
Hepatocyte Small Heterodimer Partner Mediates Sex-Specific Effects on Triglyceride Metabolism via Androgen Receptor in Male Mice

 ,
,
Abstract
:1. Introduction
2. Results
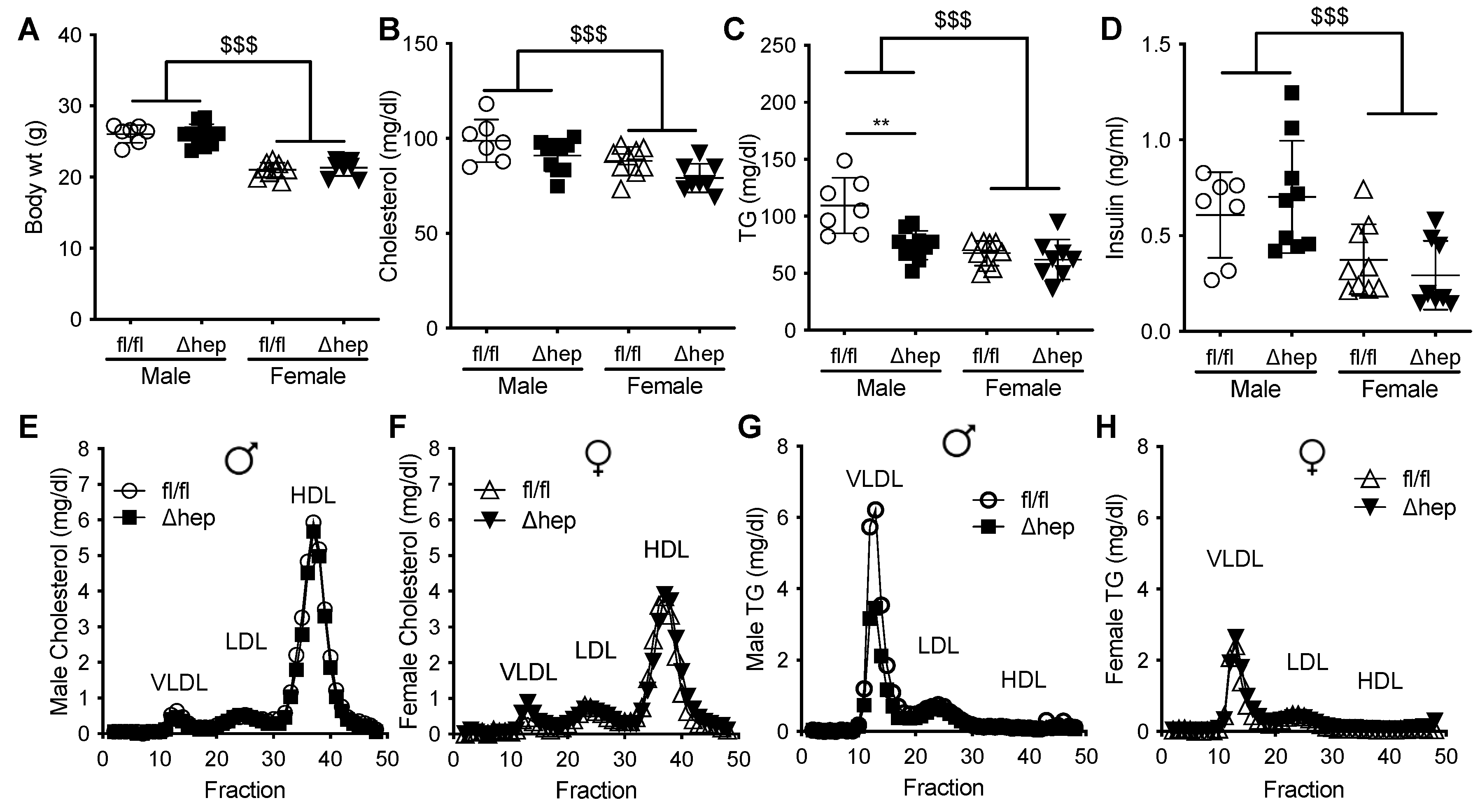
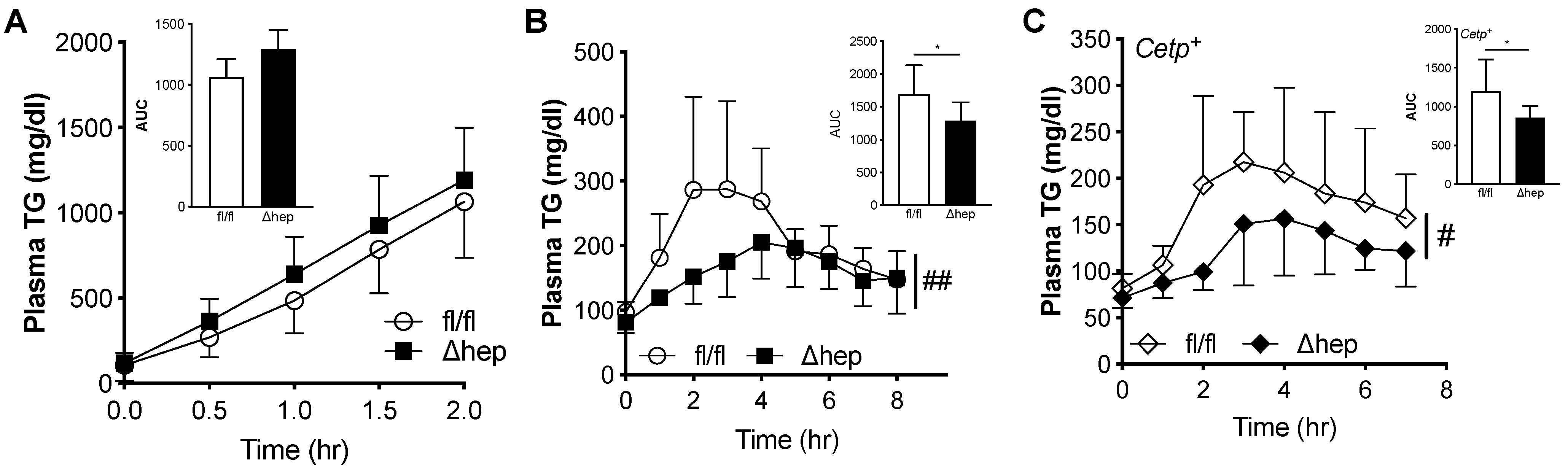
2.1. Effect of Hepatic SHP Deletion on Plasma Lipids
