Recent Trends in Marine Phycotoxins from Australian Coastal Waters


Abstract
:1. Introduction
2. Paralytic Shellfish Toxins
3. Diarrhetic Shellfish Toxins
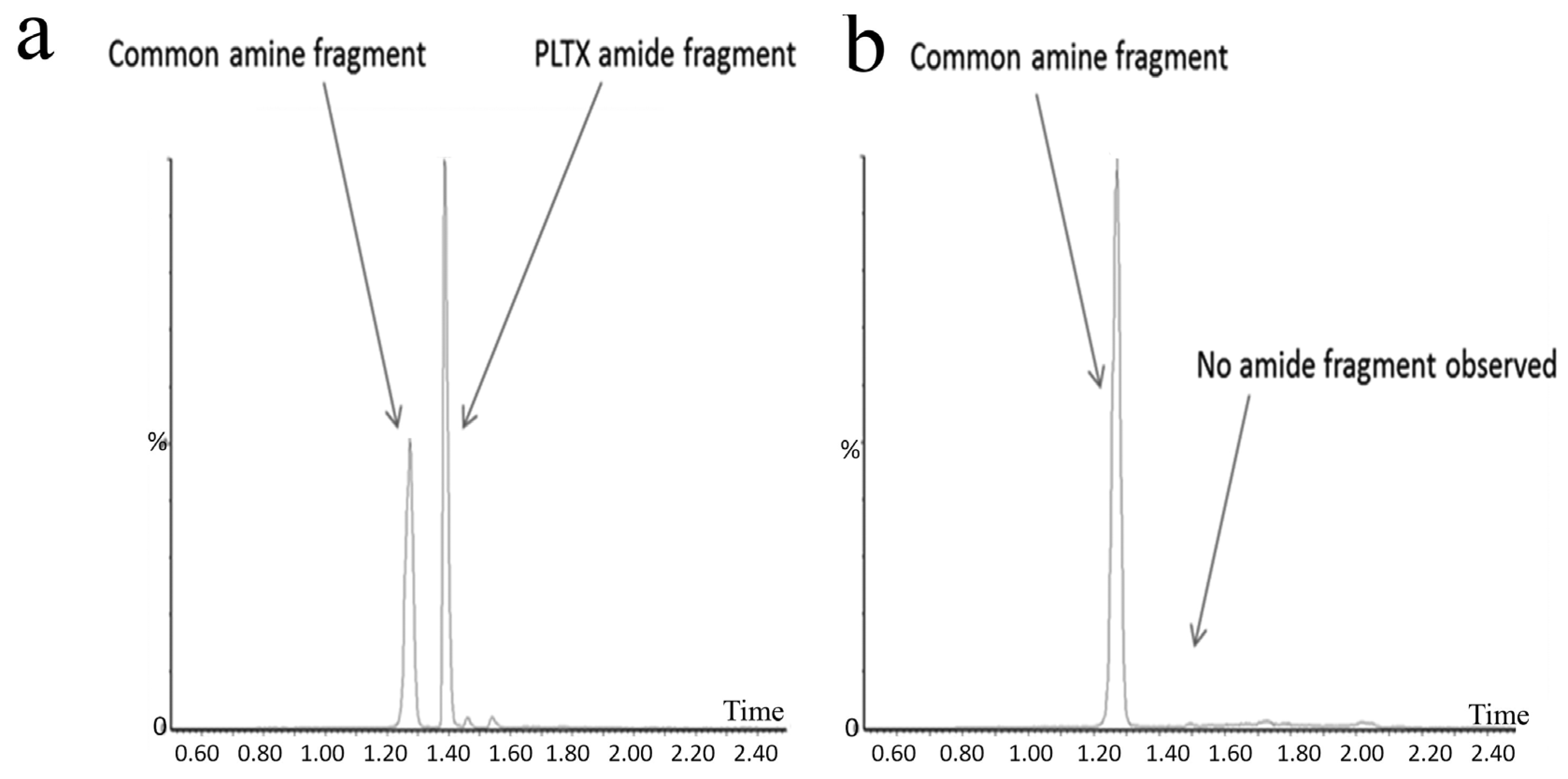
4. Palytoxins
5. Ciguatoxins and Maitotoxins
6. Tetrodotoxins
7. Fish Killing Toxins
7.1. Karlotoxins
7.2. Amphidinolides and Other Fish Killing Toxins
8. Domoic Acid
9. The Use of Molecular Genetic or ELISA Methods to Detect HABs
10. Knowledge Gaps and Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Oceans Policy Science Advisory Group. Marine Nation 2025; Australian Government: Canberra, Australia, 2013; p. 33.
