
Synthesis, Structural and Pharmacological Characterizations of CIC, a Novel α-Conotoxin with an Extended N-Terminal Tail

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Results
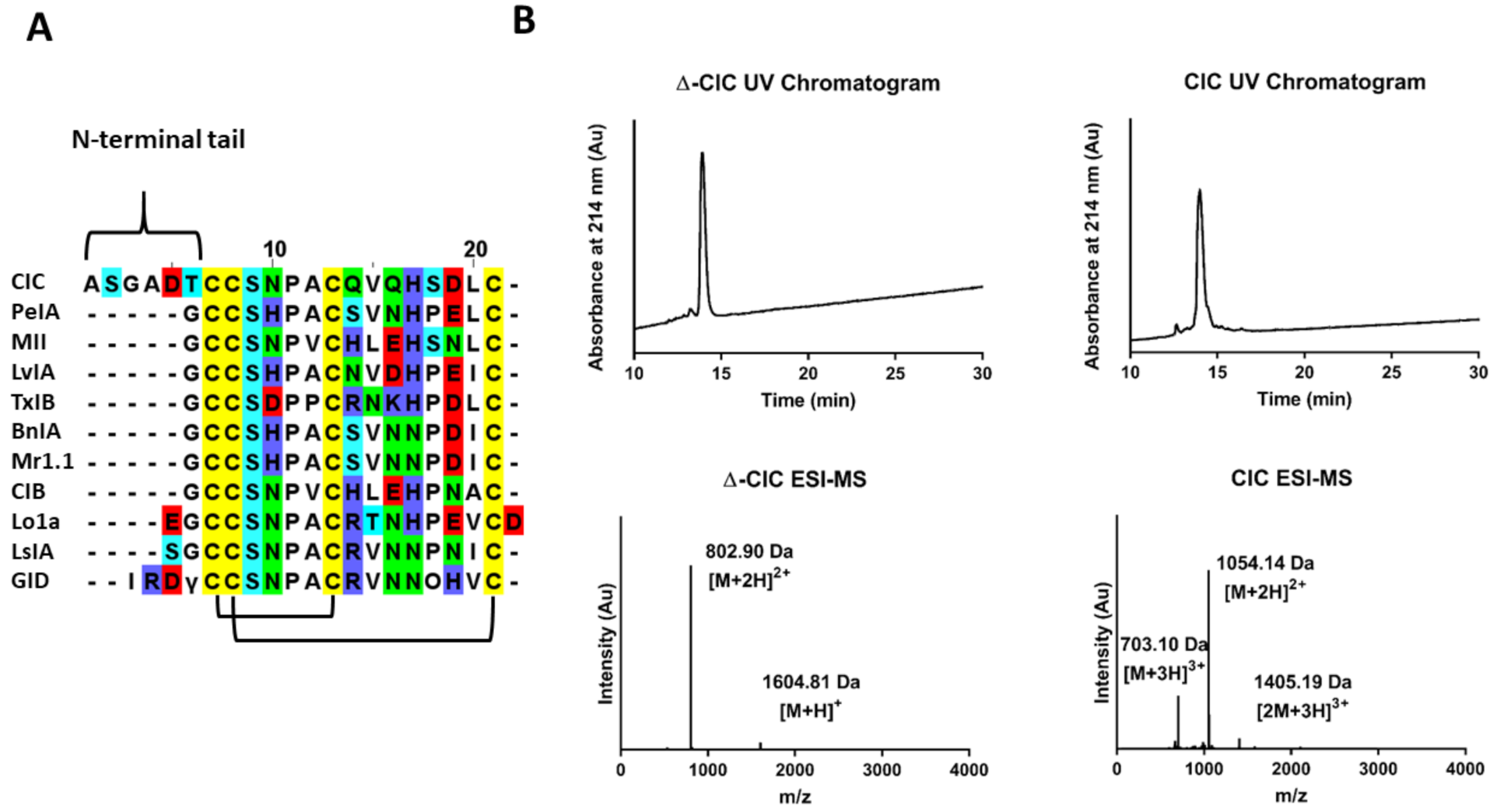
2.1. Chemical Synthesis of CIC and ∆-CIC
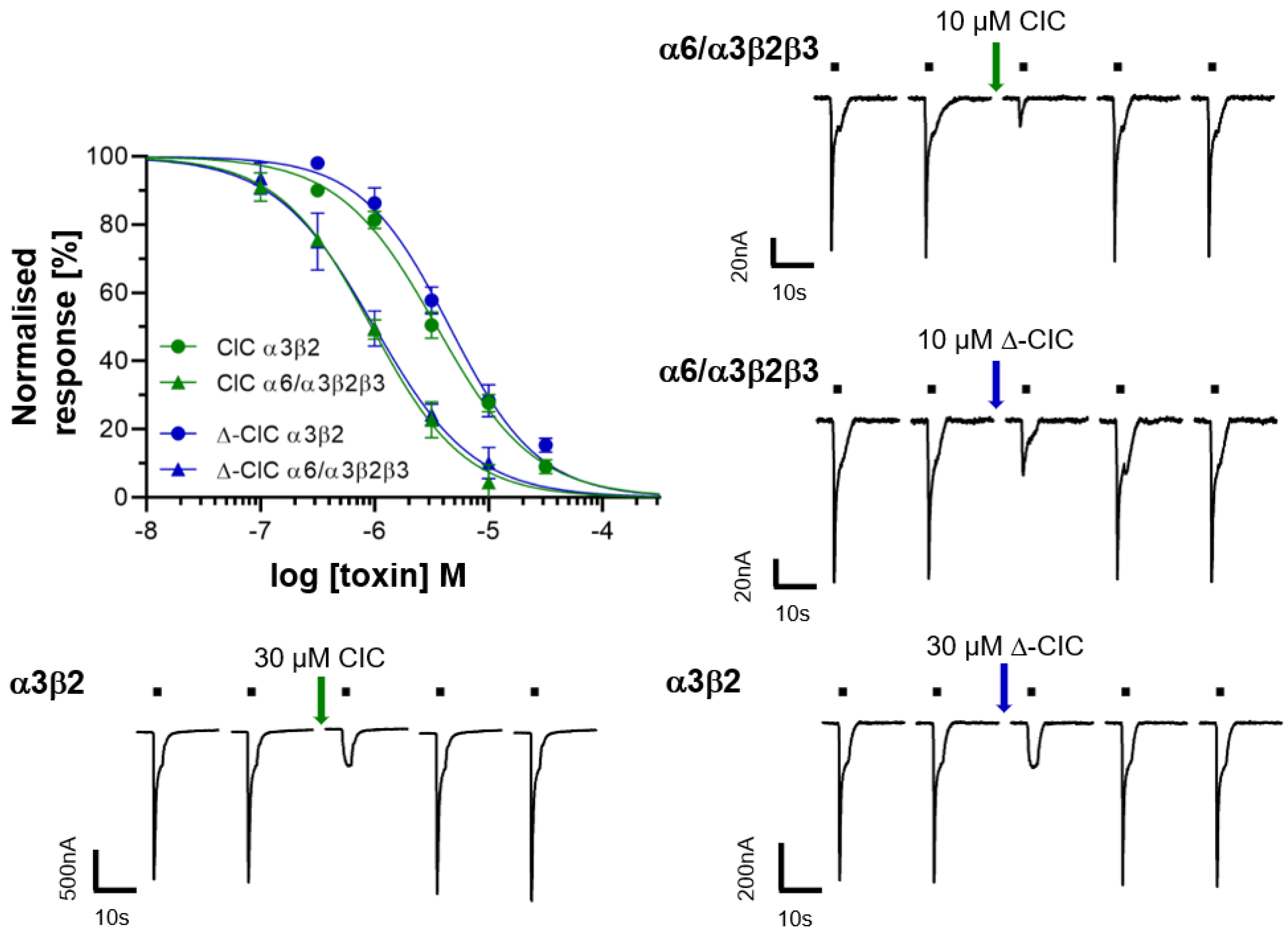
2.2. Electrophysiology of CIC and ∆-CIC on Neuronal nAChRs

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| nAChR Subtypes | Toxin (IC50) | |||
|---|---|---|---|---|
| CIC | Δ-CIC | MII | GID | |
| α3β2 | 3.51 µM | 4.56 µM | 3.7 nM | 3.1 nM |
| α6/α3β2β3 | 1.03 µM | 1.08 µM | 390 pM | N.D |
| α7 | >10 µM | >10 µM | 200 nM (56% inhibition) | 4.5 nM |
| α4β2 | >10 µM | >10 µM | 200 nM (30% inhibition) | 152 nM |
| α4β4 | >10 µM | >10 µM | 200 nM (4% inhibition) | >5 µM |
| α3β4 | >10 µM | >10 µM | 200 nM (15% inhibition) | >5 µM |
| α2β2 | >10 µM | >10 µM | 200 nM (20% inhibition) | N.D |
| α2β4 | >10 µM | >10 µM | 200 nM (4% inhibition) | N.D |
| (α1)2β1δγ | >10 µM | >10 µM | 200 nM (11% inhibition) | N.D |
| α9α10 | >10 µM | >10 µM | >1 µM | N.D |
| References | This work | This work | [14,16,17,18] | [10] |
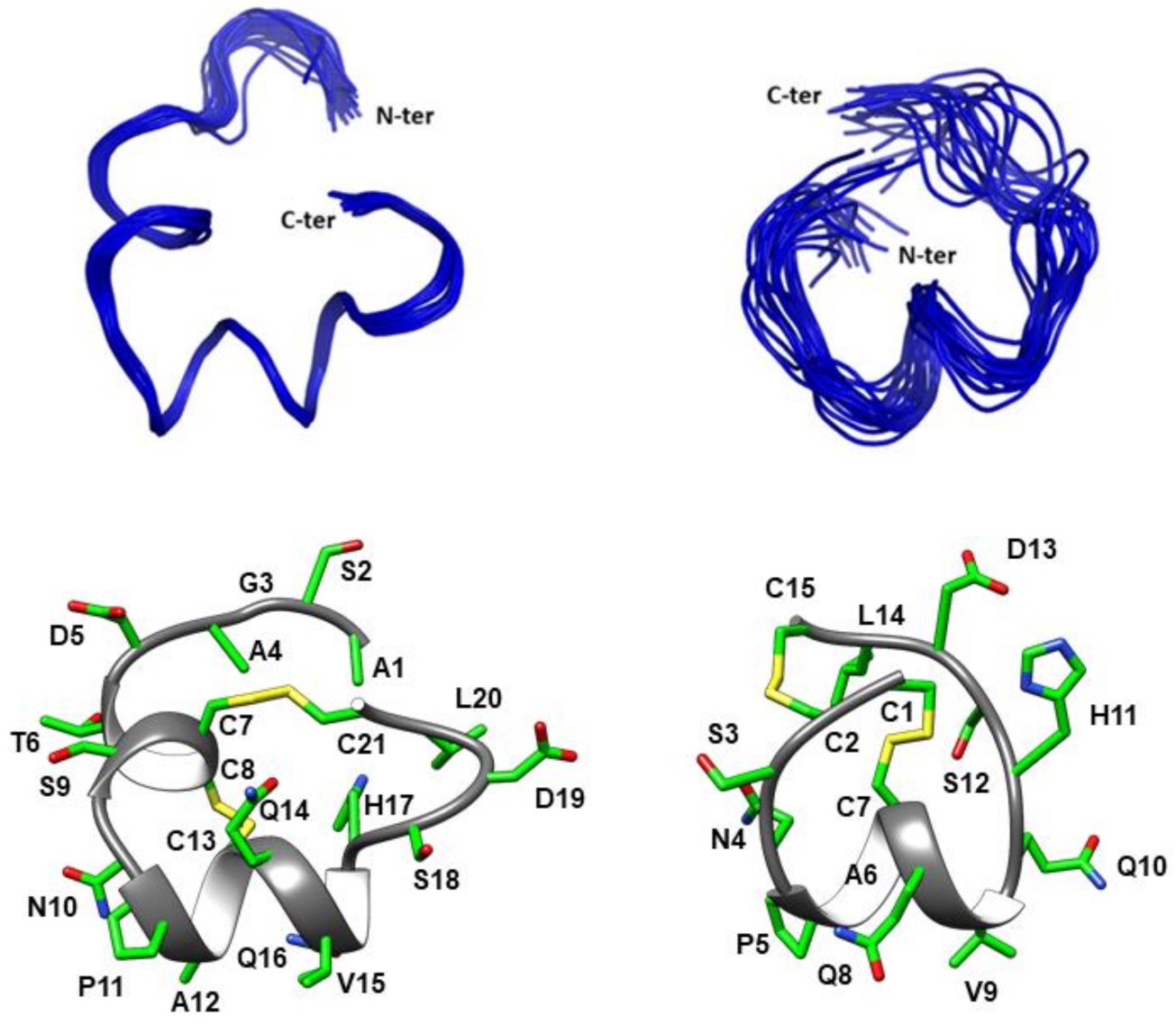
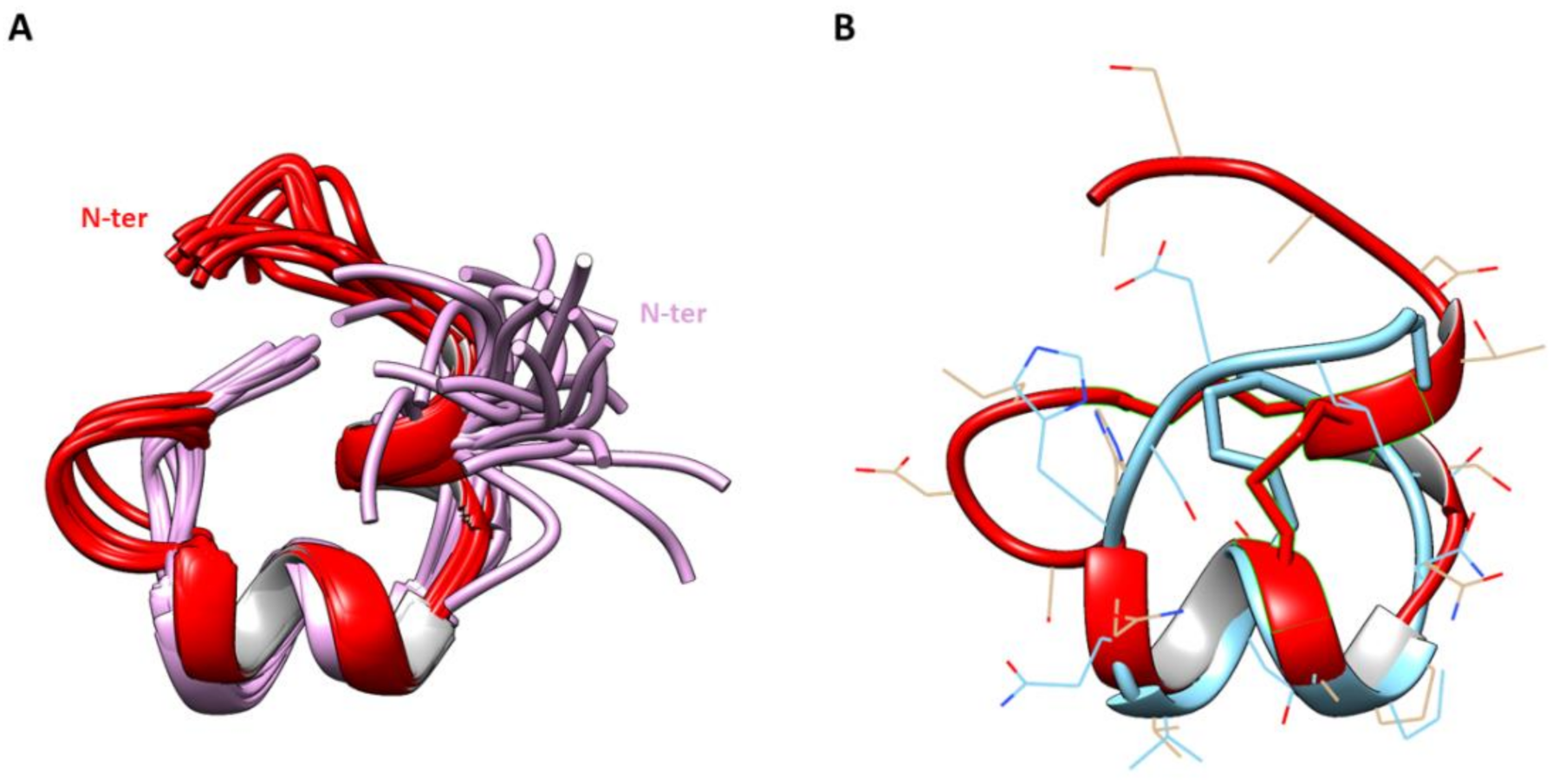
2.3. Three-Dimensional Structure Determination of CIC and ∆-CIC
3. Discussion
4. Materials and Methods
4.1. Abbreviations
4.2. Chemical Synthesis
4.3. Mass Spectrometry
4.3.1. Solvents Used for LC/MS Were of HPLC Grade
4.3.2. Preparative RP-HPLC
4.4. Electrophysiology
4.5. NMR Structure Determination
