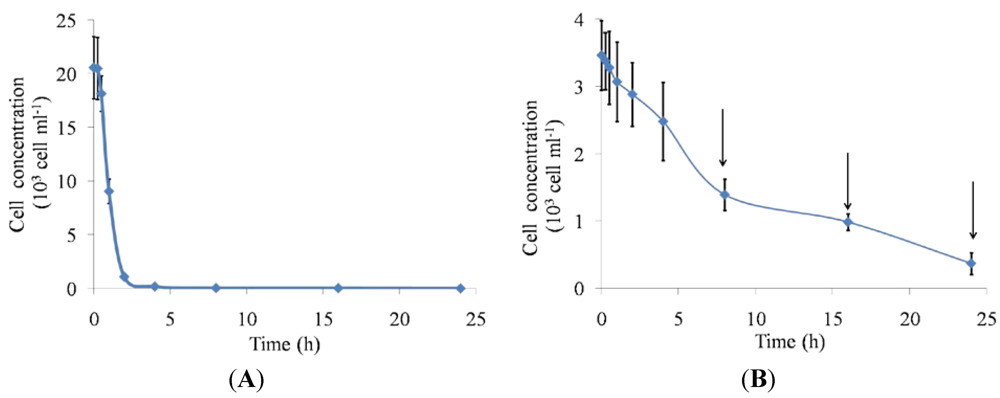
2.1. Feeding Responses

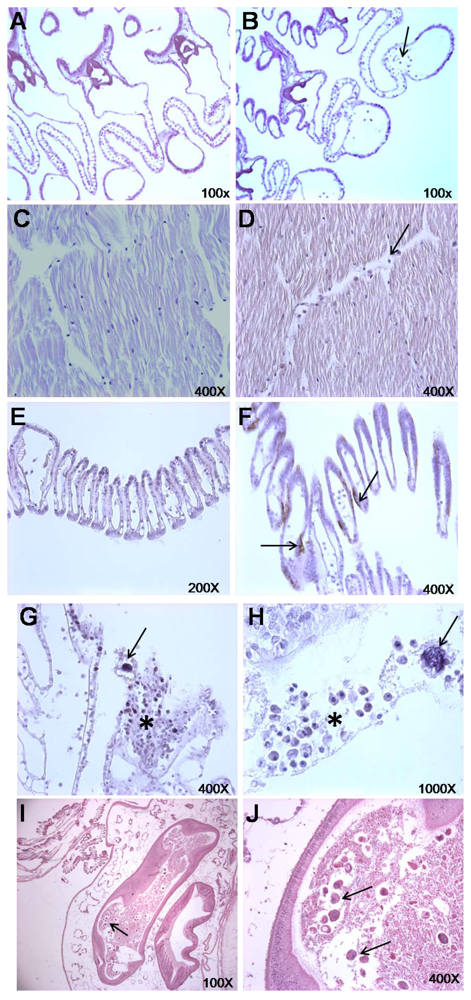
2.2. Histopathological Findings


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Negative | Positive | χ2 | P | |||
|---|---|---|---|---|---|---|---|
| N | % | n | % | ||||
| Gills | |||||||
| Epithelial melanization | 22.5 | <0.0001 | |||||
| Control | 30 | 83.4 | 6 | 16.6 | |||
| Treated | 10 | 27.8 | 26 | 72.2 | |||
| Mucus production | 14.5 | 0.0001 | |||||
| Control | 34 | 94.4 | 2 | 5.6 | |||
| Treated | 20 | 55.6 | 16 | 44.4 | |||
| Hemocyte accumulation | 10.0 | 0.0016 | |||||
| Control | 35 | 97.2 | 1 | 2.8 | |||
| Treated | 25 | 69.4 | 11 | 30.6 | |||
| Mantle | |||||||
| Epithelial melanization | 29.6 | <0.0001 | |||||
| Control | 28 | 77.8 | 8 | 22.2 | |||
| Treated | 5 | 13.9 | 31 | 86.1 | |||
| Mucus production | 22.8 | <0.0001 | |||||
| Control | 31 | 86.1 | 5 | 13.9 | |||
| Treated | 11 | 30.6 | 25 | 69.4 | |||
| Hemocyte accumulation | 22.4 | <0.0001 | |||||
| Control | 35 | 97.2 | 1 | 2.8 | |||
| Treated | 17 | 47.2 | 19 | 52.8 | |||
| Muscle | |||||||
| Hemocyte accumulation | |||||||
| Control | 33 | 91.7 | 3 | 8.3 | 22.1 | <0.0001 | |
| Treated | 14 | 38.9 | 22 | 61.1 | |||
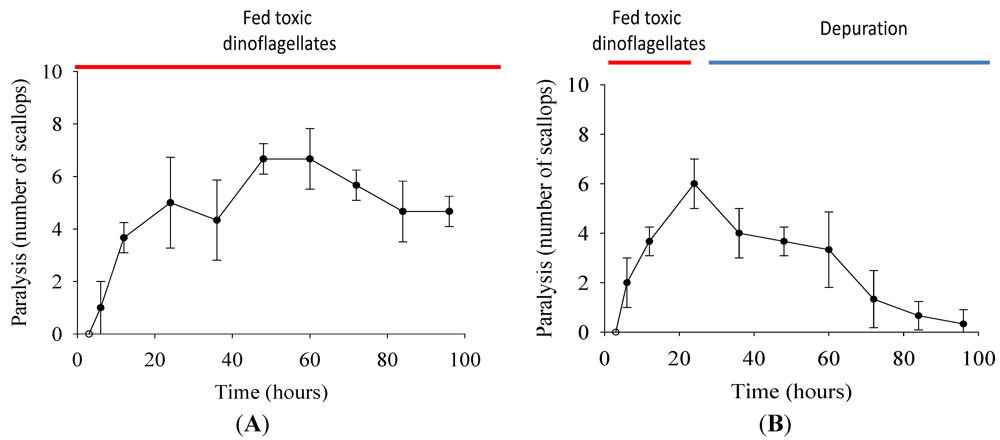
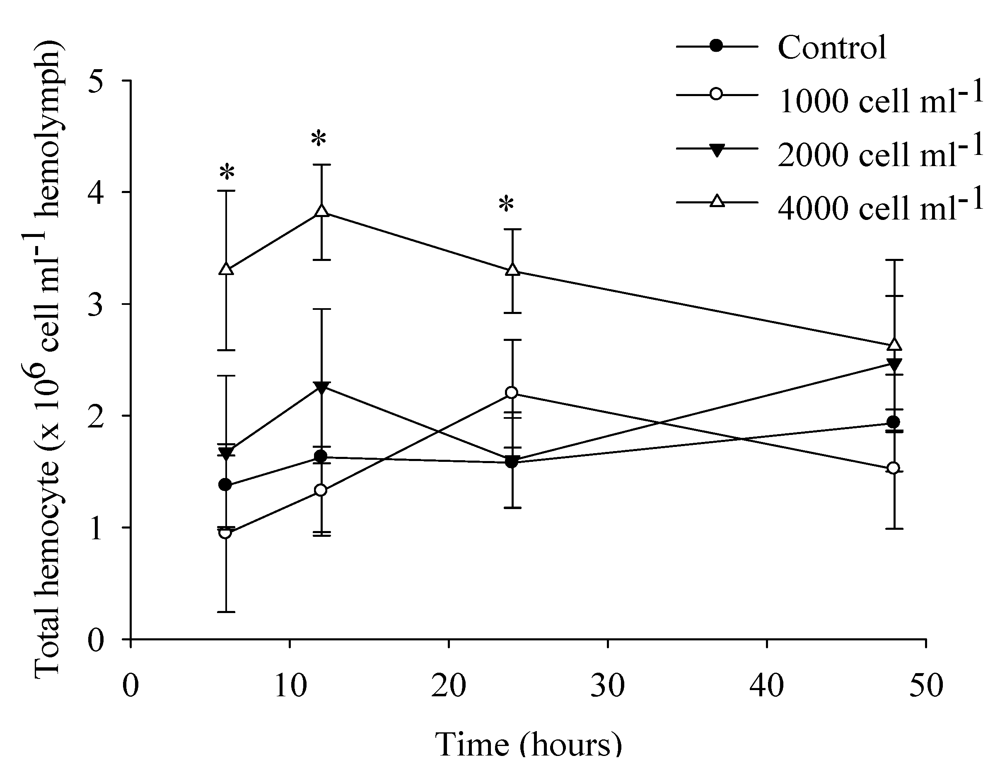
2.3. Paralysis of Scallops


2.4. Analysis of Toxins
| Extract | C1 | C2 |
|---|---|---|
| Gymnodinium catenatum | 45 ± 6 | 15 ± 6 |
| Tissue controls | ND | ND |
| Digestive gland-stomach | 38 ± 3 | 189 ± 13 |
| Mantle | 41 ± 8 | 202 ± 37 |
| Gills | 10 ± 1 | 51 ± 6 |
| Muscle, kidney, rectum | 11 ± 2 | 56 ± 8 |
3. Experimental Section
3.1. Algal Culture and Source of Scallops
3.2. Feeding Experiments
3.2.1. Experiment 1
3.2.2. Experiment 2
3.2.3. Experiment 3
3.3. Depletion of Algal Cells
3.4. Histopathological Examination
Contingency Tables
3.5. Paralysis Studies
3.5.1. Determination of Paralysis
3.5.2. Hemocytes Count at Different Toxic Cells Concentrations
3.6. Analysis of Toxins
3.6.1. Algal and Scallop Toxins
3.6.2. HPLC Analysis
4. Conclusions
Acknowledgments
References
