Molecular Differences Based on Erythrocyte Fatty Acid Profile to Personalize Dietary Strategies between Adults and Children with Obesity

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Descriptive Characteristics of the Participants
2.2. Red Blood Cell Membrane Fatty Acids Profile
2.3. Blood Biochemical Parameters
2.4. Food Groups
2.5. Nutrient Intake
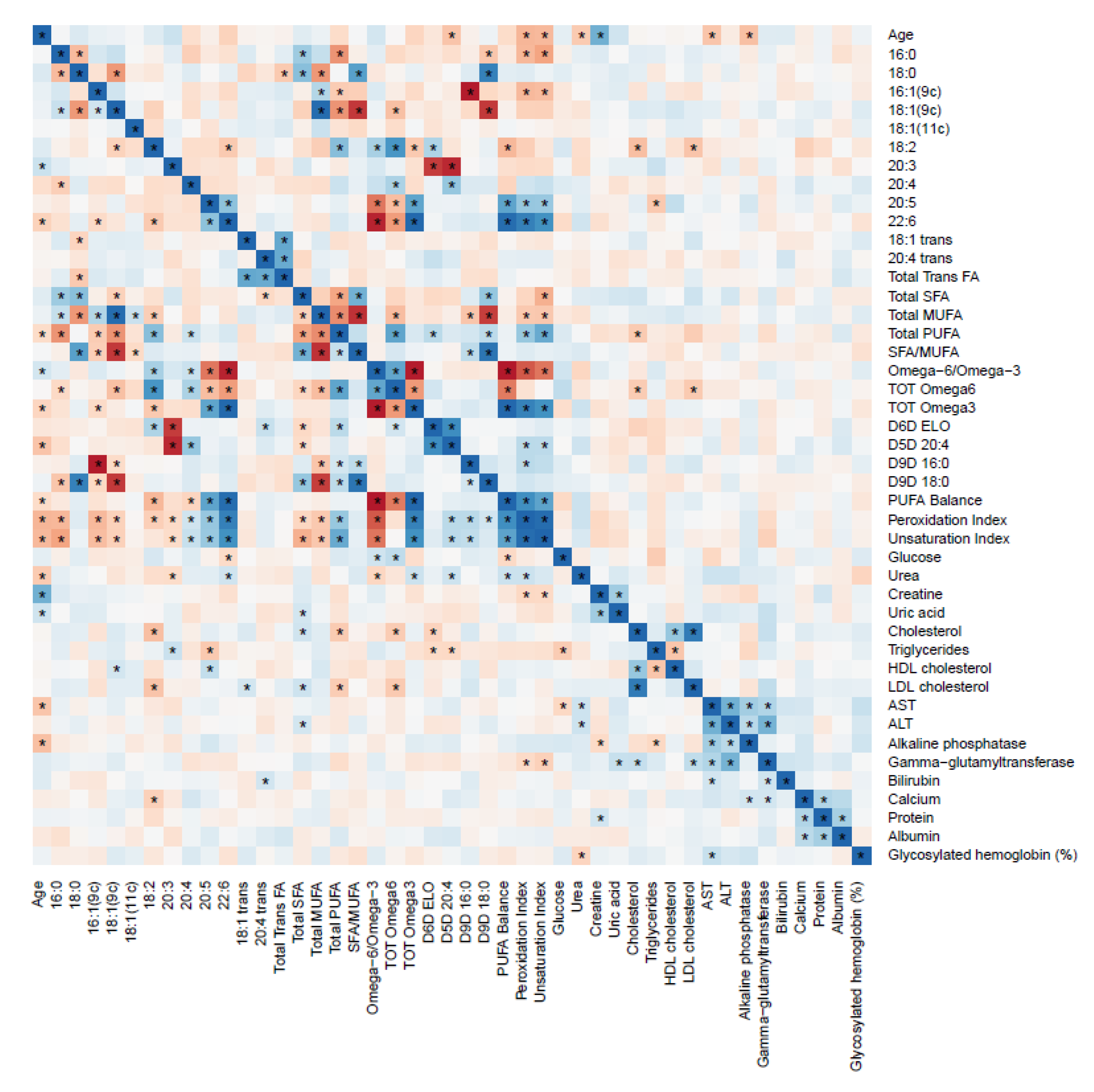
2.6. RBC FAs and Blood Biochemical Parameters Correlation
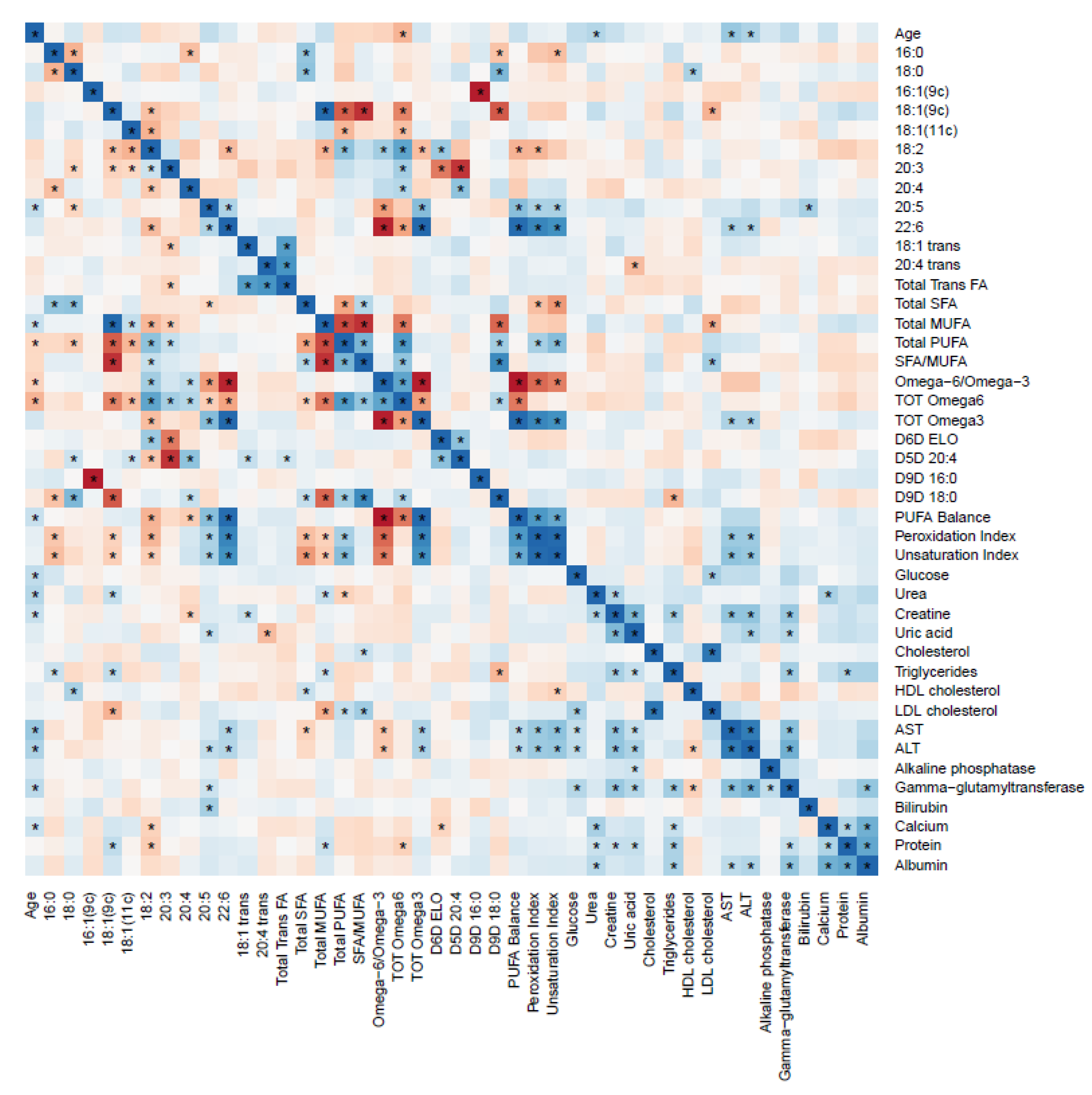
2.7. RBC FAs and Food Groups Values Correlation
3. Discussion
4. Materials and Methods
4.1. Subjects and Study Design
4.2. Anthropometric Measures
4.3. Nutrient Intakes
4.4. Red Blood Cell (RBC) Membrane Fatty Acid Analysis
4.5. Red Blood Cell Membrane Fatty Acid Cluster
4.6. Biochemical Parameters
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | arachidonic acid |
| ANCOVA | analysis of covariance |
| BMI | body mass index |
| DGLA | dihomo–gamma–linolenic acid |
| DHA | docosahexaenoic acid |
| DPA | docosapentaenoic acid |
| EDTA | ethylenediaminetetraacetic acid |
| EFA | essential fatty acid |
| EPA | eicosapentaenoic acid |
| FA | fatty acid |
| FFQ | food frequency questionnaire |
| FAME | fatty acid methyl ester |
| KMO | Kaiser–Meyer–Olkin |
| MeOH | methyl alcohol |
| MUFA | monounsaturated fatty acids |
| KOH | potassium hydroxide |
| LA | linoleic acid |
| PUFA | polyunsaturated fatty acids |
| RBC | red blood cell |
| SFA | saturated fatty acids |
| SCD1 | stearoyl–CoA desaturase–1 |
| SD | standard deviation |
| TFA | trans fatty acids |
| UI | unsaturation index |
| PI | peroxidation index |
| PCA | principal component analysis |
References
- World Health Organization. Obesity and Overweight; World Health Organization: Geneva, Switzerland, 2016; Available online: https://www.who.int/en/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 1 April 2020).