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giribaldi, J.; Dutertre, S. α-Conotoxins to explore the molecular, physiological and pathophysiological functions of neuronal nicotinic acetylcholine receptors. Neurosci. Lett. 2018, 679, 24–34. [Google Scholar] [CrossRef]
- Puillandre, N.; Duda, T.F.; Meyer, C.P.; Olivera, B.M.; Bouchet, P. One, four or 100 genera? A new classification of the cone snails. J. Molluscan Stud. 2015, 81, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutertre, S.; Jin, A.-H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of separate predation- and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, J.; Jones, A.; Lewis, R.J. Remarkable inter- and intra-species complexity of conotoxins revealed by LC/MS. Peptides 2009, 30, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Akondi, K.B.; Muttenthaler, M.; Dutertre, S.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Discovery, Synthesis, and Structure–Activity Relationships of Conotoxins. Chem. Rev. 2014, 114, 5815–5847. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Garcia, M.L. Therapeutic potential of venom peptides. Nat. Rev. Drug Discov. 2003, 2, 790–802. [Google Scholar] [CrossRef] [PubMed]
- El Hamdaoui, Y.; Wu, X.; Clark, R.J.; Giribaldi, J.; Anangi, R.; Craik, D.J.; King, G.F.; Dutertre, S.; Kaas, Q.; Herzig, V.; et al. Periplasmic Expression of 4/7 α-Conotoxin TxIA Analogs in E. coli Favors Ribbon Isomer Formation—Suggestion of a Binding Mode at the α7 nAChR. Front. Pharmacol. 2019, 10, 577. [Google Scholar] [CrossRef] [Green Version]