- Graham, H.W. Gymnodinium catenatum, a new dinoflagellate from the Gulf of California. Trans. Am. Microsc. Soc. 1943, 62, 259–261. [Google Scholar] [CrossRef]
- Mee, L.D.; Espinoza, M.; Díaz, G. Paralytic shellfish poisoning with a Gymnodinium catenatum red tide of the Pacific Coast of Mexico. Mar. Environ. Res. 1986, 19, 77–92. [Google Scholar] [CrossRef]
- Cortés, A.R.; Manrique, F.A.; Luna-Soria, R. Occurrence of red tides in the eastern coast of the Gulf of California. Rev. Lat. Microbiol. 1995, 37, 337–342. [Google Scholar]
- Ochoa, J.L.; Sánchez-Paz, A.; Cruz-Villacorta, A.; Núñez-Vázquez, E.; Sierra-Beltrán, A. Toxic events in the northwest Pacific coastline of Mexico during 1992–1995: Origin and impact. Hydrobiologia 1997, 352, 195–200. [Google Scholar]
- Ochoa, J.L.; Sierra-Beltrán, A.; Alonso-Colmenares, G.; Barradas-Sánchez, H.; Cruz-Villacorta, A.; Nuñez-Vázquez, E.; Sánchez-Paz, A. Biotoxins in the Pacific coast of Mexico. In Mycotoxins and Phycotoxins: Developments in Chemistry, Toxicology, and Food Safety, Proceedings of IX IUPAC (International Union Purity Analytical Chemistry) International Symposium on Mycotoxins and Phycotoxins, Rome, Italy, 27–31 May 1996; Miraglia, M., van Egmond, H.P., Brera, C., Gilbert, J., Eds.; Alaken: Fort Collins, CO, USA, 1998; pp. 441–448. [Google Scholar]
- Alonso-Rodríguez, R.; Ochoa, J.L. Hydrology of winter-spring “red tides” in Bahía de Mazatlán, Sinaloa, México. Harmful Algae 2004, 3, 163–171. [Google Scholar] [CrossRef]
- Gárate-Lizárraga, I.; Bustillos-Guzmán, J.J.; Morquecho, L.; Band-Schmidt, C.J.; Alonso-Rodríguez, R.; Erler, K.; Luckas, B.; Reyes-Salinas, A.; Góngora-González, D.T. Comparative paralytic shellfish toxin profiles in the strains of Gymnodinium catenatum Graham from the Gulf of California, Mexico. Mar. Pollut. Bull. 2005, 50, 208–236. [Google Scholar] [CrossRef]
- Band-Schmidt, C.J.; Bustillos-Guzmán, J.; Gárate-Lizárraga, I.; Lechuga-Devèze, C.H.; Reinhardt, K.; Luckas, B. Paralytic shellfish toxin profile in strains of the dinoflagellate Gymnodinium catenatum Graham and the scallop Argopecten ventricosus G. B. Sowerby II from Bahía Conception, Gulf of California, Mexico. Harmful Algae 2005, 4, 21–31. [Google Scholar] [CrossRef]