- Tobias, D.K.; Chen, M.; Manson, J.E.; Ludwig, D.S.; Willett, W.; Hu, F.B. Effect of low-fat diet interventions versus other diet interventions on long-term weight change in adults: A systematic review and meta-analysis. Lancet Diabetes Endocrinol. 2015, 3, 968–979. [Google Scholar] [CrossRef] [Green Version]
- Mensink, R.P.; Zock, P.L.; Kester, A.D.; Katan, M.B. Effects of dietary fatty acids and carbohydrates on the ratio of serum total to HDL cholesterol and on serum lipids and apolipoproteins: A meta-analysis of 60 controlled trials. Am. J. Clin. Nutr. 2003, 77, 1146–1155. [Google Scholar] [CrossRef] [PubMed]
- Sacks, F.M.; Katan, M. Randomized clinical trials on the effects of dietary fat and carbohydrate on plasma lipoproteins and cardiovascular disease. Am. J. Med. 2002, 113 (Suppl. 9B), 13S–24S. [Google Scholar] [CrossRef]
- Jakobsen, M.U.; O’Reilly, E.J.; Heitmann, B.L.; Pereira, M.A.; Bälter, K.; Fraser, G.E.; Goldbourt, U.; Hallmans, G.; Knekt, P.; Liu, S.; et al. Major types of dietary fat and risk of coronary heart disease: A pooled analysis of 11 cohort studies. Am. J. Clin. Nutr. 2009, 89, 1425–1432. [Google Scholar]
- Koliaki, C.; Spinos, T.; Spinou, Μ.; Brinia, Μ.-E.; Mitsopoulou, D.; Katsilambros, N. Defining the Optimal Dietary Approach for Safe, Effective and Sustainable Weight Loss in Overweight and Obese Adults. Healthcare 2018, 6, 73. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, P.J.; Carraça, E.V.; Marques, M.M.; Rutter, H.; Oppert, J.M.; De Bourdeaudhuij, I.; Lakerveld, J.; Brug, J. Successful behavior change in obesity interventions in adults: A systematic review of self-regulation mediators. BMC Med. 2015, 13, 84. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Dietetic Products, Nutrition, and Allergies. Scientific Opinion on Dietary Reference Values for fats, including saturated fatty acids, polyunsaturated fatty acids, monounsaturated fatty acids, trans fatty acids, and cholesterol. EFSA J. 2010, 8, 1461. [Google Scholar]
- FAO. Fats and Fatty Acids in Human Nutrition: Report of an Expert Consultation; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010. [Google Scholar]
- Gow, M.L.; Ho, M.; Burrows, T.L.; Baur, L.A.; Stewart, L.; Hutchesson, M.J.; Cowell, C.T.; Collins, C.E.; Garnett, S.P. Impact of dietary macronutrient distribution on BMI and cardiometabolic outcomes in overweight and obese children and adolescents: A systematic review. Nutr. Rev. 2014, 72, 453–470. [Google Scholar] [CrossRef]
- Camilleri, M.; Staiano, A. Insights on Obesity in Children and Adults: Individualizing Management. Trends Endocrinol. Metab. 2019, 30, 724–734. [Google Scholar] [CrossRef]
- Abbott, S.K.; Else, P.L.; Atkins, T.A.; Hulbert, A.J. Fatty acid composition of membrane bilayers: Importance of diet polyunsaturated fat balance. Biochim. Biophys. Acta Biomembr. 2012, 1818, 1309–1317. [Google Scholar] [CrossRef] [Green Version]