- Food and Agricultural Organization of the United Nations. The State of World Fisheries and Aquaculture 2008; FAO Fisheries and Aquaculture Department: Rome, Italy, 2009; p. 18. [Google Scholar]
- Australian Seafood CRC. Marine biotoxin diagnostic capability and capacity in Australia: A business review and implementation plan. In A Report to the Seafood Cooperative Research Centre Regarding Australia’s Existing and Future Capability and Capacity; Ridge Partners for Australian Seafood CRC: Canberra, Australia, 2011; p. 105. [Google Scholar]
- Campbell, A.; Hudson, D.; McLeod, C.; Nicholls, C.; Pointon, A. Tactical Research Fund: Review of the 2012 paralytic shellfish toxin event in Tasmania associated with the dinoflagellate alga, Alexandrium tamarense. In FRDC Project 2012/060 Appendix to the Final Report; SafeFish: Adelaide Australia, 2013; p. 93. [Google Scholar]
- Kantiani, L.; Llorca, M.; Sanchis, J.; Farre, M.; Barcelo, D. Emerging food contaminants: A review. Anal. Bioanal. Chem. 2010, 398, 2413–2427. [Google Scholar] [CrossRef] [PubMed]
- Ajani, P.; Murray, S.A. A Review of Toxic Algal Species towards Improving Management of Toxic Blooms in New South Wales; A Report to Hornsby Shire Council: New South Wales, Australia, 2016; p. 69. [Google Scholar]
- Ajani, P.; Murray, S.; Hallegraeff, G.; Lundholm, N.; Gillings, M.; Brett, S.; Armand, L. The diatom genus Pseudo-nitzschia (Bacillariophyceae) in New South Wales, Australia: Morphotaxonomy, molecular phylogeny, toxicity, and distribution. J. Phycol. 2013, 49, 765–785. [Google Scholar] [CrossRef] [PubMed]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.F.; Niedzwiadek, B.; Menard, C. Quantitative determination of paralytic shellfish poisoning toxins in shellfish using prechromatographic oxidation and liquid chromatography with fluorescence detection: Collaborative study. J. Aoac Int. 2005, 88, 1714–1732. [Google Scholar] [PubMed]
- Van de Riet, J.; Gibbs, R.S.; Muggah, P.M.; Rourke, W.A.; MacNeil, J.D.; Quilliam, M.A. Liquid chromatography post-column oxidation (PCOX) method for the determination of paralytic shellfish toxins in mussels, clams, oysters, and scallops: Collaborative study. J. Aoac Int. 2011, 94, 1154–1176. [Google Scholar] [PubMed]
- John, U.; Litaker, R.W.; Montresor, M.; Murray, S.; Brosnahan, M.L.; Anderson, D.M. Formal revision of the Alexandrium tamarense species complex (Dinophyceae) taxonomy: The introduction of five species with emphasis on molecular-based (rDNA) classification. Protist 2014, 165, 779–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, S.A.; Wiese, M.; Neilan, B.A.; Orr, R.J.S.; de Salas, M.; Brett, S.; Hallegraeff, G. A reinvestigation of saxitoxin production and sxtA in the ‘non-toxic’ Alexandrium tamarense Group V clade. Harmful Algae 2012, 18, 96–104. [Google Scholar] [CrossRef]
- Farrell, H.; Brett, S.; Ajani, P.; Murray, S. Distribution of the genus Alexandrium (Halim) and paralytic shellfish toxins along the coastline of New South Wales, Australia. Mar. Poll. Bull. 2013, 72, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Bolch, C.J.S.; de Salas, M.F. A review of the molecular evidence for ballast water introduction of the toxic dinoflagellates Gymnodinium catenatum and the Alexandrium “tamarensis complex” to Australasia. Harmful Algae 2007, 6, 465–485. [Google Scholar] [CrossRef]
- FRDC. Improved Understanding of Tasmanian Harmful Algal Blooms and Biotoxin Events to Support Seafood Risk Management; Milestone Progress Report, Project Number 2014/032; Fisheries Research and Development Corporation: Canberra, Australia, 2015; p. 11. [Google Scholar]
- Ajani, P.; Brett, S.; Krogh, M.; Scanes, P.; Webster, G.; Armand, L. The risk of harmful algal blooms (HABs) in the oyster-growing estuaries of New South Wales, Australia. Environ. Monit. Assess. 2013, 185, 5295–5316. [Google Scholar] [CrossRef] [PubMed]
- Hallegraeff, G.M.; Bolch, C.J.; Blackburn, S.I.; Oshima, Y. Species of the toxigenic dinoflagellate genus Alexandrium in southeastern Australian waters. Bot. Mar. 1991, 34, 575–587. [Google Scholar] [CrossRef]
- McMinn, A.; Hallegraeff, G.M.; Thomson, P.; Jenkinson, A.V.; Heijnis, H. Cyst and radionucleotide evidence for the recent introduction of the toxic dinoflagellate Gymnodinium catenatum into Tasmanian waters. Mar. Ecol. Progr. Ser. 1997, 161, 165–172. [Google Scholar] [CrossRef]
- Hallegraeff, G.M.; Sumner, C.E. Toxic plankton blooms affect shellfish farms. Aust. Fish. 1986, 45, 15–18. [Google Scholar]
- McLeod, C.; Dowsett, N.; Hallegraeff, G.; Harwood, D.T.; Hay, B.; Ibbott, S.; Malhi, N.; Murray, S.; Smith, K.; Tan, J. Accumulation and depuration of paralytic shellfish toxins by Australian abalone Haliotis rubra: Conclusive association with Gymnodinium catenatum dinoflagellate blooms. Food Control 2016. in review. [Google Scholar]