- Band-Schmidt, C.; Bustillos-Guzmán, J.; Morquecho, L.; Gárate-Lizárraga, I.; Alonso-Rodríguez, R.; Reyes-Salinas, A.; Erler, K.; Luckas, B. Variations of PSP toxin profiles during different growth phases in Gymnodinium catenatum (Dinophyceae) strains isolated from three locations in the Gulf of California, Mexico. J. Phycol. 2006, 42, 757–768. [Google Scholar] [CrossRef]
- Cembella, A.D.; Lewis, N.I.; Shumway, S.E. An interspecific comparison of paralytic shellfish poisons in marine bivalves: Anatomical and spatio-temporal variation in toxin composition. J. Shellfish Res. 1993, 12, 389–403. [Google Scholar]
- Bricelj, M.V.; Shumway, E.S. Paralytic shellfish toxins in bivalve mollusks: Occurrence, transfer kinetics, and biotransformation. Rev. Fish. Sci. 1998, 6, 315–383. [Google Scholar] [CrossRef]
- Weise, M.; D’Angostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar]
- Mons, M.N.; van Egmond, H.P.; Speijers, G.J.A. Paralytic Shellfish Poisoning: A Review; RIVM Report 388802; National Institute of Public Health and the Environment: Bilthoven, The Netherlands, 1998. [Google Scholar]
- Kao, C.Y.; Suzuki, C.Y.; Kleinahus, T.; Siegman, M.J. Vasomotor and respiratory depressant actions of tetrodotoxin and saxitoxin. Arch. Int. Pharmacodyn. 1967, 165, 438–450. [Google Scholar]
- Evans, M.H. Cause of death in experimental paralytic shellfish poisoning. Br. J. Exp. Pathol. 1965, 46, 245–253. [Google Scholar]
- Lagos, N. Paralytic shellfish poisoning phycotoxins: Occurrence in South America. Comments Toxicol. 2003, 9, 175–193. [Google Scholar] [CrossRef]
- Juárez-Ruiz, N.; Ramírez-Camarena, C.; Barón-Campis, S.A. (Eds.) INP-SAGARPA. Registro de mareas rojas en el Pacífico Sur de 1985 a 2004; Dirección General de Investigaciones Pesqueras Pacífico Sur, Instituto Nacional de la Pesca, Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación: Ensenada, Mexico, 2004. Available online: http://fans.cicese.mx/publicaciones/noticias/Cuadro%20Mareas%20Rojas%20Pacifico%20Sur-SEMARNAT.xls (accessed on 26 April 2012).