- Perona, J.S. Membrane lipid alterations in the metabolic syndrome and the role of dietary oils. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1690–1703. [Google Scholar] [CrossRef] [PubMed]
- Escribá, P.V. Membrane-lipid therapy: A historical perspective of membrane-targeted therapies—From lipid bilayer structure to the pathophysiological regulation of cells. Biochim. Biophys. Acta Biomembr. 2017, 1859, 1493–1506. [Google Scholar] [CrossRef] [PubMed]
- Ferreri, C.; Chatgilialoglu, C. Role of fatty acid-based functional lipidomics in the development of molecular diagnostic tools. Expert Rev. Mol. Diagn. 2012, 12, 767–780. [Google Scholar] [CrossRef] [PubMed]
- Genio, G. Morbid Obesity is Associated to Altered Fatty Acid Profile of Erythrocyte Membranes. J. Diabetes Metab. 2015, 6. [Google Scholar]
- Ghezzo, A.; Visconti, P.; Abruzzo, P.M.; Bolotta, A.; Ferreri, C.; Gobbi, G.; Malisardi, G.; Manfredini, S.; Marini, M.; Nanetti, L.; et al. Oxidative Stress and Erythrocyte Membrane Alterations in Children with Autism: Correlation with Clinical Features. PLoS ONE 2013, 8, e66418. [Google Scholar] [CrossRef] [Green Version]
- Ferreri, C.; Masi, A.; Sansone, A.; Giacometti, G.; Larocca, A.V.; Menounou, G.; Scanferlato, R.; Tortorella, S.; Rota, D.; Conti, M.; et al. Fatty Acids in Membranes as Homeostatic, Metabolic and Nutritional Biomarkers: Recent Advancements in Analytics and Diagnostics. Diagnostics 2016, 7, 1. [Google Scholar] [CrossRef]
- Sánchez González, E.; Carrascosa Lezcano, A.; Fernández García, J.M.; Ferrández Longás, A.; López de Lara, D.; López-Siguero, J.P. Estudios españoles de crecimiento: Situación actual, utilidad y recomendaciones de uso. Anal. Pediatr. 2011, 74, 193.e1–193.e16. [Google Scholar] [CrossRef]
- Lauritzen, L.; Hansen, H.S.; Jørgensen, M.H.; Michaelsen, K.F. The essentiality of long chain n-3 fatty acids in relation to development and function of the brain and retina. Prog. Lipid. Res. 2001, 40, 1–94. [Google Scholar] [CrossRef]
- Khan, F.A.; Fisher, J.G.; Sparks, E.A.; Jaksic, T. Metabolism of Infants and Children. In Pediatric Surgery: General Principles and Newborn Surgery; Puri, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 231–243. [Google Scholar]
- Amezaga, J.; Arranz, S.; Urruticoechea, A.; Ugartemendia, G.; Larraioz, A.; Louka, M.; Uriarte, M.; Ferreri, C.; Tueros, I. Altered Red Blood Cell Membrane Fatty Acid Profile in Cancer Patients. Nutrients 2018, 10, 1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harayama, T.; Riezman, H. Understanding the diversity of membrane lipid composition. Nat. Rev. Mol. Cell. Biol. 2018, 19, 281–296. [Google Scholar] [CrossRef]
- Weiser, M.J.; Butt, C.M.; Mohajeri, M.H. Docosahexaenoic Acid and Cognition throughout the Lifespan. Nutrients 2016, 8, 99. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirtland, S.J. Prostaglandin E1: A review. Prostaglandins Leukot. Essent. Fatty Acids 1988, 32, 165–174. [Google Scholar] [CrossRef]