- Kaas, Q.; Westermann, J.-C.; Halai, R.; Wang, C.K.L.; Craik, D.J. ConoServer, a database for conopeptide sequences and structures. Bioinformatics 2008, 24, 445–446. [Google Scholar] [CrossRef] [Green Version]
- Kaas, Q.; Yu, R.; Jin, A.-H.; Dutertre, S.; Craik, D.J. ConoServer: Updated content, knowledge, and discovery tools in the conopeptide database. Nucleic Acids Res. 2011, 40, D325–D330. [Google Scholar] [CrossRef]
- Nicke, A.; Loughnan, M.L.; Millard, E.L.; Alewood, P.F.; Adams, D.J.; Daly, N.L.; Craik, D.J.; Lewis, R.J. Isolation, Structure, and Activity of GID, a Novel α4/7-Conotoxin with an Extended N-terminal Sequence. J. Biol. Chem. 2003, 278, 3137–3144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cordero-Erausquin, M.; Marubio, L.M.; Klink, R.; Changeux, J.-P. Nicotinic receptor function: New perspectives from knockout mice. Trends Pharmacol. Sci. 2000, 21, 211–217. [Google Scholar] [CrossRef]
- Himaya, S.W.A.; Jin, A.-H.; Dutertre, S.; Giacomotto, J.; Mohialdeen, H.; Vetter, I.; Alewood, P.F.; Lewis, R.J. Comparative Venomics Reveals the Complex Prey Capture Strategy of the Piscivorous Cone Snail Conus catus. J. Proteome Res. 2015, 14, 4372–4381. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, J.M.; Plazas, P.V.; Watkins, M.; Gomez-Casati, M.E.; Olivera, B.M.; Elgoyhen, A.B. A Novel α-Conotoxin, PeIA, Cloned from Conus pergrandis, Discriminates between Rat α9α10 and α7 Nicotinic Cholinergic Receptors. J. Biol. Chem. 2005, 280, 