- Shumway, S.E. A review of the effects of algal blooms on shellfish and aquaculture. J. World Aquac. Soc. 1990, 21, 65–104. [Google Scholar]
- Lagos, N. Microalgal blooms: A global issue with negative impact in Chile. Biol. Res. 1998, 31, 375–386. [Google Scholar]
- Moroño, A.; Franco, F.; Miranda, M.; Reyero, M.I.; Blanco, J. The effect of mussel size, temperature, volume, food quality, and volume-specific toxin concentration on the uptake rate of PSP toxins by mussels (Mytilus galloprovincial). J. Exp. Mar. Biol. Ecol. 2001, 257, 117–132. [Google Scholar] [CrossRef]
- Gainey, L.F.; Shumway, S.E. A compendium of the responses of bivalve mollusks to toxic dinoflagellates. J. Shellfish Res. 1988, 7, 623–628. [Google Scholar]
- Bricelj, V.M.; Cembella, A.D.; Lavy, D.; Shumway, S.E.; Cucci, T.L. Comparative Physiological and Behavioral Responses to PSP Toxins in Two Bivalve Mollusks, the Soft Shell Clam, Mya arenaria, and Surf Clam, Spisula solidissima. In Harmful and Toxic Algal Blooms; Yasumoto, T., Oshima, Y., Fukuyo, Y., Eds.; UNESCO: Paris, France, 1996; pp. 405–408. [Google Scholar]
- Matsuyama, Y.; Uchida, T.; Honjo, T. Effects of harmful dinoflagellates, Gymnodinium mikimotoi and Heterocapsa circularisquama, red-tide on filtering rate of bivalve molluscs. Fish. Sci. 1999, 65, 248–253. [Google Scholar]
- Blanco-Pérez, J. Episodios Nocivos Por Fitoplancton. In Los Moluscos Pectínidos de Iberoamérica: Ciencia y Acuicultura; Maeda-Martínez, A.N., Ed.; Limusa: Mexico City, Mexico, 2001; pp. 285–324. [Google Scholar]
- Estrada, N.A.; Lagos, N.; García, C.; Maeda-Martínez, A.N.; Ascencio, F. Effects of the toxic dinoflagellate Gymnodinium catenatum on uptake and fate of paralytic shellfish poisons in the pacific giant lions-paw scallop Nodipecten subnodosus. Mar. Biol. 2007, 151, 1205–1214. [Google Scholar] [CrossRef]
- Estrada, N.; Romero, M.J.; Campa-Córdova, A.; Luna, A.; Ascencio, A. Effects of the toxic dinoflagellate, Gymnodinium catenatum on hydrolytic and antioxidant enzymes, in tissues of the giant lions-paw scallop Nodipecten subnodosus. Comp. Biochem. Physiol. Part C 2007, 146, 502–510. [Google Scholar]
- Tran, D.; Haberkorn, H.; Soudant, P.; Ciret, P.; Massabuau, J.C. Behavioral responses of Crassostrea gigas exposed to the harmful algae Alexandrium minutum. Aquaculture 2010, 298, 338–345. [Google Scholar] [CrossRef]
- Haberkorn, H.; Tran, D.; Massabuau, J.C.; Ciret, P.; Savar, V.; Soudant, P. Relationship between valve activity, microalgae concentration in the water and toxin accumulation in the digestive gland of the Pacific oyster Crassostrea gigas exposed to Alexandrium minutum. Mar. Pollut. Bull. 2011, 62, 1191–1197. [Google Scholar] [CrossRef] [Green Version]
- Hégaret, H.; Wikfors, G.H.; Soudant, P.; Lambert, C.; Shumway, S.E.; Bérard, J.B.; Lassus, P. Toxic dinoflagellates (Alexandrium fundyense and A. catenella) have minimal apparent effects on oyster hemocytes. Mar. Biol. 2007, 152, 441–447. [Google Scholar] [CrossRef]
- Estrada, N.; Rodríguez-Jaramillo, C.; Contreras, G.; Ascencio, F. Effects of induced paralysis on hemocytes and tissues of the giant lions-paw scallop by paralyzing shellfish poison. Mar. Biol. 2010, 157, 1401–1415. [Google Scholar]
- Hégaret, H.; Wikfors, G.H.; Shumway, S.E. Diverse feeding responses of five species of bivalve mollusc when exposed to three species of harmful algae. J. Shellfish Res. 2007, 26, 549–559. [Google Scholar]
- Lesser, M.P.; Shumway, S.E. Effects of toxic dinoflagellates on clearance rates and survival in juvenile bivalve mollusks. J. Shellfish Res. 1993, 12, 377–381. [Google Scholar]
- Lassus, P.; Ledoux, M.; Bohec, M.; Murail, I.; Fremy, J.M. Role of kidneys in bioaccumulation of paralytic toxins by scallop (Pecten maximus) tissues. J. Nat. Toxins 1996, 1, 107–115. [Google Scholar]
- Shumway, S.E.; Cucci, T.L. The effect of the toxic dinoflagellate Protogonyaulax tamarensis on the feeding behavior of bivalve molluscs. Aquat. Toxicol. 1987, 10, 9–27. [Google Scholar] [CrossRef]
- Oshima, Y.; Sugino, K.; Itakura, H.; Hirota, M.; Yasumoto, T. Comparative Studies on Paralytic Shellfish Toxin Profile of Dinoflagellates in Bivalves. In Toxic Marine Phytoplankton; Graneli, E., Sundstrom, B., Edler, L., Anderson, D., Eds.; Elsevier: New York, NY, USA, 1990; pp. 391–396. [Google Scholar]