- Jauregibeitia, I.; Portune, K.; Amezaga, J.; Tueros, I.; Arranz, S. A Journey through ?-3 Supplements: Future Perspectives for Precision Nutrition. J. Food Nutr. Res. 2020, 8, 556–560. [Google Scholar] [CrossRef]
- Alves-Bezerra, M.; Cohen, D.E. Triglyceride Metabolism in the Liver. Compr. Physiol. 2017, 8, 1–8. [Google Scholar] [PubMed]
- Ferreri, C.; Chatgilialoglu, C. Membrane Lipidomics for Personalized Health; John Wiley & Sons: Chichester, UK, 2015. [Google Scholar]
- Sanders, F.W.; Griffin, J.L. De novo lipogenesis in the liver in health and disease: More than just a shunting yard for glucose. Biol. Rev. Camb. Philos. Soc. 2016, 91, 452–468. [Google Scholar] [CrossRef] [Green Version]
- Silbernagel, G.; Kovarova, M.; Cegan, A.; Machann, J.; Schick, F.; Lehmann, R.; Häring, H.U.; Stefan, N.; Schleicher, E.; Fritsche, A.; et al. High Hepatic SCD1 Activity Is Associated with Low Liver Fat Content in Healthy Subjects under a Lipogenic Diet. J. Clin. Endocrinol. Metab. 2012, 97, E2288–E2292. [Google Scholar] [CrossRef]
- Albracht-Schulte, K.; Kalupahana, N.S.; Ramalingam, L.; Wang, S.; Rahman, S.M.; Robert-McComb, J.; Moustaid-Moussa, N. Omega-3 fatty acids in obesity and metabolic syndrome: A mechanistic update. J. Nutr. Biochem. 2018, 58, 1–16. [Google Scholar] [CrossRef]
- Burns, J.L.; Nakamura, M.T.; Ma, D.W.L. Differentiating the biological effects of linoleic acid from arachidonic acid in health and disease. Prostaglandins Leukot. Essent. Fatty Acids 2018, 135, 1–4. [Google Scholar] [CrossRef]
- Ruiz-Núñez, B.; Kuipers, R.S.; Luxwolda, M.F.; De Graaf, D.J.; Breeuwsma, B.B.; Dijck-Brouwer, D.A.; Muskiet, F.A.J. Saturated fatty acid (SFA) status and SFA intake exhibit different relations with serum total cholesterol and lipoprotein cholesterol: A mechanistic explanation centered around lifestyle-induced low-grade inflammation. J. Nutr. Biochem. 2014, 25, 304–312. [Google Scholar] [CrossRef]
- Chen, L.H.; Wang, Y.F.; Xu, Q.H.; Chen, S.S. Omega-3 fatty acids as a treatment for non-alcoholic fatty liver disease in children: A systematic review and meta-analysis of randomized controlled trials. Clin. Nutr. 2018, 37, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Carrascosa, A.; Fernández, J.M.; Fernández, C.; Ferrández, A.; López-Siguero, J.P.; Sánchez, E.; Sobradillo, B.; Yeste, D. Estudios españoles de crecimiento 2008. Nuevos patrones antropométricos. Endocrinol. Nutr. 2008, 55, 484–506. [Google Scholar] [CrossRef]
- Madrigal, C.; Soto-Mendez, M.J.; Hernandez-Ruiz, A.; Ruiz, E.; Valero, T.; Avila, J.M.; Lara-Villoslada, F.; Leis, R.; Victoria, E.M.; Moreno, J.M.; et al. Dietary and Lifestyle Patterns in the Spanish Pediatric Population (One to < 10 Years Old): Design, Protocol, and Methodology of the EsNuPI Study. Nutrients 2019, 11, 3050. [Google Scholar]
- Fernandez-Ballart, J.D.; Pinol, J.L.; Zazpe, I.; Corella, D.; Carrasco, P.; Toledo, E.; Perez-Bauer, M.; Martínez-González, M.A.; Salas-Salvadó, J.; Martín-Moreno, J.M. Relative validity of a semi-quantitative food-frequency questionnaire in an elderly Mediterranean population of Spain. Br. J. Nutr. 2010, 103, 1808–1816. [Google Scholar] [CrossRef] [Green Version]
