Impact of Gender and Age on Hyperthermia-Induced Changes in Respiration of Liver Mitochondria


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Isolation of Mitochondria
2.3. Determination of the Dissolved Molecular Oxygen Concentration
2.4. Measurement of Mitochondrial Respiration
2.5. Measurement of Pyruvate Dehydrogenase (PDH) and Glutamate Dehydrogenase (GDH) Activity in Liver Mitochondria
2.6. Measurement of Mitochondrial ROS Production
2.7. Statistical Analysis
3. Results
3.1. Gender and Age Dependence of the Respiratory Rates at 37 °C
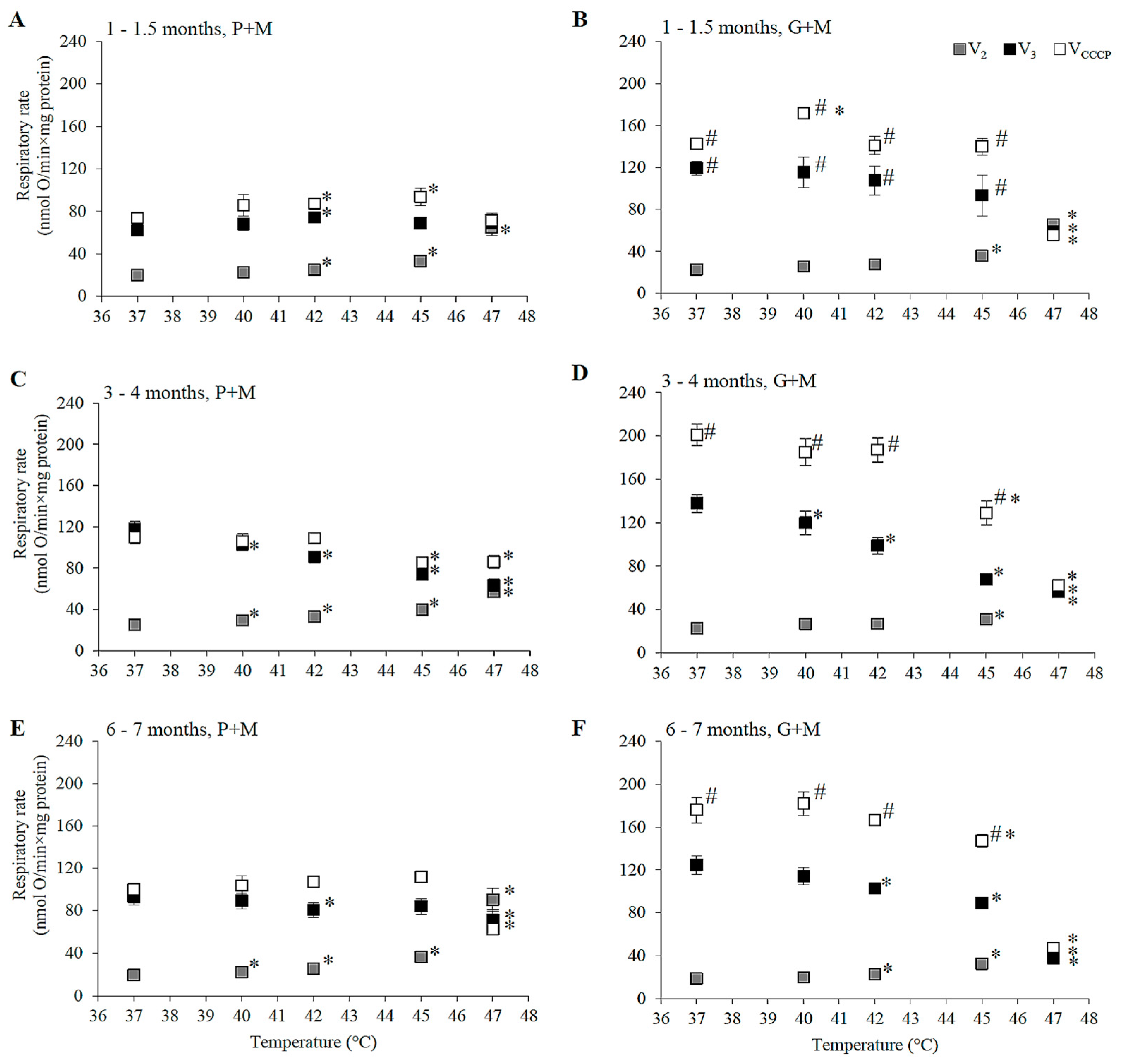
3.2. Effects of HT on Respiration of Mitochondria Oxidizing P+M
3.3. Effects of HT on Respiration in Mitochondria Oxidizing G+M
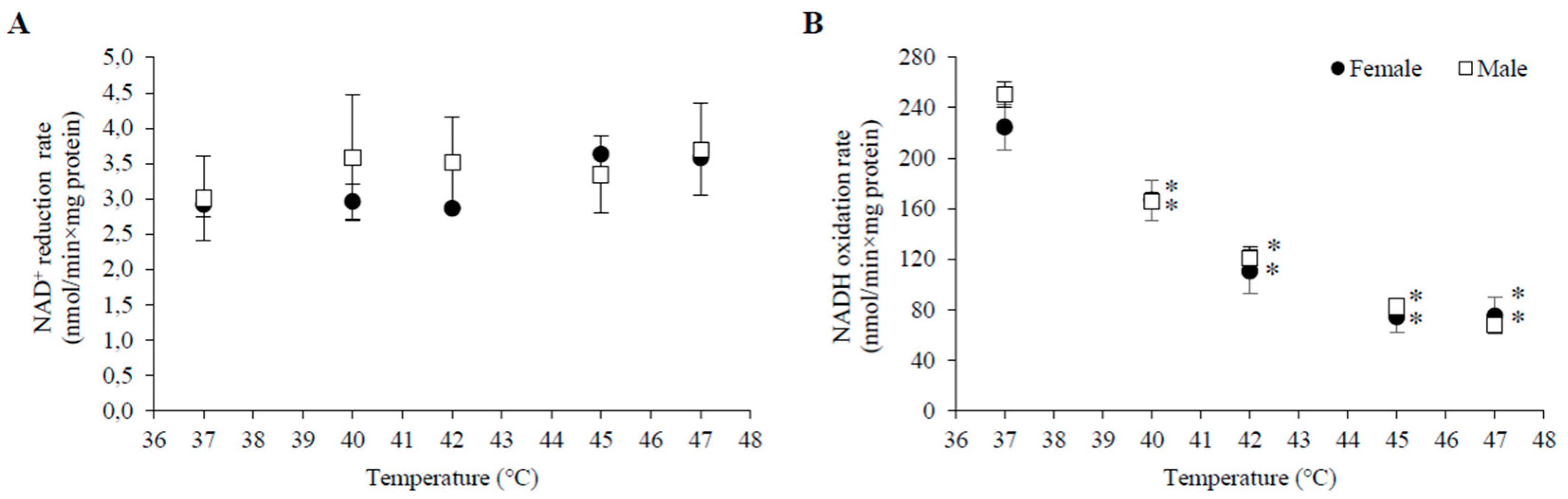
3.4. Effects of HT on Mitochondrial ROS Generation
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Mallory, M.; Gogineni, E.; Jones, G.C.; Greer, L.; Simone, C.B., II. Therapeutic hyperthermia: The old, the new, and the upcoming. Crit. Rev. Oncol. Hematol. 2016, 97, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Guevara, R.; Gianotti, M.; Oliver, J.; Roca, P. Age and sex-related changes in rat brain mitochondrial oxidative status. Exp. Gerontol. 2011, 46, 923–928. [Google Scholar] [CrossRef] [PubMed]
- Colom, B.; Oliver, J.; Garcia-Palmer, F.J. Sexual Dimorphism in the Alterations of Cardiac Muscle Mitochondrial Bioenergetics Associated to the Ageing Process. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 70, 1360–1369. [Google Scholar] [CrossRef] [PubMed]
- Lucas, R.A.; Sarma, S.; Schlader, Z.J.; Pearson, J.; Crandall, C.G. Age-related changes to cardiac systolic and diastolic function during whole-body passive hyperthermia. Exp. Physiol. 2015, 100, 422–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sholomskas, L.M.; Roche, K.L.; Bloomer, S.A. Aging impairs induction of redox factor-1 after heat stress: A potential mechanism for heat-induced liver injury. Int. J. Physiol. Pathophysiol. Pharmacol. 2015, 7, 14–26. [Google Scholar] [PubMed]
- Ratnu, V.S.; Emami, M.R.; Bredy, T.W. Genetic and epigenetic factors underlying sex differences in the regulation of gene expression in the brain. J. Neurosci. Res. 2017, 95, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Valencak, T.G.; Osterrieder, A.; Schulz, T.J. Sex matters: The effects of biological sex on adipose tissue biology and energy metabolism. Redox Biol. 2017, 12, 806–813. [Google Scholar] [CrossRef] [PubMed]