- Suzuki, Y.; Yamasaki, M.; Ota, H. Comparison of paralytic shellfish toxin profiles between the scallop Patinopecten yessoensis and the mussel Mytilus galloprovincialis. Fish. Sci. 1998, 64, 850–851. [Google Scholar]
- Hall, S.; Strichartz, G.; Moczydlowski, E.; Ravindran, A.; Reichardt, P.B. The Saxitoxins: Sources, Chemistry and Pharmacology. In Marine Toxins; American Chemical Society Symposium Series 418; Hall, S., Strichartz, G.R., Eds.; American Chemical Society: Washington, DC, USA, 1990; pp. 29–65. [Google Scholar]
- Lehane, L. Paralytic Shellfish Poisoning: A Review; National Office of Animal and Plant Health, Agriculture, Fisheries and Forestry: Canberra, Australia, 2000. [Google Scholar]
- Gárate, L.I.; Bustillos, J.J.; Alonso, R.R.; Luckas, D. Comparative paralytic shellfish toxins profile in two marine bivalves during outbreaks of Gymnodinium catenatum (Dinophyceae) in the Gulf of California. Mar. Pollut. Bull. 2004, 48, 378–402. [Google Scholar] [CrossRef]
- Fast, D.M.; Cembella, D.A.; Ross, W.N. In vitro transformation of paralytic shellfish toxins in the clams Mya arenaria and Protothaca staminea. Harmful Algae 2005, 191, 1–12. [Google Scholar]
- Jaime, E.; Gerdts, G.; Luckas, B. In vitro transformation of PSP toxins by different shellfish tissues. Harmful Algae 2007, 6, 308–316. [Google Scholar] [CrossRef]
- Shumway, S.E.; Cucci, T.L.; Lesser, M.P.; Bourne, N.; Bunting, B. Particle clearance and selection in three species of juvenile scallops. Aquac. Int. 1997, 5, 89–99. [Google Scholar]
- Brillant, M.G.S.; MacDonald, B.A. Postingestive selection in the sea scallop (Placopecten magellanicus) on the basis of chemical properties of particles. Mar. Biol. 2002, 141, 457–465. [Google Scholar] [CrossRef]
- Newell, R.I.E.; Jordan, S.J. Preferential ingestion of organic material by the American oyster Crassostrea virginica. Mar. Ecol. Prog. Ser. 1983, 13, 47–53. [Google Scholar] [CrossRef]
- Negri, A.; Stirling, D.; Quilliam, M.; Blackburn, S.; Bolch, C.; Burton, I.; Eaglesham, G.; Thomas, K.; Walter, J.; Willis, R. Three novel hydroxybenzoate saxitoxin analogues isolated from the dinoflagellate Gymnodinium catenatum. Chem. Res. Toxicol. 2003, 16, 1029–1033. [Google Scholar] [CrossRef]
- Llewellyn, L.; Negri, A.; Quilliam, M. High affinity for the rat brain sodium channel of newly discovered hydroxybenzoate saxitoxin analogues from the dinoflagellate Gymnodinium catenatum. Toxicon 2004, 43, 101–104. [Google Scholar] [CrossRef]
- Negri, A.P.; Bolch, C.J.S.; Geier, S.; Green, D.H.; Park, T.-G.; Blackburn, S.I. Widespread presence of hydrophobic paralytic shellfish toxins in Gymnodinium catenatum. Harmful Algae 2007, 6, 774–780. [Google Scholar] [CrossRef]
- Vale, P. Complex profiles of hydrophobic paralytic shellfish poisoning compounds in Gymnodinium catenatum identified by liquid chromatography with fluorescence detection and mass spectrometry. J. Chromatogr. A 2008, 1195, 85–93. [Google Scholar] [CrossRef]
- Bustillos-Guzmán, J.; Vale, P.; Band-Schmidt, C. Presence of benzoate type toxins in Gymnodinium catenatum Graham isolated from the Mexican Pacific. Toxicon 2011, 57, 922–926. [Google Scholar] [CrossRef]
- Seki, T.; Satake, M.; Mackenzie, L.; Kaspar, H.F.; Yasumoto, T. Gymnodimine, a new marine toxin of unprecedented structure isolated from New Zealand oysters and the dinoflagellate, Gymnodinium sp. Tetrahedron Lett. 1995, 36, 7093–7096. [Google Scholar]
- Miles, C.O.; Wilkins, A.L.; Stirling, D.J.; MacKenzie, A.L. New Analogue of Gymnodimine from a Gymnodinium species. J. Agric. Food Chem. 2000, 48, 1373–1376. [Google Scholar]
- Stirling, D.J. Survey of historical New Zealand shellfish samples for accumulation of gymnodimine. N. Z. J. Mar. Freshw. Res. 2001, 35, 851–857. [Google Scholar]
- Biré, R.; Krys, S.; Frémy, J.M.; Dragacci, S.; Stirling, D.J.; Kharrat, R. First evidence on occurrence of gymnodimine in clams from Tunisia. J. Nat. Toxins 2002, 11, 269–275. [Google Scholar]
- Emura, A.; Matsuyama, Y.; Oda, T. Evidence for the production of a novel proteinaceous hemolytic exotoxin by dinoflagellate Alexandrium taylori. Harmful Algae 2004, 3, 29–37. [Google Scholar] [CrossRef]