- Sansone, A.; Tolika, E.; Louka, M.; Sunda, V.; Deplano, S.; Melchiorre, M.; Anagnostopoulos, D.; Chatgilialoglu, C.; Formisano, C.; Di Micco, R.; et al. Hexadecenoic Fatty Acid Isomers in Human Blood Lipids and Their Relevance for the Interpretation of Lipidomic Profiles. PLoS ONE 2016, 11, e0152378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacometti, G.; Ferreri, C.; Sansone, A.; Chatgilialoglu, C.; Marzetti, C.; Spyratou, E.; Georgakilas, A.G.; Marini, M.; Abruzzo, P.M.; Bolotta, A.; et al. High predictive values of RBC membrane-based diagnostics by biophotonics in an integrated approach for Autism Spectrum Disorders. Sci. Rep. 2017, 7, 9854. [Google Scholar] [PubMed]
- Van Der Vegt, S.G.L.; Ruben, A.M.T.; Were, J.M.; Palsma, D.M.H.; Verhoef, C.W.; de Gier, J.; Staal, G.E. Counterflow centrifugation of red cell populations: A cell age related separation technique. Br. J. Haematol. 1985, 61, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Breil, C.; Abert Vian, M.; Zemb, T.; Kunz, W.; Chemat, F. "Bligh and Dyer" and Folch Methods for Solid-Liquid-Liquid Extraction of Lipids from Microorganisms. Comprehension of Solvatation Mechanisms and towards Substitution with Alternative Solvents. Int. J. Mol. Sci. 2017, 18, 708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreri, C.; Faraone Mennella, M.R.; Formisano, C.; Landi, L.; Chatgilialoglu, C. Arachidonate geometrical isomers generated by thiyl radicals: The relationship with trans lipids detected in biological samples. Free Radic. Biol. Med. 2002, 33, 1516–1526. [Google Scholar] [CrossRef]
- Jacobs, S.; Jäger, S.; Jansen, E.; Peter, A.; Stefan, N.; Boeing, H.; Schulze, M.B.; Kröger, J. Associations of Erythrocyte Fatty Acids in the De Novo Lipogenesis Pathway with Proxies of Liver Fat Accumulation in the EPIC-Potsdam Study. PLoS ONE 2015, 10, e0127368. [Google Scholar] [CrossRef]
- Jauregibeitia, I.P.K.; Rica, I.; Tueros, I.; Velasco, O.; Grau, G.; Trebolazabala, N.; Castaño, L.; Larocca, A.V.; Ferreri, C.; Arranz, S. Fatty Acid Profile of Mature Red Blood Cell Membranes and Dietary Intake as a New Approach to Characterize Children with Overweight and Obesity. Nutrients 2020, 12, 3446. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Group with Obesity | Group with Normal Weight | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pediatric n = 83 | Adult n = 61 | Ancova | Pediatric n = 113 | Adult n = 30 | Ancova | |||||
| Fatty Acid (%) | Mean | SE | Mean | SE | p * | Mean | SE | Mean | SE | p * |
| Palmitic acid (C16:0) | 22.31 | 0.16 | 23.22 | 0.22 | 0.01 | 22.51 | 0.10 | 22.72 | 0.23 | 0.43 |
| Stearic acid (C18:0) | 18.22 | 0.18 | 17.54 | 0.24 | 0.06 | 17.68 | 0.10 | 17.66 | 0.23 | 0.94 |
| TOTAL SFA | 40.55 | 0.16 | 40.78 | 0.21 | 0.48 | 40.21 | 0.10 | 40.22 | 0.23 | 0.97 |
| Palmitoleic acid (C16:1) | 0.46 | 0.02 | 0.37 | 0.03 | 0.08 | 0.41 | 0.02 | 0.48 | 0.03 | 0.08 |