- Demarest, T.G.; McCarthy, M.M. Sex differences in mitochondrial (dys)function: Implications for neuroprotection. J. Bioenerg. Biomembr. 2015, 47, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Clayton, J.A.; Collins, F.S. Policy: NIH to balance sex in cell and animal studies. Nature 2014, 509, 282–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Schadt, E.E.; Wang, S.; Wang, H.; Arnold, A.P.; Ingram-Drake, L.; Drake, T.A.; Lusis, A.J. Tissue-specific expression and regulation of sexually dimorphic genes in mice. Genome Res. 2006, 16, 995–1004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harada, H.; Pavlick, K.P.; Hines, I.N.; Hoffman, J.M.; Bharwani, S.; Gray, L.; Wolf, R.E.; Grisham, M.B. Selected contribution: Effects of gender on reduced-size liver ischemia and reperfusion injury. J. Appl. Physiol. 2001, 91, 2816–2822. [Google Scholar] [CrossRef] [PubMed]
- Jarrar, D.; Wang, P.; Cioffi, W.G.; Bland, K.I.; Chaudry, I.H. The female reproductive cycle is an important variable in the response to trauma-hemorrhage. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, H1015–H1021. [Google Scholar] [CrossRef] [PubMed]
- Colantoni, A.; Idilman, R.; De Maria, N.; La Paglia, N.; Belmonte, J.; Wezeman, F.; Emanuele, N.; Van Thiel, D.H.; Kovacs, E.J.; Emanuele, M.A. Hepatic apoptosis and proliferation in male and female rats fed alcohol: Role of cytokines. Alcohol. Clin. Exp. Res. 2003, 27, 1184–1189. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, K.; Tabuchi, Y.; Kondo, T. Hyperthermia: An effective strategy to induce apoptosis in cancer cells. Apoptosis 2015, 20, 1411–1419. [Google Scholar] [CrossRef] [PubMed]
- Slimen, I.B.; Najar, T.; Ghram, A.; Dabbebi, H.; Ben Mrad, M.; Abdrabbah, M. Reactive oxygen species, heat stress and oxidative-induced mitochondrial damage. A review. Int. J. Hyperth. 2014, 30, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Ventura-Clapier, R.; Moulin, M.; Piquereau, J.; Lemaire, C.; Mericskay, M.; Veksler, V.; Garnier, A. Mitochondria: A central target for sex differences in pathologies. Clin. Sci. 2017, 131, 803–822. [Google Scholar] [CrossRef] [PubMed]
- Lash, L.H.; Qian, W.; Putt, D.A.; Hueni, S.E.; Elfarra, A.A.; Krause, R.J.; Parker, J.C. Renal and hepatic toxicity of trichloroethylene and its glutathione-derived metabolites in rats and mice: Sex-, species-, and tissue-dependent differences. J. Pharmacol. Exp. Ther. 2001, 297, 155–164. [Google Scholar] [PubMed]