- Negri, A.P.; Bolch, C.J.S.; Geier, S.; Green, D.H.; Park, T.G.; Blackburn, S.I. Widespread presence of hydrophobic paralytic shellfish toxins in Gymnodinium catenatum. Harmful Algae 2007, 6, 774–780. [Google Scholar] [CrossRef]
- Hallegraeff, G.M.; Fraga, S. Bloom dynamics of the toxic dinoflagellate Gymnodinium catenatum, with emphasis on Tasmanian and Spanish coastal waters. In Physiological Ecology of Harmful Algal Blooms; Anderson, D.M., Cembella, A.D., Hallegraeff, G.M., Eds.; Springer: Berlin, Germany, 1998; pp. 59–80. [Google Scholar]
- Reguera, B.; Riobo, P.; Rodriguez, F.; Diaz, P.A.; Pizarro, G.; Paz, B.; Franco, J.M.; Blanco, J. Dinophysis toxins: Causative organisms, distribution and fate in shellfish. Mar. Drugs 2014, 12, 394–461. [Google Scholar] [CrossRef] [PubMed]
- Reguera, B.; Velo-Suárez, L.; Raine, R.; Park, M.G. Harmful Dinophysis species: A review. Harmful Algae 2012, 14, 87–106. [Google Scholar] [CrossRef]
- Simoes, E.; Vieira, R.C.; Schramm, M.A.; Mello, D.F.; Pontinha, V.D.A.; Da Silva, P.M.; Barracco, M.A. Impact of harmful algal blooms (Dinophysis acuminata) on the immune system of oysters and mussels from Santa Catarina, Brazil. J. Mar. Biol. Assoc. UK 2015, 95, 773–781. [Google Scholar] [CrossRef]
- Carmody, E.P.; James, K.J.; Kelly, S.S. Dinophysistoxin-2: The predominant diarrhoetic shellfish toxin in Ireland. Toxicon 1996, 34, 351–359. [Google Scholar] [CrossRef]
- Traore, A.; Baudrimont, I.; Ambaliou, S.; Dano, S.D.; Creppy, E.E. DNA breaks and cell cycle arrest induced by okadaic acid in Caco-2 cells, a human colonic epithelial cell line. Arch. Toxicol. 2001, 75, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Wallace, G.M. Diarrhetic Shellfish Toxins in Tasmanian Coastal Waters: Causative Dinoflagellate Organisms, Dissolved Toxins and Shellfish Depuration; University of Tasmania: Tasmania, Australia, 2011. [Google Scholar]
- Miles, C.O.; Wilkins, A.L.; Munday, R.; Dines, M.H.; Hawkes, A.D.; Briggs, L.R.; Sandvik, M.; Jensen, D.J.; Cooney, J.M.; Holland, P.T.; et al. Isolation of pectenotoxin-2 from Dinophysis acuta and its conversion to pectenotoxin-2 seco acid, and preliminary assessment of their acute toxicities. Toxicon 2004, 43, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.; McIntyre, L.; Ritson, M.; Stone, J.; Bronson, R.; Bitzikos, O.; Rourke, W.; Galanis, E.; Outbreak Investigation Team. Outbreak of diarrhetic shellfish poisoning associated with mussels, British Columbia, Canada. Mar. Drugs 2013, 11, 1669–1676. [Google Scholar] [CrossRef] [PubMed]
- Lembeye, G.; Yasumoto, Y.; Zhao, J.; Fernández, R. DSP Outbreak in Chilean Fjords. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 525–529. [Google Scholar]
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a new type of shellfish poisoning in the Tohoku district. Bull. Jpn. Soc. Fish. Sci. 1978, 44, 1249–1255. [Google Scholar] [CrossRef]
- Quaine, J.; Kraa, E.; Holloway, J.; White, K.; McCarthy, R.; Delpech, V.; Trent, M.; McAnulty, J. Outbreak of gastroenteritis linked to eating pipis. N. S. W. Pub. Health Bull. 1997, 8, 103–104. [Google Scholar]
- MacKenzie, L.; Holland, P.; McNabb, P.; Beuzenberg, V.; Selwood, A.; Suzuki, T. Complex toxin profiles in phytoplankton and Greenshell mussels (Perna canaliculus), revealed by LC-MS/MS analysis. Toxicon 2002, 40, 1321–1330. [Google Scholar] [CrossRef]
- Madigan, T.L.; Lee, K.G.; Padula, D.J.; McNabb, P.; Pointon, A.M. Diarrhetic shellfish poisoning (DSP) toxins in South Australian shellfish. Harmful Algae 2006, 5, 119–123. [Google Scholar] [CrossRef]
- Van Dolan, F.M.; Fire, S.E.; Leighfield, T.A.; Mikulski, C.M.; Doucette, G.J. Determination of paralytic phellfish toxins in shellfish by receptor binding assay: Collaborative study. J. AOAC Int. 2012, 95, 795–812. [Google Scholar] [CrossRef]
- Turner, A.D.; McNabb, P.S.; Harwood, D.T.; Selwood, A.I.; Boundy, M.J. Single-Laboratory Validation of a Multitoxin Ultra-Performance LC-Hydrophilic Interaction LC-MS/MS Method for Quantitation of Paralytic Shellfish Toxins in Bivalve Shellfish. J. Aoac Int. 2015, 98, 609–621. [Google Scholar] [PubMed]
- McNabb, P.; Selwood, A.I.; Holland, P.T. Multiresidue method for determination of algal toxins in shellfish: Single-laboratory validation and interlaboratory study. J. Aoac Int. 2005, 88, 761–772. [Google Scholar] [PubMed]
- Van den Top, H.J.; Gerssen, A.; McCarron, P.; van Egmond, H.P. Quantitative determination of marine lipophilic toxins in mussels, oysters and cockles using liquid chromatography-mass spectrometry: Inter-laboratory validation study. Food Add. Cont. Part A 2011, 28, 1745–1757. [Google Scholar] [CrossRef] [PubMed]