- Arzul, G.; Seguel, M.; Guzman, L.; Erard-Le Denn, E. Comparison of allelopathic properties in three toxic Alexandrium species. J. Exp. Mar. Biol. Ecol. 1999, 232, 285–295. [Google Scholar] [CrossRef]
- Tillmann, U.; Alpermann, T.; John, U.; Cembella, A. Allelochemical interactions and short-term effects of the dinoflagellate Alexandrium on selected photoautotrophic and heterotrophic protists. Harmful Algae 2007, 7, 52–64. [Google Scholar]
- Lush, G.J.; Hallegraeff, G.M.; Munday, B.L. High Toxicity of the Red Tide Dinoflagellate Alexandrium minutum to the Brine Shrimp Artemia salina. In Harmful and Toxic Algal Blooms; Yasumoto, T., Oshima, Y., Fukuyo, Y., Eds.; UNESCO: Paris, France, 1996; pp. 389–392. [Google Scholar]
- Bagoien, E.; Miranda, A.; Reguera, B.; Franco, J.M. Effects of two paralytic shellfish toxin producing dinofagellates on the pelagic harpacticoid copepod Euterpina acutifrons. Mar. Biol. 1996, 126, 361–369. [Google Scholar] [CrossRef]
- Brillant, M.G.S.; MacDonald, B.A. Postingestive selection in the sea scallop, Placopecten magellanicus (Gmelin): the role of particle size and density. J. Exp. Mar. Biol. Ecol. 2000, 253, 211–227. [Google Scholar] [CrossRef]
- Beecham, J. Literature Review on Particle Assimilation by Molluscs and Crustaceans; Cefas Contract C2706; The Centre for Environment, Fisheries and Aquaculture Science: Lowestoft, UK, 2008; p. 19. [Google Scholar]
- Cheng, T. Bivalves. In Invertebrate Blood Cells; Ratcliffe, N.A., Rowley, A.F., Eds.; Academic Press: New York, NY, USA, 1981; pp. 233–300. [Google Scholar]
- Fisher, W.S. Structure and Functions of Oyster Hemocytes. In ImmunityinInvertebrates; Brehélin, M., Ed.; Springer Verlag: Berlin, Germany, 1986; pp. 25–35. [Google Scholar]
- Wikfors, G.H.; Smolowitz, R.M. Detrimental Effects of a Prorocentrum Isolate upon Hard Clams and Bay Scallops in Laboratory Feeding Studies. In Toxic Phytoplankton Blooms in the Sea, 1st; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: New York, NY, USA, 1993; pp. 447–452. [Google Scholar]
- Beninger, P.G.; Le Pennec, M.; Donval, A. Mode of particle ingestion in five species of suspension-feeding bivalve molluscs. Mar. Biol. 1991, 108, 255–261. [Google Scholar]
- Beninger, P.G.; Le Pennec, M. A histochemical study of the bucco-oesophageal glands of the blue mussel. Mytilus edulis L.: The importance of mucus in ingestion. J. Mar. Biol. 1993, 73, 237–240. [Google Scholar] [CrossRef]
- Pales, E.E.; Perrigault, M.; Ward, J.E.; Shumway, S.E.; Allam, B. Lectins associated with the feeding organs of the oyster Crassostrea virginica can mediate particle selection. Biol. Bull. 2009, 217, 130–141. [Google Scholar]
- Vargas-Albores, F.; Barracco, M.A. Mecanismos de Defensa de los Moluscos Bivalvos, con Énfasis en Pectínidos. In Los Moluscos Pectínidos de Iberoamérica: Ciencia y Acuicultura; Maeda-Martínez, A.N., Ed.; Limusa: Mexico City, Mexico, 2001; pp. 127–146. [Google Scholar]
- Hégaret, H.; Shumway, S.; Wikfors, G.; Pate, S.; Burkholder, J. Potential transport of harmful algae via relocation of bivalve molluscs. Mar. Ecol. Prog. Ser. 2008, 361, 169–179. [Google Scholar]
- Galimany, E.; Sunila, I.; Hégaret, H.; Ramón, M.; Wikfors, G.H. Pathology and immune response of the blue mussel (Mytilus edulis L.) after an exposure to the harmful dinoflagellate Prorocentrum minimum. Harmful Algae 2008, 7, 630–638. [Google Scholar] [CrossRef]
- Bricelj, V.M.; Lee, J.H.; Cembella, A.D. Influence of dinoflagellate cell toxicity on uptake and loss of paralytic shellfish toxins in the northern quahog Mercenaria mercenaria. Mar. Ecol. Prog. Ser. 1991, 74, 33–46. [Google Scholar] [CrossRef]
- Li, S.C.; Wang, W.X.; Hsieh, D.P.H. Effects of toxic dinoflagellate Alexandrium tamarense on the energy budgets and growth of two marine bivalves. Mar. Environ. Res. 2002, 53, 145–160. [Google Scholar] [CrossRef]
- Hwang, D.F.; Chueh, C.H.; Jeng, S.S. Susceptibility of fish, crustacean and mollusk to tetrodotoxin and paralytic shellfish poison. Nippon Suis. Gak. 1990, 56, 337–343. [Google Scholar]
- Lu, Y.H.; Hwang, D.F. Effects of toxic dinoflagellates and toxin biotransformation in bivalves. J. Nat. Toxins 2002, 11, 315–322. [Google Scholar]