| Oleic acid (9c C18:1) | 16.55 | 0.20 | 17.08 | 0.27 | 0.20 | 17.46 | 0.12 | 17.79 | 0.27 | 0.29 |
| cis–Vaccenic acid (11c C18:1) | 1.17 | 0.04 | 1.34 | 0.06 | 0.05 | 1.22 | 0.02 | 1.32 | 0.05 | 0.07 |
| TOTAL MUFA | 18.19 | 0.21 | 18.78 | 0.29 | 0.18 | 19.10 | 0.13 | 19.57 | 0.28 | 0.17 |
| Linoleic acid (C18:2) | 14.00 | 0.27 | 12.39 | 0.37 | 0.01 | 14.22 | 0.12 | 13.12 | 0.27 | <0.01 |
| DGLA (C20:3) | 2.35 | 0.07 | 2.07 | 0.09 | 0.04 | 2.05 | 0.04 | 1.81 | 0.08 | 0.02 |
| ARA (C20:4) | 19.73 | 0.21 | 19.39 | 0.29 | 0.44 | 18.75 | 0.14 | 18.32 | 0.32 | 0.26 |
| TOTAL ω6 | 36.12 | 0.29 | 33.84 | 0.39 | <0.01 | 35.02 | 0.16 | 33.27 | 0.35 | <0.01 |
| EPA (C20:5) | 0.49 | 0.04 | 0.63 | 0.06 | 0.10 | 0.59 | 0.02 | 0.65 | 0.05 | 0.31 |
| DHA (C22:6) | 4.52 | 0.19 | 5.84 | 0.26 | <0.01 | 4.93 | 0.10 | 5.83 | 0.23 | <0.01 |
| TOTAL ω3 | 5.01 | 0.21 | 6.47 | 0.29 | <0.01 | 5.52 | 0.11 | 6.48 | 0.26 | <0.01 |
| TOTAL PUFA | 41.12 | 0.24 | 40.31 | 0.32 | 0.10 | 40.54 | 0.15 | 39.88 | 0.33 | 0.09 |
| Trans C18:1 | 0.08 | 0.01 | 0.07 | 0.01 | 0.65 | 0.08 | 0.01 | 0.10 | 0.02 | 0.34 |
| Trans C20:4 | 0.07 | 0.01 | 0.08 | 0.01 | 0.59 | 0.07 | 0.01 | 0.10 | 0.02 | 0.14 |
| TOTAL TRANS | 0.15 | 0.01 | 0.15 | 0.02 | 0.98 | 0.16 | 0.01 | 0.14 | 0.02 | 0.46 |
| Indexes | ||||||||||
| ω6/ω3 | 7.55 | 0.27 | 5.51 | 0.37 | <0.01 | 6.66 | 0.15 | 5.11 | 0.34 | <0.01 |
| SFA/MUFA | 2.24 | 0.03 | 2.19 | 0.04 | 0.37 | 2.12 | 0.02 | 2.07 | 0.04 | 0.32 |
| Omega–3 Index | 5.01 | 0.21 | 6.47 | 0.29 | <0.01 | 5.52 | 0.11 | 6.48 | 0.26 | <0.01 |
| ∆6D+ELO 20:3/18:2 a | 0.17 | 0.006 | 0.17 | 0.008 | 0.96 | 0.14 | 0.003 | 0.14 | 0.006 | – |
| ∆5D 20:4/20:3 | 8.60 | 0.31 | 9.51 | 0.41 | 0.15 | 9.45 | 0.21 | 10.55 | 0.47 | 0.05 |
| ∆9D 16:1/16:0 | 0.02 | 0.001 | 0.016 | 0.001 | 0.06 | 0.018 | 0.001 | 0.02 | 0.001 | – |
| ∆9D 18:1/18:0 | 0.91 | 0.02 | 0.98 | 0.02 | 0.07 | 0.99 | 0.01 | 1.00 | 0.01 | 0.67 |
| DNL Index 16:0/18:2 | 1.62 | 0.03 | 1.85 | 0.05 | <0.01 | 1.59 | 0.02 | 1.72 | 0.03 | <0.01 |
| PUFA BALANCE | 12.13 | 0.52 | 16.09 | 0.70 | <0.01 | 13.61 | 0.27 | 16.21 | 0.61 | <0.01 |
| Peroxidation Index | 136.77 | 1.42 | 145.07 | 1.91 | 0.01 | 136.81 | 0.82 | 141.31 | 1.89 | 0.04 |
| Unsaturation index | 161.53 | 0.92 | 165.51 | 1.24 | 0.04 | 161.25 | 0.61 | 163.57 | 1.37 | 0.15 |
| Group with Obesity | Group with Normal Weight | |||||
|---|---|---|---|---|---|---|
| Pediatric n = 69 | Adult n = 44 | p * | Pediatric n = 34 | Adult n = 30 | p * | |
| Med (Q1–Q3) | Med (Q1–Q3) | Med (Q1–Q3) | Med (Q1–Q3) | |||
| Glucose (mg/dL) | 85 (79–89.25) | 97 (90.5–107.5) | <0.01 | 84 (81–89) | 85 (79.75–92) | 0.38 |
| Uric Acid (mg/dL) | 4.95 (4.375–5.7) | 5.6 (4.9–6.95) | <0.01 | 3.95 (3.37–4.62) | 4.75 (3.8–5.22) | 0.03 |
| Total Cholesterol (mg/dL) | 150 (132.7–172) | 180 (158–211) | <0.01 | 165 (148.5–186.7) | 176.5 (141.7–206.2) | 0.46 |
| Triglycerides (mg/dL) | 76 (55.5–108.7) | 123 (89.5–180.5) | <0.01 | 65.5 (46–86) | 68 (58.75–84.75) | 0.48 |