- Smienk, H.; Dominguez, E.; Rodriguez-Velasco, M.L.; Clarke, D.; Kapp, K.; Katikou, P.; Cabado, A.G.; Otero, A.; Vieites, J.M.; Razquin, P.; et al. Quantitative determination of the okadaic acid toxins group by a colorimetric phosphatase inhibition assay: Interlaboratory study. J. Aoac Int. 2013, 96, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Iacovo, E.D.; Fattorusso, E.; Forino, M.; Tartaglione, L. LC-MS of palytoxin and its analogues: State of the art and future perspectives. Toxicon 2011, 57, 376–389. [Google Scholar] [CrossRef] [PubMed]
- Selwood, A.I.; van Ginkel, R.; Harwood, D.T.; McNabb, P.S.; Rhodes, L.R.; Holland, P.T. A sensitive assay for palytoxins, ovatoxins and ostreocins using LC-MS/MS analysis of cleavage fragments from micro-scale oxidation. Toxicon 2012, 60, 810–820. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Hoppenrath, M.; Dorantes-Aranda, J.J.; Harwood, D.T.; Murray, S.A. Molecular and phylogenetic characterization of Ostreopsis (Dinophyceae) and the description of a new species, Ostreopsis rhodesae sp. nov. from a subtropical Australian lagoon. Harmful Algae 2016, 60, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Yogi, K.; Oshiro, N.; Inafuku, Y.; Hirama, M.; Yasumoto, T. Detailed LC-MS/MS Analysis of ciguatoxins revealing distinct regional and species characteristics in fish and causative alga from the Pacific. Anal. Chem. 2011, 83, 8886–8891. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.J.; Mak, Y.L.; Murphy, M.B.; Lam, J.C.W.; Chan, W.H.; Wang, M.; Chan, L.L.; Lam, P.K.S. Validation of an accelerated solvent extraction liquid chromatography–tandem mass spectrometry method for Pacific ciguatoxin-1 in fish flesh and comparison with the mouse neuroblastoma assay. Anal. Bioanal. Chem. 2011, 400, 3165–3175. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Jester, E.L.E.; Granade, H.R.; Plakas, S.M.; Dickey, R.W. Caribbean ciguatoxin profile in raw and cooked fish implicated in ciguatera. Food Chem. 2012, 131, 192–198. [Google Scholar] [CrossRef]
- Yogi, K.; Sakugawa, S.; Oshiro, N.; Ikehara, T.; Sugiyama, K.; Yasumoto, T. Determination of toxins involved in ciguatera fish poisoning in the Pacific by LC/MS. J. Aoac Int. 2014, 97, 398–402. [Google Scholar] [CrossRef] [PubMed]
- Darius, H.T.; Ponton, D.; Revel, T.; Cruchet, P.; Ung, A.; Fouc, M.T.; Chinain, M. Ciguatera risk assessment in two toxic sites of French Polynesia using the receptor-binding assay. Toxicon 2007, 50, 612–626. [Google Scholar] [CrossRef] [PubMed]
- Hardison, D.R.; Holland, W.C.; McCall, J.R.; Bourdelais, A.J.; Baden, D.G.; Darius, H.T.; Chinain, M.; Tester, P.A.; Shea, D.; Quintana, H.A.F.; et al. Fluorescent receptor binding assay for detecting ciguatoxins in fish. PLoS ONE 2016. [Google Scholar] [CrossRef] [PubMed]
- Caillaud, A.; Eixarch, H.; de la Iglesia, P.; Rodriguez, M.; Dominguez, L.; Andree, K.B.; Diogene, J. Towards the standardisation of the neuroblastoma (neuro-2a) cell-based assay for ciguatoxin-like toxicity detection in fish: Application to fish caught in the Canary Islands. Food Add. Cont. A 2012, 29, 1000–1010. [Google Scholar] [CrossRef] [PubMed]
- Naritia, H.; Noguchi, T.; Maruyama, J.; Ueda, Y.; Hashimoto, K.; Watanabe, Y.; Hida, K. Occurrence of tetrodotoxin in a trumpet shellfish “boshubora” Charonia sauliae. Nippon Suisan Gakkaishi 1981, 47, 934–941. [Google Scholar]
- Nagashima, Y.J.; Maruyama, T.; Noguchi, T.; Hashimoto, K. Analysis of paralytic shellfish poison and tetrodotoxin by ion-pair high-performance liquid chromatography. Nippon Suisan Gakkaishi 1987, 53, 819–823. [Google Scholar] [CrossRef]
- McNabb, P.S.; Taylor, D.I.; Ogilvie, S.C.; Wilkinson, L.; Anderson, A.; Hamon, D.; Wood, S.A.; Peaice, B.M. First detection of tetrodotoxin in the bivalve Paphies australis by liquid chromatography coupled to triple quadrupole mass spectrometry with and without precolumn reaction. J. Aoac Int. 2014, 97, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Adolf, J.E.; Bachvaroff, T.R.; Deeds, J.R.; Place, A.R. Ichthyotoxic Karlodinium veneficum (Ballantine) J Larsen in the upper Swan River estuary (Western Australia): Ecological conditions leading to a fish kill. Harmful Algae 2015, 48, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Bachvaroff, T.R.; Adolf, J.E.; Squier, A.H.; Harvey, H.R.; Place, A.R. Characterization and quantification of karlotoxins by liquid chromatography-mass spectrometry. Harmful Algae 2008, 7, 473–484. [Google Scholar] [CrossRef]