- Bricelj, V.M.; Connell, L.; Konoki, K.; MacQuarrie, S.P.; Scheuer, T.; Catterall, W.A.; Trainer, V.L. Sodium channel mutation leading to saxitoxin resistance in clams increases risk of PSP. Nature 2005, 434, 763–767. [Google Scholar]
- Haberkorn, H.; Lambert, C.; Le Goïc, N.; Moal, J.; Suquet, M.; Guéguen, M.; Sunila, I.; Soudant, P. Effects of Alexandrium minutum exposure on nutrition-related processes and reproductive output in oysters Crassostrea gigas. Harmful Algae 2010, 9, 427–439. [Google Scholar] [CrossRef] [Green Version]
- Bricelj, V.M.; Lee, J.H.; Cembella, A.D.; Anderson, D.M. Uptake of Alexandrium fundyense by Mytilus edulis and Mercenaria mercenaria under Controlled Conditions. In Toxic Marine Phytoplankton; Graneli, E., Sundstrom, B., Edler, L., Anderson, D.M., Eds.; Elsevier Science: New York, NY, USA, 1990; pp. 269–275. [Google Scholar]
- Connell, L.B.; MacQuarrie, S.P.; Twarog, B.M.; Iszard, M.; Bricelj, M. Population differences in nerve resistance to paralytic shellfish toxins in softshell clam, Mya arenaria, associated with sodium channel mutations. Mar. Biol. 2010, 150, 1227–1236. [Google Scholar]
- Hummert, C.; Riutscher, M.; Reinhardt, K.; Luckas, B. Analysis of characteristic PSP profiles produced by Pyrodinium bahamense and several strains of Alexandrium using HPLC based on ion-pair chromatographic separation, post column oxidation and fluorescence detection. Chromatographia 1997, 45, 312–316. [Google Scholar] [CrossRef]
- Yu, R.C.; Hummert, C.; Luckas, B.; Qian, P.Y.; Zhou, M.J. A modified HPLC method for analysis of PSP toxins in algae and shellfish from China. Chromatographia 1998, 48, 671–676. [Google Scholar]
- Raymond, W.M.K.; Weng-Xiong, W.; Paul, K.S.L.; Peter, K.N.Y. The uptake, distribution and elimination of paralytic shellfish toxins in mussels and fish exposed to toxic dinoflagellates. Aquat. Toxicol. 2006, 80, 82–91. [Google Scholar]
- Oshima, Y. Chemical and Enzymatic Transformation of Paralytic Shellfish Toxins in Marine Organisms. In Harmful Marine Algal Blooms; Lassus, P., Arzul, G., Erard, E., Gentien, P., Marcaillou, C., Eds.; Technique et Documentation-Lavoisier, Intercept: Paris, France, 1995; pp. 475–480. [Google Scholar]
- Doucette, G.; Maneiro, I.; Riveiro, I.; Svensen, C. 22 Phycotoxin pathways in aquatic food webs: Transfer, accumulation and degradation. Ecol. Harmful Algae 2006, 189, 283–295. [Google Scholar]
- Shimizu, Y.; Yoshioka, M. Transformation of paralytic shellfish toxins as demonstrated in scallop homogenates. Science 1981, 212, 547–549. [Google Scholar]
- Sullivan, J.J.; Iwaoka, W.T.; Liston, J. Enzymatic transformation of PSP toxins in the littleneck clam (Protothaca staminea). Biochem. Biophys. Res. Commun. 1983, 114, 465–472. [Google Scholar] [CrossRef]
- Murakami, R.; Yamamoto, K.; Noguchi, T. Toxicity and paralytic shellfish poison composition of three species of bivalves collected in Ibaraki Prefecture, Japan. J. Food Hyg. Soc. Jpn. 1999, 40, 46–54. [Google Scholar] [CrossRef]
- Cho, Y.; Ogawa, N.; Takahashi, M.; Lin, P.H.; Oshima, Y. Purification and characterization of paralytic shellfish toxin–transforming enzyme, sulfocarbamoylase I, from the Japanese bivalve Peronidia venulosa. Biochim. Biophys. Acta 2008, 1784, 1277–1285. [Google Scholar]
- Cembella, D.A.; Shumway, E.S.; Larocque, R. Sequestering and putative transformation of paralytic shellfish toxins by the sea scallop Placopecten magellanicus: Seasonal and spatial scales in natural populations. J. Exp. Mar. Biol. Ecol. 1994, 180, 1–22. [Google Scholar] [CrossRef]
- Haya, K.; Martin, L.J.; Robinson, M.C.S.; Martin, D.J.; Khots, A. Does uptake of Alexandrium fundyense cysts contribute to the levels of PSP toxin found in the sea scallop, Planopecten magellanicus? Harmful Algae 2003, 2, 75–81. [Google Scholar] [CrossRef]
- Wong, K.C.; Hung, P.; Lee, L.H.K.; Mok, T.; Kam, M.K. Effect of steam cooking on distribution of paralytic shellfish toxins in different tissue compartments of scallops Patinopecten yessoensis. Food Chem. 2009, 114, 72–80. [Google Scholar] [CrossRef]
- Jiang, T.; Niu, T.; Xu, X. Transfer and metabolism of paralytic shellfish poisoning from scallop (Chlamys nobilis) to spiny lobster (Panulirus stimpsoni). Toxicon 2006, 48, 998–994. [Google Scholar]
