
A New Member of the TBC1D15 Family from Chiloscyllium plagiosum: Rab GTPase-Activating Protein Based on Rab7 as a Substrate

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Blast Analysis of the Shark TBC1D15 Genome Sequence

2.2. Homology Analysis of the Shark TBC1D15 Amino Acid Sequence

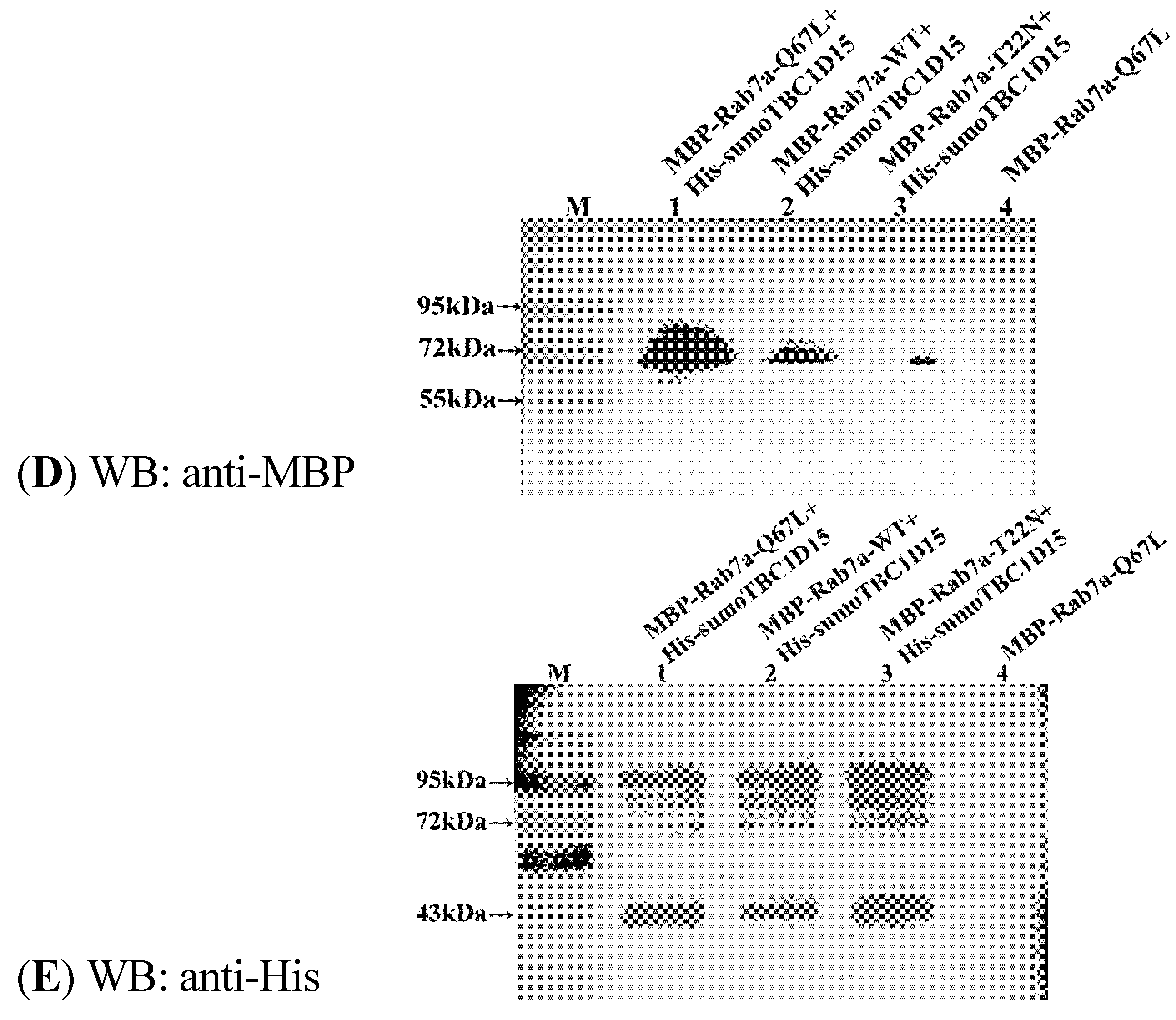
2.3. Shark TBC1D15 Interacts with Rab7 in Vitro


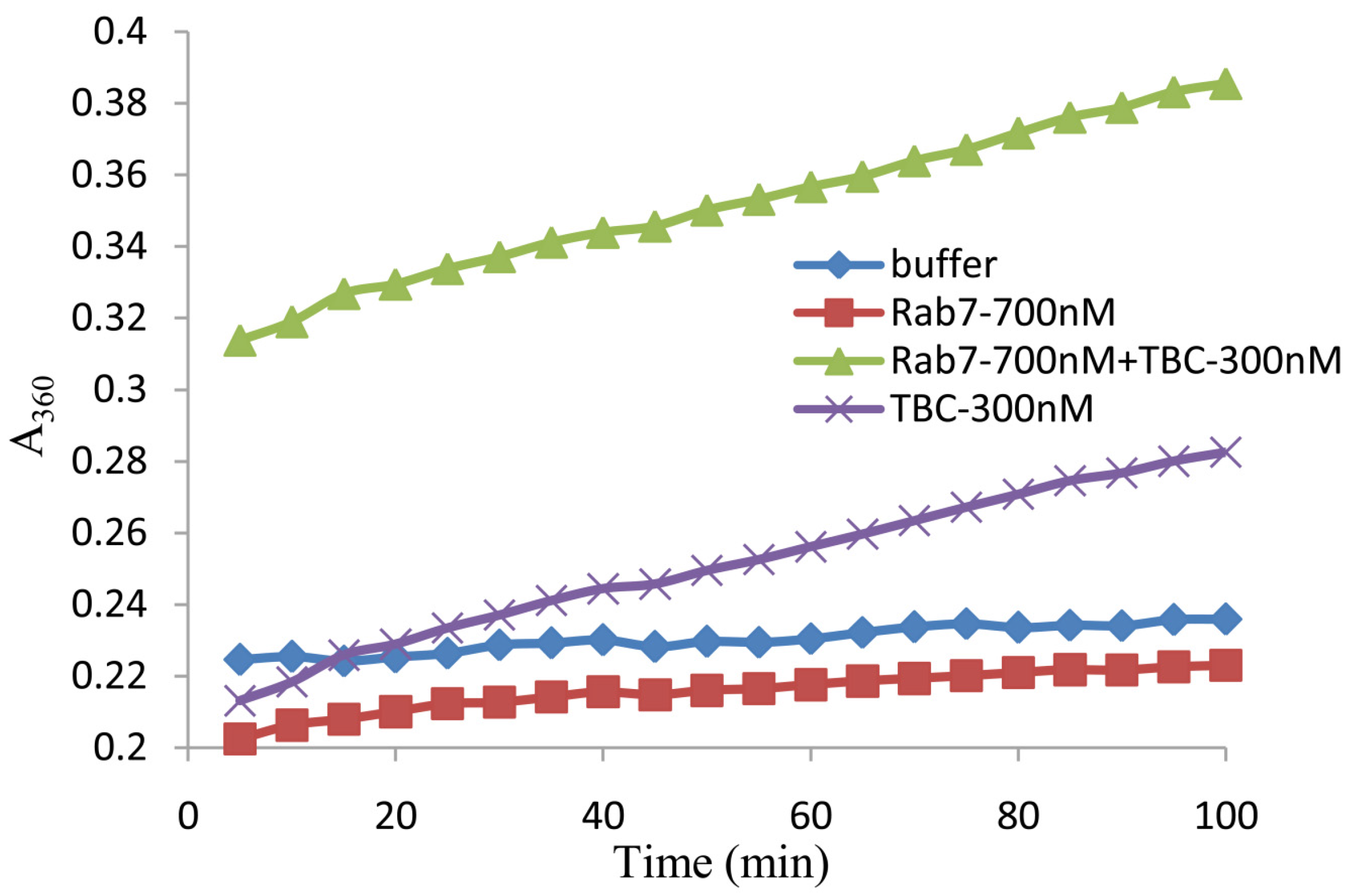
2.4. In Vitro Analysis of the GAP Activity of Shark TBC1D15

3. Discussion
4. Experimental Methods
4.1. Construction of the Expression Vector
- Upstream: 5′-CACGGATCCATGCTGGTGGGTCCGATCGG-3′
- Downstream: 5′-CCGCGGCCGCTTACCACTTGAAGCTCTTCTTCTTG-3′
- Pf1-Q67L: 5′-GTCACAATGCAGATATGGGACACAGCAGGACTGGAACGG-3′
- Pr1-Q67L: 5′-GCAGGACTGGAACGGTTCCAGTCTCTCGGTGTGGCCTTC-3′
- Pf2-T22N: 5′-GGAGATTCTGGAGTCGGGAAGAACTCACTC-3′
- Pr2-T22N: 5′-GGGAAGAACTCACTCATGAACCAGTATGTG-3′
4.2. Purification of Recombinant Proteins
4.3. Protein-Protein Interaction Assays in Vitro (Pull-Down Assays)
4.4. GTPase Activity Assay
5. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Ortis, F.; Naamane, N.; Flamez, D.; Ladriere, L.; Moore, F.; Cunha, D.A.; Colli, M.L.; Thykjaer, T.; Thorsen, K.; Orntoft, T.F.; et al. Cytokines interleukin-1beta and tumor necrosis factor-alpha regulate different transcriptional and alternative splicing networks in primary beta-cells. Diabetes 2010, 59, 358–374. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhao, M.Y.; An, W. Increased hepatic apoptosis in high-fat diet-induced NASH in rats may be associated with downregulation of hepatic stimulator substance. J. Mol. Med. 2011, 89, 1207–1217. [Google Scholar] [CrossRef]
- LaBrecque, D.R.; Steele, G.; Fogerty, S.; Wilson, M.; Barton, J. Purification and physical-chemical characterization of hepatic stimulator substance. Hepatology 1987, 7, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.J.; Lv, Z.B.; Li, Q.; Wei, L.J.; Zhang, L.; Wu, W.T. Study on hepatoprotective effect of peptide S-8300 from shark liver. World J. Gastroenterol. 2005, 11, 1809–1812. [Google Scholar] [CrossRef] [PubMed]