- Lapworth, C.; Hallegraeff, G.; Ajani, P. Identification of Domoic-Acid-Producing Pseudo-Nitzschia Species in Australian Water. In Harmful Algal Blooms 2000, Proceedings of the Ninth International Conference on Harmful Algal Blooms, Hobart, Australia, 7–11 February 2000; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 2001; pp. 38–41. [Google Scholar]
- Quilliam, M.A.; Xie, M.; Hardstaff, W.R. Rapid extraction and cleanup for liquid-chromatographic determination of domoic acid in unsalted seafood. J. Aoac Int. 1995, 78, 543–554. [Google Scholar]
- Ajani, P.; Larsson, M.E.; Rubio, A.; Bush, S.; Brett, S.; Farrell, H. Modelling bloom formation of the toxic dinoflagellates Dinophysis acuminata and Dinophysis caudata in a highly modified estuary, south eastern Australia. Estuar. Coast. Shelf Sci. 2016, 183, 95–106. [Google Scholar] [CrossRef]
- Moore, R.E.; Scheuer, P.J. Palytoxin-new marine toxin from a coelenterate. Science 1971, 172, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Grauso, L.; Tartaglione, L.; Florio, C.; Lorenzon, P.; De Bortoli, M.; et al. Stereostructure and Biological Activity of 42-Hydroxy-palytoxin: A new palytoxin analogue from Hawaiian Palythoa subspecies. Chem. Res. Toxicol. 2009, 22, 1851–1859. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.E.; Bartolini, G. Structure of palytoxin. J. Am. Chem. Soc. 1981, 103, 2491–2494. [Google Scholar] [CrossRef]
- Munday, R. Palytoxin toxicology: Animal studies. Toxicon 2011, 57, 470–477. [Google Scholar] [CrossRef] [PubMed]
- Usami, M.; Satake, M.; Ishida, S.; Inoue, A.; Kan, Y.; Yasumoto, T. Palytoxin analogs from the dinoflagellate Ostreopsis siamensis. J. Am. Chem. Soc. 1995, 117, 5389–5390. [Google Scholar] [CrossRef]
- Brissard, C.; Herve, F.; Sibat, M.; Sechet, V.; Hess, P.; Amzil, Z.; Herrenknecht, C. Characterization of ovatoxin-h, a new ovatoxin analog, and evaluation of chromatographic columns for ovatoxin analysis and purification. J. Chromatogr. A 2015, 1388, 87–101. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Battocchi, C.; Crinelli, R.; Carloni, E.; Magnani, M.; et al. Unique toxin profile of a Mediterranean Ostreopsis cf. ovata Sstrain: HR LC-MSn Characterization of Ovatoxin-f, a New Palytoxin Congener. Chem. Res. Toxicol. 2012, 25, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Grillo, C.; Melchiorre, N. Putative palytoxin and its new analogue, ovatoxin-a, in Ostreopsis ovata collected along the Ligurian coasts during the 2006 toxic outbreak. J. Am. Soc. Mass Spectrom. 2008, 19, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Altares, M.; Casanova, A.; Bane, V.; Diogene, J.; Furey, A.; de la Iglesia, P. Confirmation of pinnatoxins and spirolides in shellfish and passive samplers from Catalonia (Spain) by liquid chromatography coupled with triple quadrupole and high-resolution hybrid tandem mass spectrometry. Mar. Drugs 2014, 12, 3706–3732. [Google Scholar] [CrossRef] [PubMed]
- Tartaglione, L.; Mazzeo, A.; Dell’Aversano, C.; Forino, M.; Giussani, V.; Capellacci, S.; Penna, A.; Asnaghi, V.; Faimali, M.; Chiantore, M.; et al. Chemical, molecular, and eco-toxicological investigation of Ostreopsis sp from Cyprus Island: Structural insights into four new ovatoxins by LC-HRMS/MS. Anal. Bioanal. Chem. 2016, 408, 915–932. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, L.; Towers, N.; Briggs, L.; Munday, R.; Adamson, J. Uptake of palytoxin-like compounds by shellfish fed Ostreopsis siamensis (Dinophyceae). N Zeal. J. Mar. Freshw. Res. 2002, 36, 631–636. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Dello Iacovo, E.; Fattorusso, E.; Forino, M.; Tartaglione, L.; Benedettini, G.; Onorari, M.; Serena, F.; Battocchi, C.; et al. First finding of Ostreopsis cf. ovata toxins in marine aerosols. Environ. Sci. Technol. 2014, 48, 3532–3540. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Hoppenrath, M.; Harwood, T.; Brett, S.; Rhodes, L.; Murray, S. Molecular phylogeny, morphology and toxigenicity of Ostreopsis cf. siamensis (Dinophyceae) from temperate south-east Australia. Phycol. Res. 2016, 64, 146–159. [Google Scholar] [CrossRef]
- Fleming, L.E.; Baden, D.G.; Bean, J.A.; Weisman, R.; Blythe, D.G. Seafood toxin diseases: Issues in epidemiology and community outreach. In Harmful Algae; Reguera, B., Blanco, J., Fernandez, M.L., Wyatt, T., Eds.; Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO: Santiago, Spain, 1998; pp. 245–248. [Google Scholar]
- Sims, J.K. Theoretical discourse on the pharmacology of toxic marine injestions. Ann. Emerg. Med. 1987, 16, 1006–1015. [Google Scholar] [CrossRef]
- Kohli, G.S.; Murray, S.A.; Neilan, B.A.; Rhodes, L.L.; Harwood, D.T.; Smith, K.F.; Meyer, L.; Capper, A.; Brett, S.; Hallegraeff, G.M. High abundance of the potentially maitotoxic dinoflagellate Gambierdiscus carpenteri in temperate waters of New South Wales, Australia. Harmful Algae 2014, 39, 134–145. [Google Scholar] [CrossRef]