- Band-Schmidt, C.J.; Bustillos-Guzmán, J.J.; López-Cortés, D.J.; Gárate-Lizárraga, I.; Núñez-Vázquez, E.J.; Hernández-Sandoval, F.E. Ecological and Physiological Studies of Gymnodinium catenatum in the Mexican Pacific: A review. Mar. Drugs 2010, 8, 1935–1961. [Google Scholar] [CrossRef]
- Morquecho, M.L.; Lechuga-Devéze, C.H. Dinoflagellates in recent sediments from Bahía Concepción, Gulf of California. Bot. Mar. 2003, 46, 132–141. [Google Scholar] [CrossRef]
- Gárate-Lizárraga, I.; Bustillos-Guzmán, J.J.; Alonso-Rodríguez, R.; Luckas, B. Comparative paralytic shellfish toxin profiles in two marine bivalves during outbreaks of Gymnodinium catenatum (Dinophyceae) in the Gulf of California. Mar. Pollut. Bull. 2004, 48, 378–402. [Google Scholar] [CrossRef]
- de la Garza-Aguilar, J. Intoxicación alimentaria por ingestión de mariscos contaminados. Salud Pública Méx. 1983, 25, 145–150. [Google Scholar]
- Mee, L.D.; Ramírez-Flores, A.; Flores-Verdugo, F.; González-Frías, F. Coastal upwelling and fertility of the Southern Gulf of California: Impact of the 1982–83 ENSO event. Trop. Ocean Atmos. Newsl. 1985, 31, 9–10. [Google Scholar]
- Morey-Gaines, G. Gymnodinium catenatum Graham (Dinophyceae): Morphology and affinities with armoured forms. Phycologia 1982, 21, 154–163. [Google Scholar] [CrossRef]
- Hurst, J.W.; Gilfillan, E.S. Paralytic Shellfish Poisoning in Maine. In Proceedings of the 10th National Shellfish Sanitation Workshop, Hunt Valley, MD, USA, 29–30 June 1977; Wilt, D.S., Ed.; US Department of Health, Education and Welfare, Food and Drug Administration: Washington, DC, USA, 1977; pp. 152–161. [Google Scholar]
- Oshima, Y.; Yasumoto, T.; Kodama, M.; Ogata, T.; Fukuyo, Y.; Matsuura, F. Features of paralytic shellfish poison occurring in Tohoku district. Nippon Suisan Gakkaishi 1982, 48, 525–530. [Google Scholar]
- Takatani, T.; Morita, T.; Anami, A.; Akaeda, H.; Kamijyo, Y.; Tsutsumi, K.; Noguchi, T. Appearance of Gymnodinium catenatum in association with the toxification of bivalves in Kamae, Oita Prefecture, Japan. J. Food Hyg. Soc. Jpn. 1998, 39, 275–280. [Google Scholar] [CrossRef]
- Morquecho, E.L. Coleccion de dinoflagelados marinos CODIMAR. Centro de Investigaciones Biológicas del Noroeste, S.C. La Paz, B.C.S., México, 2008. Available online: http://www.cibnor.mx/es/investigacion/colecciones-biologicas/codimar/entrada (accessed on 25 April 2012).
- Doblin, M.; Blackburn, S.I.; Hallegraeff, G.M. Comparative study of selenium requirements of three phytoplankton species: Gymnodinium catenatum, Alexandrium minutum (Dinophyta) and Chaetoceros cf. tenuissimus (Bacyllariophyta). J. Plankton Res. 1999, 21, 1153–1169. [Google Scholar] [CrossRef]
- Rodríguez-Jaramillo, C.; Hurtado, M.A.; Romero-Vivas, E.; Ramírez, J.L.; Manzano, M.; Palacios, E. Gonadal development and histochemistry of the tropical oyster Crassostrea corteziensis (Hertlein, 1951) during an annual reproductive cycle. J. Shellfish Res. 2008, 27, 1129–1141. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Escobedo-Lozano, A.Y.; Estrada, N.; Ascencio, F.; Contreras, G.; Alonso-Rodriguez, R. Accumulation, Biotransformation, Histopathology and Paralysis in the Pacific Calico Scallop Argopecten ventricosus by the Paralyzing Toxins of the Dinoflagellate Gymnodinium catenatum. Mar. Drugs 2012, 10, 1044-1065. https://0-doi-org.brum.beds.ac.uk/10.3390/md10051044
Escobedo-Lozano AY, Estrada N, Ascencio F, Contreras G, Alonso-Rodriguez R. Accumulation, Biotransformation, Histopathology and Paralysis in the Pacific Calico Scallop Argopecten ventricosus by the Paralyzing Toxins of the Dinoflagellate Gymnodinium catenatum. Marine Drugs. 2012; 10(5):1044-1065. https://0-doi-org.brum.beds.ac.uk/10.3390/md10051044
Chicago/Turabian StyleEscobedo-Lozano, Amada Y., Norma Estrada, Felipe Ascencio, Gerardo Contreras, and Rosalba Alonso-Rodriguez. 2012. "Accumulation, Biotransformation, Histopathology and Paralysis in the Pacific Calico Scallop Argopecten ventricosus by the Paralyzing Toxins of the Dinoflagellate Gymnodinium catenatum" Marine Drugs 10, no. 5: 1044-1065. https://0-doi-org.brum.beds.ac.uk/10.3390/md10051044