- OU, Y.; Li, Q. Purification and characterization of hepatocyte regeneration stimulatory factor from shark liver. J. Chin. Pharm. Sci. 2003, 12, 175–180. [Google Scholar]
- Lv, Z.; Ou, Y.; Li, Q.; Zhang, W.; Ye, B.; Wu, W. Expression, purification and bioactivities analysis of recombinant active peptide from shark liver. Mar. Drugs 2009, 7, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, Y.; Chen, J.; Zhang, W.; Sheng, Q.; Chen, J.; Yu, W.; Nie, Z.; Zhang, Y.; Wu, W.; et al. A shark liver gene-derived active peptide expressed in the silkworm, Bombyx mori: Preliminary studies for oral administration of the recombinant protein. Mar. Drugs 2013, 11, 1492–1505. [Google Scholar] [CrossRef]
- Huang, F.J.; Wu, T. Purification and characterization of a new peptide (S-8300) from shark liver. J. Food Biochem. 2010, 34, 962–970. [Google Scholar] [CrossRef]
- Huang, F.; Wu, W. Antidiabetic effect of a new peptide from Squalus mitsukurii liver (S-8300) in streptozocin-induced diabetic mice. J. Pharm. Pharmacol. 2005, 57, 1575–1580. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Wu, W. Antidiabetic effect of a new peptide from Squalus mitsukurii liver (S-8300) in alloxan-diabetes. Clin. Exp. Pharmacol. Physiol. 2005, 32, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Gao, Z.; Guo, Q.; Wang, T.; Lu, C.; Chen, Y.; Sheng, Q.; Chen, J.; Nie, Z.; Zhang, Y.; et al. Anti-diabetic effects of CTB-APSL fusion protein in type 2 diabetic mice. Mar. Drugs 2014, 12, 1512–1529. [Google Scholar] [CrossRef] [PubMed]
- Neuwald, A.F. A shared domain between a spindle assembly checkpoint protein and Ypt Rab-specific GTPase-activators. Trends Biochem. Sci. 1997, 22, 243–244. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, M. TBC proteins: GAPs for mammalian small GTPase Rab? Biosci. Rep. 2011, 31, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Walsh, B.; Mitchell, C.A.; Rowe, T. TBC domain family, member 15 is a novel mammalian Rab GTPase-activating protein with substrate preference for Rab7. Biochem. Biophys. Res. Commun. 