- Kretzschmar, A.L.; Verma, A.; Harwood, D.T.; Hoppenrath, M.; Murray, S. Characterization of Gambierdiscus lapillus sp. nov. (Gonyaulacales, Dinophyceae): A new toxic dinoflagellate from the Great Barrier Reef (Australia). J. Phycol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.; Momigliano, P.; Heimann, K.; Blair, D. Molecular phylogenetics and morphology of Gambierdiscus yasumotoi from tropical eastern Australia. Harmful Algae 2014, 39, 242–252. [Google Scholar] [CrossRef]
- Chinain, M.; Faust, M.A.; Pauillac, S. Morphology and molecular analyses of three toxic species of Gambierdiscus (Dinophyceae): G. pacificus sp nov. G. australes sp nov. and G. polynesiensis sp nov. J. Phycol. 1999, 35, 1282–1296. [Google Scholar] [CrossRef]
- Fraga, S.; Rodriguez, F.; Caillaud, A.; Diogene, J.; Raho, N.; Zapata, M. Gambierdiscus excentricus sp nov (Dinophyceae), a benthic toxic dinoflagellate from the Canary Islands (NE Atlantic Ocean). Harmful Algae 2011, 11, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Holland, W.C.; Litaker, R.W.; Tomas, C.R.; Kibler, S.R.; Place, A.R.; Davenport, E.D.; Tester, P.A. Differences in the toxicity of six Gambierdiscus (Dinophyceae) species measured using an in vitro human erythrocyte lysis assay. Toxicon 2013, 65, 15–33. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Sellin, M.; Poli, M.A.; Norton, R.S.; MacLeod, J.K.; Sheil, M.M. Purification and characterization of ciguatoxins from moray eel (Lycodontis javanicus, Muraenidae). Toxicon 1991, 29, 1115–1127. [Google Scholar] [CrossRef]
- Nagai, H.; Murata, M.; Torigoe, K.; Satake, M.; Yasumoto, T. Gambieric acids, new potent antifungal substances with unprecedented polyether structures from a marine dinoflagellate Gambierdiscus toxicus. J. Org. Chem. 1992, 57, 5448–5453. [Google Scholar] [CrossRef]
- Satake, M.; Murata, M.; Yasumoto, T. Gambierol—A new toxic polyether compound isolated from the marine dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1993, 115, 361–362. [Google Scholar] [CrossRef]
- Rodriguez, I.; Genta-Jouve, G.; Alfonso, C.; Calabro, K.; Alonso, E.; Sanchez, J.A.; Alfonso, A.; Thomas, O.P.; Botana, L.M. Gambierone, a ladder-shaped polyether from the dinoflagellate Gambierdiscus belizeanus. Org. Lett. 2015, 17, 2392–2395. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.J.; Lewis, R.J. Purification and characterisation of large and small maitotoxins from cultured Gambierdiscus toxicus. Nat. Toxins 1994, 2, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Yasumoto, T. The structure elucidation and biological activities of high molecular weight algal toxins: Maitotoxin, prymnesins and zooxanthellatoxins. Nat. Prod. Rep. 2000, 17, 293–314. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Naoki, H.; Iwashita, T.; Matsunaga, S.; Sasaki, M.; Yokoyama, A.; Yasumoto, T. Structure of maitotoxin. J. Am. Chem. Soc. 1993, 115, 2060–2062. [Google Scholar] [CrossRef]
- Kohli, G.S.; Papiol, G.G.; Rhodes, L.L.; Harwood, D.T.; Selwood, A.; Jerrett, A.; Murray, S.A.; Neilan, B.A. A feeding study to probe the uptake of maitotoxin by snapper (Pagrus auratus). Harmful Algae 2014, 37, 125–132. [Google Scholar] [CrossRef]
- Richlen, M.L.; Morton, S.L.; Barber, P.H.; Lobel, P.S. Phylogeography, morphological variation and taxonomy of the toxic dinoflagellate Gambierdiscus toxicus (Dinophyceae). Harmful Algae 2008, 7, 614–629. [Google Scholar] [CrossRef]
- Sumner, J. Hazards affecting Australian seafood. In Part 2 Supporting Information; SafeFish and the Australian Seafood Cooperative Research Centre: Adelaide, Australia, 2011; p. 46. [Google Scholar]
- Farrell, H.; Zammit, A.; Manning, J.; Shadbolt, C.; Szabo, L.; Harwood, D.T.; McNabb, P.; Turahui, J.A.; van den Berg, D.J. Clinical diagnosis and chemical confirmation of ciguatera fish poisoning in New South Wales, Australia. Commun. Dis. Intell. Q. Rep. 2016, 40, E1–E6. [Google Scholar] [PubMed]
- Murray, S.A.; Farrell, H.; Harwood, T.; Zammit, A. Is ciguatera moving south in Australia? Harmful Algae News 2016, 54, 5–6. [Google Scholar]
- Isbister, G.K.; Son, J.; Wang, F.; Maclean, C.J.; Lin, C.S.Y.; Ujma, J.; Balit, C.R.; Smith, B.; Milder, D.G.; Kiernan, M.C. Puffer fish poisoning: A potentially life-threatening condition. Med. J. Aust. 2002, 177, 650–653. [Google Scholar] [PubMed]
- Kodama, M.; Sato, S.; Ogata, T. Alexandrium tamarense as a source of tetrodotoxin in the scallop Patinopecten yessoensis. In Proceedings of the 5th International Conf on Toxic Marine Phytoplankton, University of Rhode Island, RI, USA, 28 October–1 November 1991.
- Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First detection of tetrodotoxin in Greek shellfish by UPLC-MS/MS potentially linked to the presence of the dinoflagellate Prorocentrum minimum. Toxins 2015, 7, 1779–1807. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the pufferfish toxin tetrodotoxin in European bivalves, England, 2013 to 2014. Eurosurveillance 2015, 20, 2–8. [Google Scholar] [CrossRef]
- Munday, R. Toxicology of seafood toxins: A critical review. In Seafood and Freshwater Toxins: Pharmalogy, Physiology and Detection; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 197–289. [Google Scholar]
- Kodama, M.; Sato, S.; Sakamoto, S.; Ogata, T. Occurrence of tetrodotoxin in Alexandrium tamarense, a causative dinoflagellate of paralytic shellfish poisoning. Toxicon 1996, 34, 1101–1105. [Google Scholar] [CrossRef]
- Place, A.R.; Bowers, H.A.; Bachvaroff, T.R.; Adolf, J.E.; Deeds, J.R.; Sheng, J. Karlodinium veneficum-the little dinoflagellate with a big bite. Harmful Algae 2012, 14, 179–195. [Google Scholar] [CrossRef]
- Hallegraeff, G.M. Aquaculturist’s Guide to Harmful Australian Microalgae, 3rd ed.; School of Plant Science, University of Tasmania: Tasmania, Australia, 2015; p. 150. [Google Scholar]
- Murray, S.A.; Kohli, G.S.; Farrell, H.; Spiers, Z.B.; Place, A.R.; Dorantes-Aranda, J.J.; Ruszczyk, J. A fish kill associated with a bloom of Amphidinium carterae in a coastal lagoon in Sydney, Australia. Harmful Algae 2015, 49, 19–28. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Frame, E.R.; Kendrick, P.S. Domoic acid and fish behavior: A review. Harmful Algae 2012, 13, 126–130. [Google Scholar] [CrossRef]
- Ajani, P.; Kim, J.H.; Han, M.S.; Murray, S.A. The first report of the potentially harmful diatom Pseudo-nitzschia caciantha from Australian coastal waters. Phycol. Res. 2016, 64, 312–317. [Google Scholar] [CrossRef]
- Hallegraeff, G.M. Species of the genus Pseudo-nitzschia in Australian waters. Botanica Marina 1994, 37, 397–411. [Google Scholar] [CrossRef]
- Penna, A.; Galluzzi, L. The quantitative real-time PCR applications in the monitoring of marine harmful algal bloom (HAB) species. Environ. Sci. Pollut. Res. 2013, 20, 6851–6862. [Google Scholar] [CrossRef]
- Medlin, L.K.; Kegel, J.U. Validation of the detection of Pseudo-nitzschia spp. using specific RNA probes tested in a microarray format: Calibration of signal based on variability of RNA content with environmental conditions. Harmful Algae 2014, 37, 183–193. [Google Scholar] [CrossRef]
- Groben, R.; Medlin, L. In situ hybridization of phytoplankton using fluorescently labeled rRNA probes. In Molecular Evolution: Producing the Biochemical Data, Part B; Zimmer, E.A., Roalson, E.H., Eds.; Academic Press: Cambridge, MA, USA, 2005; Volume 395, pp. 299–310. [Google Scholar]
- Garibo, D.; Campbell, K.; Casanova, A.; de la Iglesia, P.; Fernandez-Tejedor, M.; Diogene, J.; Elliott, C.T.; Campas, M. SPR immunosensor for the detection of okadaic acid in mussels using magnetic particles as antibody carriers. Sens. Actuators B-Chem. 2014, 190, 822–828. [Google Scholar] [CrossRef]
- Mcleod, C.; Burrell, S.; Holland, P.T. Review of the Currently Available Field Methods for Detection of Marine Biotoxins in Shellfish Flesh; FS102086; Food Standards Agency and Food Standards Scotland (FSS): Scotland, UK, 2015; p. 86. [Google Scholar]
- McPartlin, D.A.; Lochhead, M.J.; Connell, L.B.; Doucette, G.J.; O’Kennedy, R.J. Use of biosensors for the detection of marine toxins. Essays Biochem. 2016, 60, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Boundy, M.J.; Selwood, A.I.; Harwood, D.T.; McNabb, P.S.; Turner, A.D. Development of a sensitive and selective liquid chromatography-mass spectrometry method for high throughput analysis of paralytic shellfish toxins using graphitised carbon solid phase extraction. J. Chromatogr. A 2015, 1387, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.A.; Wiese, M.; Stuken, A.; Brett, S.; Kellmann, R.; Hallegraeff, G.; Neilan, B.A. sxtA-Based quantitative molecular assay t identify saxitoxin-producing harmful algal blooms in marine waters. Appl. Environ. Microbiol. 2011, 77, 7050–7057. [Google Scholar] [CrossRef] [PubMed]
- Ruvindy, R.; Bolch, C.; Hallegraeff, G.M.; Smith, K.; Van Asten, M.; Murray, S.A. Investigating co-occurring blooms of A. pacificum and A. fundyense using sxtA-based and species specific qPCR assays in eastern Australian waters. In Proceedings of the 17th International Conference on Harmful Algae, Santa Catarina, Brazil, 9–14 October 2016.