30107–30112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cartier, G.E.; Yoshikami, D.; Gray, W.R.; Luo, S.; Olivera, B.M.; McIntosh, J.M. A New α-Conotoxin Which Targets α3β2 Nicotinic Acetylcholine Receptors. J. Biol. Chem. 1996, 271, 7522–7528. [Google Scholar] [CrossRef] [Green Version]
- Giribaldi, J.; Wilson, D.; Nicke, A.; El Hamdaoui, Y.; Laconde, G.; Faucherre, A.; Moha, O.; Maati, H.; Daly, N.; Enjalbal, C.; et al. Synthesis, Structure and Biological Activity of CIA and CIB, Two α-Conotoxins from the Predation-Evoked Venom of Conus catus. Toxins 2018, 10, 222. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, J.M.; Azam, L.; Staheli, S.; Dowell, C.; Lindstrom, J.M.; Kuryatov, A.; Garrett, J.E.; Marks, M.J.; Whiteaker, P. Analogs of α-Conotoxin MII Are Selective for α6-Containing Nicotinic Acetylcholine Receptors. Mol. Pharmacol. 2004, 65, 944–952. [Google Scholar] [CrossRef] [PubMed]
- Klimis, H.; Adams, D.J.; Callaghan, B.; Nevin, S.; Alewood, P.F.; Vaughan, C.W.; Mozar, C.A.; Christie, M.J. A novel mechanism of inhibition of high-voltage activated calcium channels by α-conotoxins contributes to relief of nerve injury-induced neuropathic pain. Pain 2011, 152, 259–266. [Google Scholar] [CrossRef]
- Everhart, D.; Cartier, G.E.; Malhotra, A.; Gomes, A.V.; McIntosh, J.M.; Luetje, C.W. Determinants of Potency on α-Conotoxin MII, a Peptide Antagonist of Neuronal Nicotinic Receptors. Biochemistry 2004, 43, 2732–2737. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Nicke, A.; Lewis, R.J. β2 Subunit Contribution to 4/7 α-Conotoxin Binding to the Nicotinic Acetylcholine Receptor. J. Biol. Chem. 2005, 280, 30460–30468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, D.T.; Bansal, P.S.; Carter, D.A.; Vetter, I.; Nicke, A.; Dutertre, S.; Daly, N.L. Characterisation of a Novel A-Superfamily Conotoxin. Biomedicines 2020, 8, 128. [Google Scholar] [CrossRef]
- Kuryatov, A.; Olale, F.; Cooper, J.; Choi, C.; Lindström, J. Human α6 AChR subtypes: Subunit composition, assembly, and pharmacological responses. Neuropharmacology 2000, 39, 2570–2590. [Google Scholar] [CrossRef]
- Skinner, S.P.; Fogh, R.H.; Boucher, W.; Ragan, T.J.; Mureddu, L.G.; Vuister, G.W. CcpNmr AnalysisAssign: A flexible platform for integrated NMR analysis. J. Biomol. NMR 2016, 66, 111–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Güntert, P. Automated NMR Structure Calculation with CYANA. In Protein NMR Techniques; Humana Press: Totowa, NJ, USA, 2004; pp. 353–378. [Google Scholar]
- Shen, Y.; Delaglio, F.; Cornilescu, G.; Bax, A. TALOS+: A hybrid method for predicting protein backbone torsion angles from NMR chemical shifts. J. Biomol. NMR 2009, 44, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Koradi, R.; Billeter, M.; Wüthrich, K. MOLMOL: A program for display and analysis of macromolecular structures. J. Mol. Graph. 1996, 14, 51–55. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera? A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]




| Experimental Restraints | CIC | Δ-CIC |
|---|---|---|