2005, 335, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Vind, B.F.; Pehmoller, C.; Treebak, J.T.; Birk, J.B.; Hey-Mogensen, M.; Beck-Nielsen, H.; Zierath, J.R.; Wojtaszewski, J.F.; Hojlund, K. Impaired insulin-induced site-specific phosphorylation of TBC1 domain family, member 4 (TBC1D4) in skeletal muscle of type 2 diabetes patients is restored by endurance exercise-training. Diabetologia 2011, 54, 157–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, H.; Kane, S.; Sano, E.; Miinea, C.P.; Asara, J.M.; Lane, W.S.; Garner, C.W.; Lienhard, G.E. Insulin-stimulated phosphorylation of a Rab GTPase-activating protein regulates GLUT4 translocation. J. Biol. Chem. 2003, 278, 14599–14602. [Google Scholar] [CrossRef] [PubMed]
- Cartee, G.D. Roles of TBC1D1 and TBC1D4 in insulin- and exercise-stimulated glucose transport of skeletal muscle. Diabetologia 2015, 58, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Gabernet-Castello, C.; O’Reilly, A.J.; Dacks, J.B.; Field, M.C. Evolution of Tre-2/Bub2/Cdc16 (TBC) Rab GTPase-activating proteins. Mol. Biol. Cell 2013, 24, 1574–1583. [Google Scholar] [CrossRef]
- Mellouk, N.; Weiner, A.; Aulner, N.; Schmitt, C.; Elbaum, M.; Shorte, S.L.; Danckaert, A.; Enninga, J. Shigella subverts the host recycling compartment to rupture its vacuole. Cell Host Microbe 2014, 16, 517–530. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Eathiraj, S.; Munson, M.; Lambright, D.G. TBC-domain GAPs for Rab GTPases accelerate GTP hydrolysis by a dual-finger mechanism. Nature 2006, 442, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Albert, S.; Will, E.; Gallwitz, D. Identification of the catalytic domains and their functionally critical arginine residues of two yeast GTPase-activating proteins specific for Ypt/Rab transport GTPases. EMBO J. 1999, 18, 5216–5225. [Google Scholar] [CrossRef] [PubMed]
- Peralta, E.R.; Martin, B.C.; Edinger, A.L. Differential effects of TBC1D15 and mammalian Vps39 on Rab7 activation state, lysosomal morphology, and growth factor dependence. J. Biol. Chem. 2010, 285, 16814–16821. [Google Scholar] [CrossRef] [PubMed]