- Farrell, H.; O’Connor, W.; Seebacher, F.; Harwood, D.T.; Murray, S. Molecular detection of the sxtA gene from saxitoxin-producing Alexandrium minutum in commercial oysters. J. Shellf. Res. 2016, 35, 169–177. [Google Scholar] [CrossRef]


{kind=link}
| Phycotoxin Group | Chemical Structure (Example of Toxin within Group) | Microalgae | Method of Detection | Reference |
|---|---|---|---|---|
| Paralytic Shellfish Toxins |  Saxitoxin (STX) | Alexandrium fundyense Alexandrium pacificum Alexandrium australiense Alexandrium minutum Gymnodinium catenatum | PSP Mouse bioassay | AOAC.OMA 959.08 |
| HPLC-FLD (pre-column oxidation) | AOAC-OMA 2005.06 [9] | |||
| HPLC-FLD (post-column oxidation) | AOAC.OMA 2011.02 [10] | |||
| Receptor Binding Assay | AOAC.OMA 2011.27 [36] | |||
| LCMS | [37] | |||
| Diarrhetic Shellfish Toxins |  Okadaic acid (OA) | Dinophysis acuminate Dinophysis tripos Dinophysis fortii Dinophysis caudate | LC–MS (acidic mobile phase) | [38] |
| LC-MS (alkaline mobile phase) | [39] | |||
| Pectenotoxins |  Pectenotoxin-2 (PTX-2) | HPLC/MS | [28,35] | |
| Protein Phosphatase Assay PP2A | [40] | |||
| Palytoxins, Ovatoxins and Ostreocins |  Palytoxin (PLTX) | Ostreopsis cf. siamensis Ostreopsis ovata | LC-MS (intact) | [41] |
| LC-MS (substructures) | [42] [43] | |||
| Ciguatoxins |  Ciguatoxin-3C (CTX-3C) | Gambierdiscus spp. G. lapillus sp. nov. | LCMS | [44,45,46,47] |
| Receptor Binding Assay R (radiolabelled) | [48] | |||
| Maitotoxins |  Maitotoxin-1 (MTX) | Receptor Binding Assay F (fluorescent) | [49] | |
| Neuroblastoma Cell Assay (N2A) | [50] | |||
| Tetrodotoxins |  Tetrodotoxin (TTX) | Alexandrium tamarense (?) Prorocentrum minimum (?) | GC-MS | [51] |
| LC-FLD | [52] | |||
| LC-MS | [53] | |||
| Karlotoxins |  Karlotoxin-1 (KmTx-1) | Karlodinium veneficum (synonym: Karlodinium micrum, Gymnodinium veneficum) | LC-MS/MS | [54] |
| HPLC-MS | [55] | |||
| Domoic Acid |  Domoic acid (DA) | Pseudo-nitzschia australis Pseudo-nitzschia cuspidate Pseudo-nitzschia multistriata | ELISA | AOAC-OMA 2006.02 [56] |
| LC-MS/MS | [7] | |||
| HPLC-UV | [57] |
| Sample (All Composites of 12 Individuals) | Toxin Concentration (mg/kg) | ||||
|---|---|---|---|---|---|
| PTX-2 | PTX-2 seco Acid a | Okadaic Acid | Okadaic Acid Released by Hydrolysis | Total DSTs | |
| Whole scallop | 0.023 | 0.51 | 0.018 | <0.050 | 0.041 |
| Processed scallop b | <0.010 | 0.037 | 0.014 | <0.050 | 0.014 |
| Whole razorfish | 0.013 | 0.15 | 0.051 | 0.089 | 0.153 |
| Processed razorfish c | <0.010 | 0.023 | <0.010 | <0.050 | <0.050 |
| Whole oyster | 0.11 | 0.79 | 0.023 | 0.12 | 0.253 |
| FSANZ maximum limit for DSP toxins in bivalve molluscs | 0.2 | ||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ajani, P.; Harwood, D.T.; Murray, S.A. Recent Trends in Marine Phycotoxins from Australian Coastal Waters. Mar. Drugs 2017, 15, 33. https://0-doi-org.brum.beds.ac.uk/10.3390/md15020033
Ajani P, Harwood DT, Murray SA. Recent Trends in Marine Phycotoxins from Australian Coastal Waters. Marine Drugs. 2017; 15(2):33. https://0-doi-org.brum.beds.ac.uk/10.3390/md15020033
Chicago/Turabian StyleAjani, Penelope, D. Tim Harwood, and Shauna A. Murray. 2017. "Recent Trends in Marine Phycotoxins from Australian Coastal Waters" Marine Drugs 15, no. 2: 33. https://0-doi-org.brum.beds.ac.uk/10.3390/md15020033