- Valle, A.; Guevara, R.; García-Palmer, F.J.; Roca, P.; Oliver, J. Sexual dimorphism in liver mitochondrial oxidative capacity is conserved under caloric restriction conditions. Am. J. Physiol. Cell Physiol. 2007, 293, C1302–C1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catala-Niell, A.; Estrany, M.E.; Proenza, A.M.; Gianotti, M.; Llado, I. Skeletal muscle and liver oxidative metabolism in response to a voluntary isocaloric intake of a high fat diet in male and female rats. Cell Physiol. Biochem. 2008, 22, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Nadal-Casellas, A.; Amengual-Cladera, E.; Proenza, A.M.; Llado, I.; Gianotti, M. Long-term high-fat-diet feeding impairs mitochondrial biogenesis in liver of male and female rats. Cell Physiol. Biochem. 2010, 26, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Chweih, H.; Castilho, R.F.; Figueira, T.R. Tissue and sex specificities in Ca2+ handling by isolated mitochondria in conditions avoiding the permeability transition. Exp. Physiol. 2015, 100, 1073–1092. [Google Scholar] [CrossRef] [PubMed]
- Nauciene, Z.; Zukiene, R.; Degutyte-Fomins, L.; Mildaziene, V. Mitochondrial membrane barrier function as a target of hyperthermia. Medicina 2012, 48, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Chance, B.; Williams, G.R. Respiratory enzymes in oxidative phosphorylation. I. Kinetics of oxygen utilization. J. Biol. Chem. 1955, 217, 383–393. [Google Scholar] [PubMed]
- Stocco, D.M.; Cascarano, J.; Wilson, M.A. Quantitation of mitochondrial DNA, RNA, and protein in starved and starved-refed rat liver. J. Cell Physiol. 1977, 90, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Tummino, P.J.; Gafni, A. A comparative study of succinate-supported respiration and ATP/ADP translocation in liver mitochondria from adult and old rats. Mech. Ageing Dev. 1991, 59, 177–188. [Google Scholar] [CrossRef]
- Sastre, J.; Pallardo, F.V.; Pla, R.; Pellin, A.; Juan, G.; O’Connor, J.E.; Estrela, J.M.; Miquel, J.; Viña, J. Aging of the liver: Age-associated mitochondrial damage in intact hepatocytes. Hepatology 1996, 24, 1199–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robb, G.W.; Amann, R.P.; Killian, G.J. Daily sperm production and epididymal sperm reserves of pubertal and adult rats. J. Reprod. Fertil. 