| HDL cholesterol (mg/dL) | 44.6 (40.0–54.25) | 47 (41.75–56) | 0.32 | 55 (48.5–64.5) | 59 (48.5–71) | 0.48 |
| LDL cholesterol (mg/dL) | 88.4 (71.25–98) | 118 (95–141) | <0.01 | 95 (77–110) | 102 (72.5–121) | 0.35 |
| AST/GOT (U/L) | 22 (19–26.25) | 20 (16.5–26.5) | 0.12 | 26 (22–27) | 19 (16.75–23.5) | <0.01 |
| ALT/GPT (U/L) | 18.5 (15–23.25) | 23 (15–35.5) | 0.03 | 16 (13.75–18) | 17 (12.75–21.25) | 0.59 |
| Bilirubin (mg/dL) | 0.4 (0.3–0.6) | 0.4 (0.2–0.5) | 0.5 | 0.6 (0.4–1) | 0.4 (0–0.625) | 0.01 |
| Pediatric Group with Obesity, n = 83 | Adult Group with Obesity, n = 61 | Mann-Whitney U Test | |||
|---|---|---|---|---|---|
| Mean | SD | Mean | SD | p | |
| Macronutrient | |||||
| Calories (Kcal/day) | 2044.1 | 564.3 | 2480.1 | 794.2 | <0.01 |
| Proteins (%E) | 16.5 | 2.1 | 16.3 | 3.2 | 0.54 |
| Carbohydrates (%E) | 46.7 | 5.3 | 36.3 | 6.6 | <0.01 * |
| Simple sugars (%E) | 21.7 | 4.9 | 19.1 | 6.4 | <0.01 |
| Lipids (%E) | 33.6 | 6.3 | 42.6 | 6.3 | <0.01 * |
| Individual Fatty Acids | |||||
| C14:0 | 1.0 | 0.5 | 0.8 | 0.3 | 0.08 |
| C16:0 | 6.1 | 1.3 | 5.6 | 1.0 | 0.02 * |
| C18:0 | 2.3 | 0.6 | 2.3 | 0.5 | 0.93 |
| Total SFA | 10.8 | 2.9 | 11.2 | 2.2 | 0.09 |
| C16:1 | 0.5 | 0.1 | 0.5 | 0.1 | 0.86 |
| C18:1 | 14.1 | 3.5 | 18.9 | 4.1 | <0.01 * |
| Total MUFA | 15.0 | 3.6 | 19.9 | 4.2 | <0.01 * |
| C18:2 | 4.1 | 1.7 | 6.9 | 2.5 | <0.01 |
| C20:4 | 0.04 | 0.01 | 0.08 | 0.04 | <0.01 |
| Total ω6 | 4.1 | 1.7 | 7.0 | 2.5 | <0.01 |
| C18:3 | 0.54 | 0.13 | 0.79 | 0.34 | <0.01 |
| C20:5 (EPA) | 0.07 | 0.06 | 0.07 | 0.05 | 0.24 |
| C22:5 (DPA) | 0.02 | 0.01 | 0.02 | 0.01 | 0.48 |
| 22:6 (DHA) | 0.15 | 0.09 | 0.14 | 0.08 | 0.62 |
| Total ω3 | 0.79 | 0.22 | 1.03 | 0.39 | <0.01 |
| Total PUFA | 5.1 | 1.8 | 8.2 | 2.7 | <0.01 |
| ω6/ω3 | 5.5 | 2.2 | 7.2 | 2.8 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jauregibeitia, I.; Portune, K.; Gaztambide, S.; Rica, I.; Tueros, I.; Velasco, O.; Grau, G.; Martín, A.; Castaño, L.; Larocca, A.V.; et al. Molecular Differences Based on Erythrocyte Fatty Acid Profile to Personalize Dietary Strategies between Adults and Children with Obesity. Metabolites 2021, 11, 43. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11010043
Jauregibeitia I, Portune K, Gaztambide S, Rica I, Tueros I, Velasco O, Grau G, Martín A, Castaño L, Larocca AV, et al. Molecular Differences Based on Erythrocyte Fatty Acid Profile to Personalize Dietary Strategies between Adults and Children with Obesity. Metabolites. 2021; 11(1):43. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11010043
Chicago/Turabian StyleJauregibeitia, Iker, Kevin Portune, Sonia Gaztambide, Itxaso Rica, Itziar Tueros, Olaia Velasco, Gema Grau, Alicia Martín, Luis Castaño, Anna Vita Larocca, and et al. 2021. "Molecular Differences Based on Erythrocyte Fatty Acid Profile to Personalize Dietary Strategies between Adults and Children with Obesity" Metabolites 11, no. 1: 43. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11010043