- Feldman, D.E.; Chen, C.; Punj, V.; Machida, K. The TBC1D15 oncoprotein controls stem cell self-renewal through destabilization of the Numb-p53 complex. PLoS ONE 2013, 8, e57312. [Google Scholar] [CrossRef]
- Onoue, K.; Jofuku, A.; Ban-Ishihara, R.; Ishihara, T.; Maeda, M.; Koshiba, T.; Itoh, T.; Fukuda, M.; Otera, H.; Oka, T.; et al. Fis1 acts as a mitochondrial recruitment factor for TBC1D15 that is involved in regulation of mitochondrial morphology. J. Cell Sci. 2012, 126, 176–185. [Google Scholar] [CrossRef] [PubMed]
- Zaid, H.; Antonescu, C.N.; Randhawa, V.K.; Klip, A. Insulin action on glucose transporters through molecular switches, tracks and tethers. Biochem. J. 2008, 413, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Larance, M.; Ramm, G.; Stockli, J.; van Dam, E.M.; Winata, S.; Wasinger, V.; Simpson, F.; Graham, M.; Junutula, J.R.; Guilhaus, M.; et al. Characterization of the role of the Rab GTPase-activating protein AS160 in insulin-regulated GLUT4 trafficking. J. Biol. Chem. 2005, 280, 37803–37813. [Google Scholar] [CrossRef] [PubMed]
- Holman, G.D.; Sakamoto, K. Regulating the motor for GLUT4 vesicle traffic. Cell Metab. 2008, 8, 344–346. [Google Scholar] [CrossRef] [PubMed]
- Ishikura, S.; Koshkina, A.; Klip, A. Small G proteins in insulin action: Rab and Rho families at the crossroads of signal transduction and GLUT4 vesicle traffic. Acta Physiol. 2008, 192, 61–74. [Google Scholar] [CrossRef]
- Peck, G.R.; Chavez, J.A.; Roach, W.G.; Budnik, B.A.; Lane, W.S.; Karlsson, H.K.; Zierath, J.R.; Lienhard, G.E. Insulin-stimulated phosphorylation of the Rab GTPase-activating protein TBC1D1 regulates GLUT4 translocation. J. Biol. Chem. 2009, 284, 30016–30023. [Google Scholar] [CrossRef] [PubMed]
- O’Gorman, D.J.; Karlsson, H.K.; McQuaid, S.; Yousif, O.; Rahman, Y.; Gasparro, D.; Glund, S.; Chibalin, A.V.; Zierath, J.R.; Nolan, J.J. Exercise training increases insulin-stimulated glucose disposal and GLUT4 (SLC2A4) protein content in patients with type 2 diabetes. Diabetologia 2006, 49, 2983–2992. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, K.; Holman, G.D. Emerging role for AS160/TBC1D4 and TBC1D1 in the regulation of GLUT4 traffic. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E29–E37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liu, Y.; Zhang, X.; Pan, J.; Nie, Z.; Zhang, W.; Yu, W.; Chen, J.; Liu, L.; Li, J.; et al. The identification of microRNAs in the whitespotted bamboo shark (Chiloscyllium plagiosum) liver by Illumina sequencing. Gene 2013, 527, 259–265. [Google Scholar] [CrossRef]
- Lu, C.; Zhang, J.; Nie, Z.; Chen, J.; Zhang, W.; Ren, X.; Yu, W.; Liu, L.; Jiang, C.; Zhang, Y.; et al. Study of microRNAs related to the liver regeneration of the whitespotted bamboo shark, Chiloscyllium plagiosum. BioMed. Res. Int. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.; Rodriguez, A.D.; Taglialatela-Scafati, O.; Fusetani, N. Marine pharmacology in 2009–2011: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Mar. Drugs 2013, 11, 2510–2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Y.; Mi, Y.J.; Dai, Y.K.; Fu, H.L.; Cui, D.X.; Jin, W.L. The inverse F-BAR domain protein srGAP2 acts through srGAP3 to modulate neuronal differentiation and neurite outgrowth of mouse neuroblastoma cells. PLoS ONE 2013, 8, e57865. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Wang, W.; Cheng, D.; Wang, T.; Lu, C.; Chen, J.; Nie, Z.; Zhang, W.; Lv, Z.; Wu, W.; et al. A New Member of the TBC1D15 Family from Chiloscyllium plagiosum: Rab GTPase-Activating Protein Based on Rab7 as a Substrate. Mar. Drugs 2015, 13, 2955-2966. https://0-doi-org.brum.beds.ac.uk/10.3390/md13052955
Li Y, Wang W, Cheng D, Wang T, Lu C, Chen J, Nie Z, Zhang W, Lv Z, Wu W, et al. A New Member of the TBC1D15 Family from Chiloscyllium plagiosum: Rab GTPase-Activating Protein Based on Rab7 as a Substrate. Marine Drugs. 2015; 13(5):2955-2966. https://0-doi-org.brum.beds.ac.uk/10.3390/md13052955
Chicago/Turabian StyleLi, Yuanyuan, Weidong Wang, Dandan Cheng, Tao Wang, Conger Lu, Jian Chen, Zuoming Nie, Wenping Zhang, Zhengbing Lv, Wutong Wu, and et al. 2015. "A New Member of the TBC1D15 Family from Chiloscyllium plagiosum: Rab GTPase-Activating Protein Based on Rab7 as a Substrate" Marine Drugs 13, no. 5: 2955-2966. https://0-doi-org.brum.beds.ac.uk/10.3390/md13052955