1978, 54, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Goldman, J.M.; Laws, S.C.; Balchak, S.K.; Cooper, R.L.; Kavlock, R.J. Endocrine-disrupting chemicals: Prepubertal exposures and effects on sexual maturation and thyroid activity in the female rat. A focus on the EDSTAC recommendations. Crit. Rev. Toxicol. 2000, 30, 135–196. [Google Scholar] [CrossRef] [PubMed]
- Zukiene, R.; Nauciene, Z.; Silkuniene, G.; Vanagas, T.; Gulbinas, A.; Zimkus, A.; Mildažienė, V. Contribution of mitochondria to injury of hepatocytes and liver tissue by hyperthermia. Medicina 2017, 53, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Gornal, A.G.; Bardawill, C.J.; David, M.M. Determination of serum protein by means of the burette reaction. J. Biol. Chem. 1949, 177, 751–766. [Google Scholar]
- Ellis, G.; Goldberg, D.M. Optimal conditions for the kinetic assay of serum glutamate dehydrogenase activity at 37 degrees C. Clin. Chem. 1972, 18, 523–527. [Google Scholar] [PubMed]
- Hinman, L.M.; Blass, J.P. An NADH-linked spectrophotometric assay for pyruvate dehydrogenase complex in crude tissue homogenates. J. Biol. Chem. 1981, 256, 6583–6586. [Google Scholar] [PubMed]
- Starkov, A.A. Measurement of mitochondrial ROS production. Methods Mol. Biol. 2010, 648, 245–255. [Google Scholar] [PubMed]
- Willis, W.T.; Jackman, M.R.; Bizeau, M.E.; Pagliassotti, M.J.; Hazel, J.R. Hyperthermia impairs liver mitochondrial function in vitro. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, 1240–1246. [Google Scholar] [CrossRef] [PubMed]
- Justo, R.; Boada, J.; Frontera, M.; Oliver, J.; Bermudez, J.; Gianotti, M. Gender dimorphism in rat liver mitochondrial oxidative metabolism and biogenesis. Am. J. Physiol. Cell Physiol. 2005, 289, C372–C378. [Google Scholar] [CrossRef] [PubMed]
- Hafner, R.P.; Brown, G.C.; Brand, M.D. Analysis of the control of respiration rate, phosphorylation rate, proton leak rate and protonmotive force in isolated mitochondria using the ‘top-down’ approach of metabolic control theory. Eur. J. Biochem. 1990, 188, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Mildaziene, V.; Baniene, R.; Nauciene, Z.; Marcinkeviciute, A.; Morkuniene, R.; Borutaite, V.; Kholodenko, B.; Brown, G.C. Ca2+ stimulates both the respiratory and phosphorylation subsystems in rat heart mitochondria. Biochem. J. 1996, 320, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Krishnamoorthy, G.; Hinkle, P.C. Non-ohmic proton conductance of mitochondria and liposomes. Biochemistry 1984, 23, 1640–1645. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Shin, S.; Cho, S.S.; Park, J. Purification and characterization of glutamate dehydrogenase as another isoprotein binding to the membrane of rough endoplasmic reticulum. J. Cell Biochem. 2000, 76, 244–253. [Google Scholar] [CrossRef]
- Ahn, C.S.; Metallo, C.M. Mitochondria as biosynthetic factories for cancer proliferation. Cancer Metab. 2015, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Friday, E.; Oliver, R., III; Turturro, F.; Welbourne, T. Role of Glutamate Dehydrogenase in Cancer Growth and Homeostasis. In Dehydrogenases; Canuto, R.A., Ed.; InTech: London, UK, 2012; ISBN 978-953-307-019-3. [Google Scholar] [Green Version]
- Yang, L.; Moss, T.; Mangala, L.S.; Marini, J.; Zhao, H.; Wahlig, S.; Armaiz-Pena, G.; Jiang, D.; Achreja, A.; Win, J.; et al. Metabolic shifts toward glutamine regulate tumor growth, invasion and bioenergetics in ovarian cancer. Mol. Syst. Biol. 2014, 10, 728. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Li, D.; Alesi, G.; Fan, J.; Kang, H.; Lu, Z.; Boggon, T.J.; Jin, P.; Yi, H.; Wright, E.R.; et al. Glutamate Dehydrogenase 1 Signals through Antioxidant Glutathione Peroxidase 1 to Regulate Redox Homeostasis and Tumor Growth. Cancer Cell 2015, 27, 257–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinelli, J.B.; Yoon, H.; Ringel, A.E.; Jeanfavre, S.; Clish, C.B.; Haigis, M.C. Metabolic recycling of ammonia via glutamate dehydrogenase supports breast cancer biomass. Science 2017, 358, 941–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zukiene, R.; Nauciene, Z.; Ciapaite, J.; Mildaziene, V. Acute temperature resistance threshold in heart mitochondria: Febrile temperature activates function but exceeding it collapses the membrane barrier. Int. J. Hyperth. 2010, 26, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Mármol, F.; Sanchez, J.; Lopez, D.; Martinez, N.; Xaus, C.; Peralta, C.; Roselló-Catafau, J.; Mitjavila, M.T.; Puig-Perellada, P. Role of oxidative stress and adenosine nucleotides in the liver of aging rats. Physiol. Res. 2010, 59, 553–560. [Google Scholar] [PubMed]