| Interproton distance restraints | 230 | 88 |
| Intraresidue | 64 | 36 |
| Sequential | 75 | 45 |
| Medium range (i − j < 5) | 79 | 7 |
| Long range (i − j ≥5) | 12 | 0 |
| Dihedral-angle restraints | 30 | 15 |
| R.m.s. deviations from the mean coordinate structure (Å) | ||
| Backbone atoms | 0.79 +/− 0.3 | 2.06 +/− 0.73 |
| All heavy atoms | 1.11 +/− 0.22 | 2.97 +/− 0.69 |
| Ramachandran statistics | ||
| % in most favoured region | 82.6% | 62.2% |
| % in additionally allowed region | 17.4% | 33.7% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giribaldi, J.; Haufe, Y.; Evans, E.R.J.; Wilson, D.T.; Daly, N.L.; Enjalbal, C.; Nicke, A.; Dutertre, S. Synthesis, Structural and Pharmacological Characterizations of CIC, a Novel α-Conotoxin with an Extended N-Terminal Tail. Mar. Drugs 2021, 19, 141. https://0-doi-org.brum.beds.ac.uk/10.3390/md19030141
Giribaldi J, Haufe Y, Evans ERJ, Wilson DT, Daly NL, Enjalbal C, Nicke A, Dutertre S. Synthesis, Structural and Pharmacological Characterizations of CIC, a Novel α-Conotoxin with an Extended N-Terminal Tail. Marine Drugs. 2021; 19(3):141. https://0-doi-org.brum.beds.ac.uk/10.3390/md19030141
Chicago/Turabian StyleGiribaldi, Julien, Yves Haufe, Edward R. J. Evans, David T. Wilson, Norelle L. Daly, Christine Enjalbal, Annette Nicke, and Sébastien Dutertre. 2021. "Synthesis, Structural and Pharmacological Characterizations of CIC, a Novel α-Conotoxin with an Extended N-Terminal Tail" Marine Drugs 19, no. 3: 141. https://0-doi-org.brum.beds.ac.uk/10.3390/md19030141