- Muñiz, P.; Barchino, M.J.G.; Iradi, A.; Mahiques, E.; Marco, V.; Oliva, M.R.; Sáez, G.T. Age-related changes of liver antioxidant enzymes and 8-hydroxy-2′-deoxyguanosine during fetal-neonate transition and early rat development. IUBMB Life 2000, 49, 497–500. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | Substrate, Metabolic State | Difference of Respiratory Rate (%) as Compared to 37 °C (p < 0.05) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Temperature, °C | |||||||||
| 40 | 42 | 45 | 47 | 40 | 42 | 45 | 47 | ||
| Male | Female | ||||||||
| 1–1.5 months | P+M | ||||||||
| V2 | - | +25 | +65 | +201 | - | +13 | +43 | +162 | |
| V3 | - | +19 | - | - | +17 | +22 | +17 | - | |
| VCCCP | - | +18 | +27 | - | +27 | +31 | +40 | - | |
| G+M | |||||||||
| V2 | - | - | +57 | +187 | - | - | +26 | +100 | |
| V3 | - | - | - | −51 | - | - | −30 | −52 | |
| VCCCP | +21 | - | - | −61 | +11 | - | −12 | −59 | |
| 3–4 months | P+M | ||||||||
| V2 | +16 | +28 | +60 | +132 | +8 | +27 | +77 | +241 | |
| V3 | −14 | −23 | −37 | −46 | +8 | −17 | −21 | ||
| VCCCP | - | - | −20 | −21 | +20 | +25 | +24 | - | |
| G+M | |||||||||
| V2 | - | - | +35 | +143 | - | +25 | +48 | +150 | |
| V3 | -13 | −28 | −50 | −58 | −16 | −22 | −39 | −51 | |
| VCCCP | - | - | −36 | −69 | - | - | −20 | −65 | |
| 6–7 months | P+M | ||||||||
| V2 | +16 | +32 | +89 | +373 | +27 | +31 | +46 | +374 | |
| V3 | - | −13 | - | −24 | +8 | - | - | - | |
| VCCCP | −36 | +13 | +22 | +22 | |||||
| G+M | |||||||||
| V2 | - | +22 | +76 | +132 | - | +35 | +56 | +187 | |
| V3 | - | −18 | −29 | −70 | −13 | −15 | −28 | −47 | |
| VCCCP | - | - | −16 | −70 | - | - | - | −64 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šilkūnienė, G.; Žūkienė, R.; Naučienė, Z.; Degutytė-Fomins, L.; Mildažienė, V. Impact of Gender and Age on Hyperthermia-Induced Changes in Respiration of Liver Mitochondria. Medicina 2018, 54, 62. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina54040062
Šilkūnienė G, Žūkienė R, Naučienė Z, Degutytė-Fomins L, Mildažienė V. Impact of Gender and Age on Hyperthermia-Induced Changes in Respiration of Liver Mitochondria. Medicina. 2018; 54(4):62. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina54040062
Chicago/Turabian StyleŠilkūnienė, Giedrė, Rasa Žūkienė, Zita Naučienė, Laima Degutytė-Fomins, and Vida Mildažienė. 2018. "Impact of Gender and Age on Hyperthermia-Induced Changes in Respiration of Liver Mitochondria" Medicina 54, no. 4: 62. https://0-doi-org.brum.beds.ac.uk/10.3390/medicina54040062