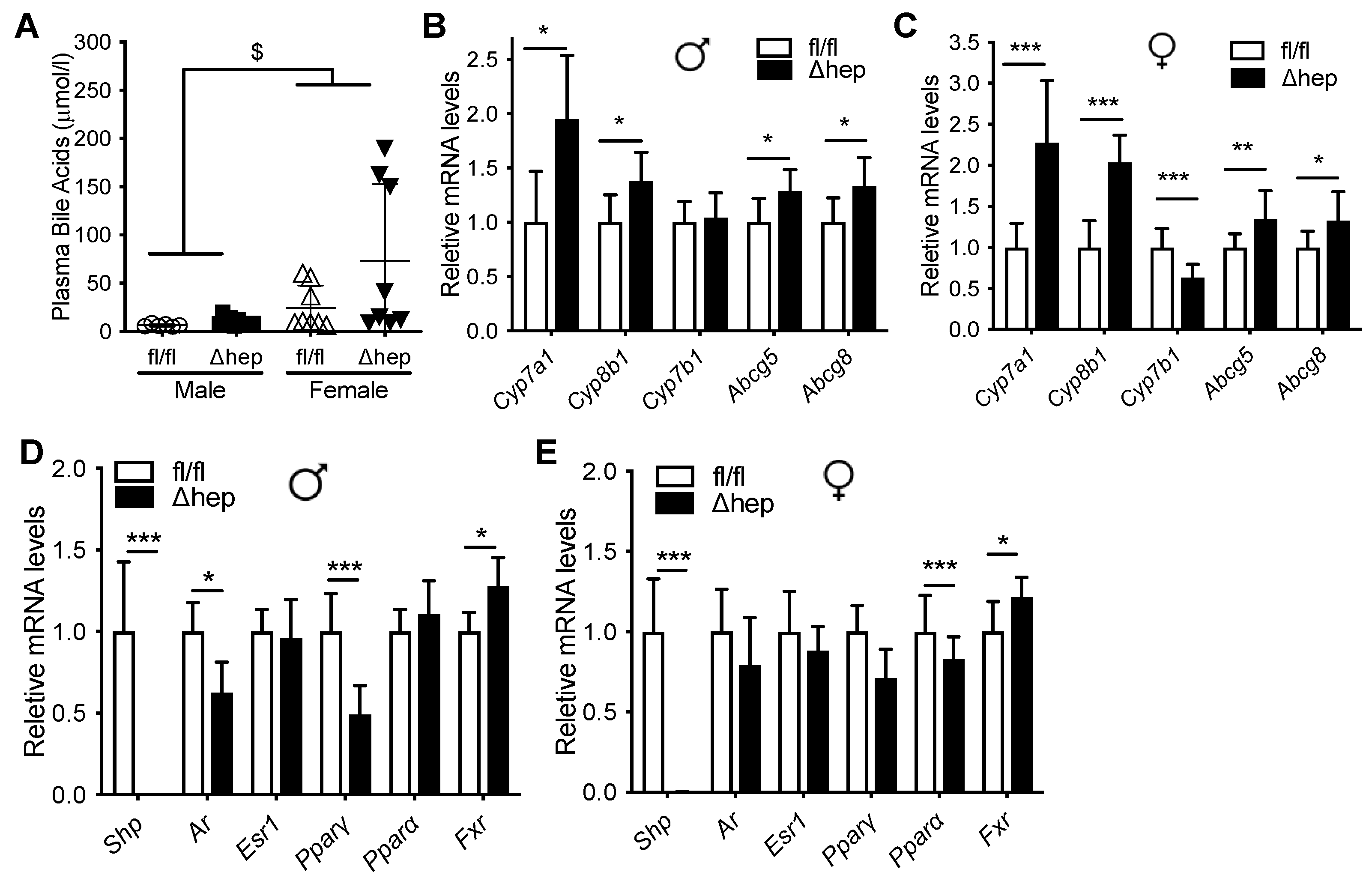
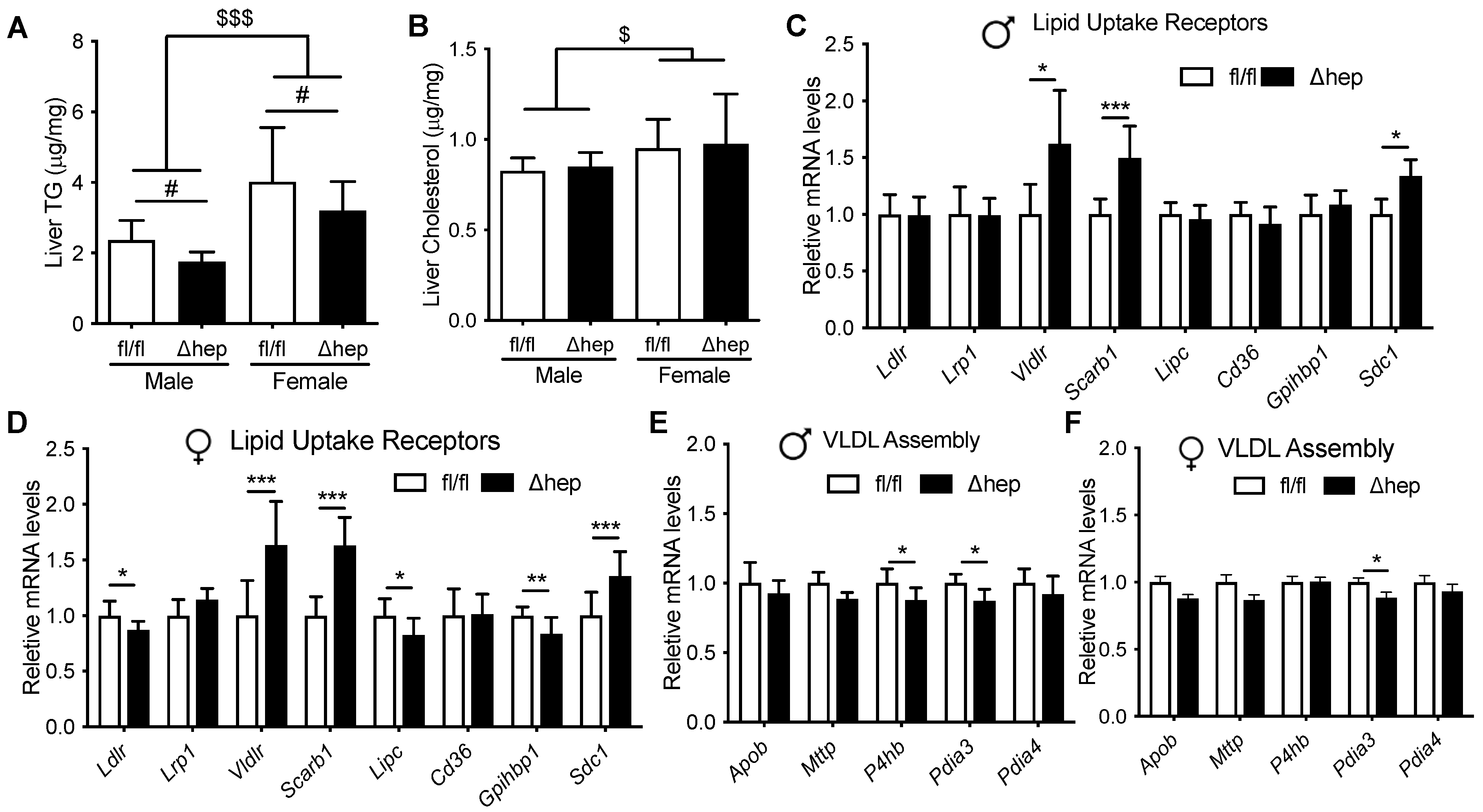
2.2. Metabolic Changes after Hepatic Deletion of SHP
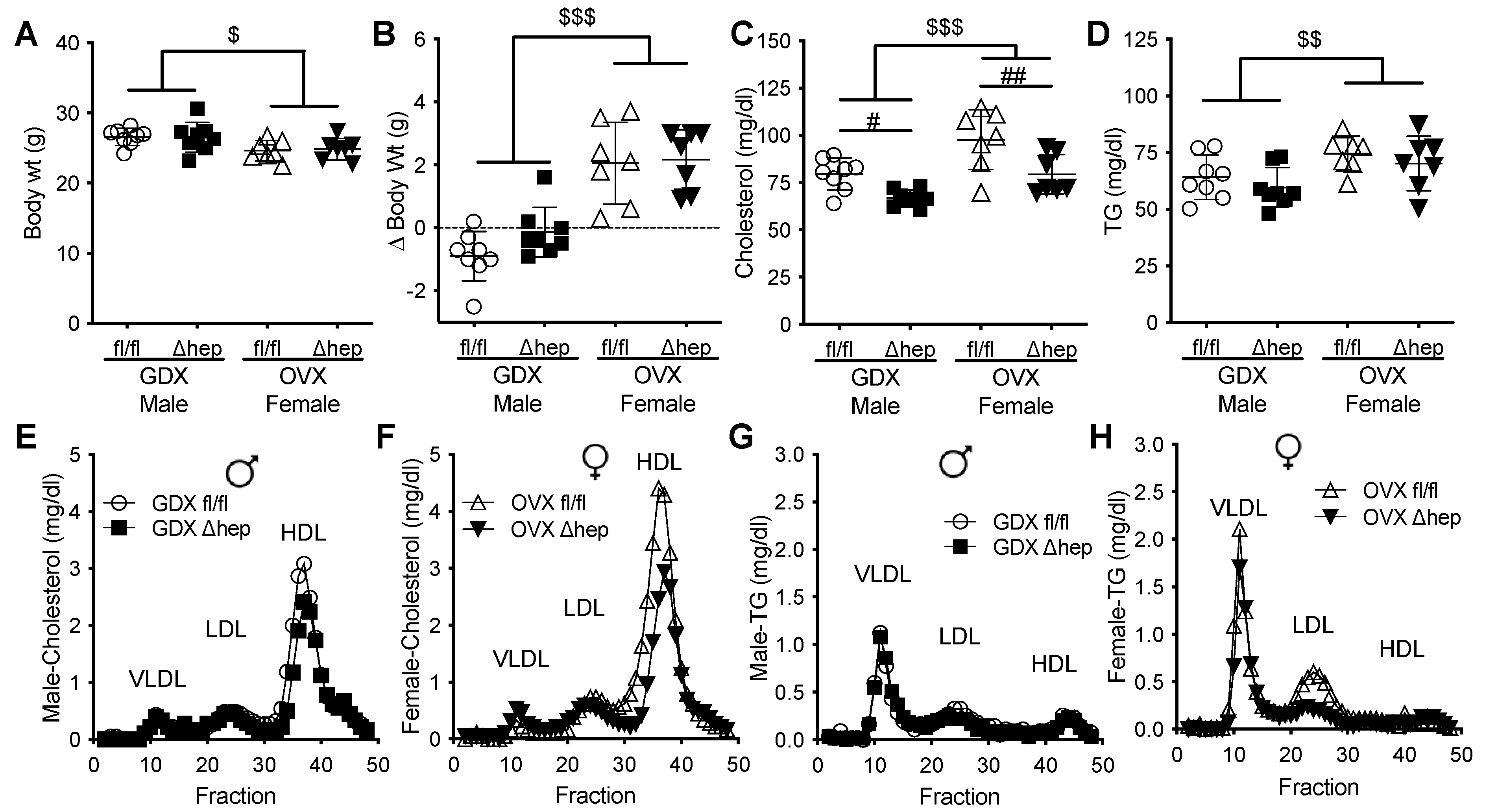
2.3. Effect of Gonadectomy on Hepatic SHP Regulation of Plasma TGs in Males and Females
2.4. Hepatic Deletion of SHP Promotes TG Clearance in Hypertriglyceridemic Mice
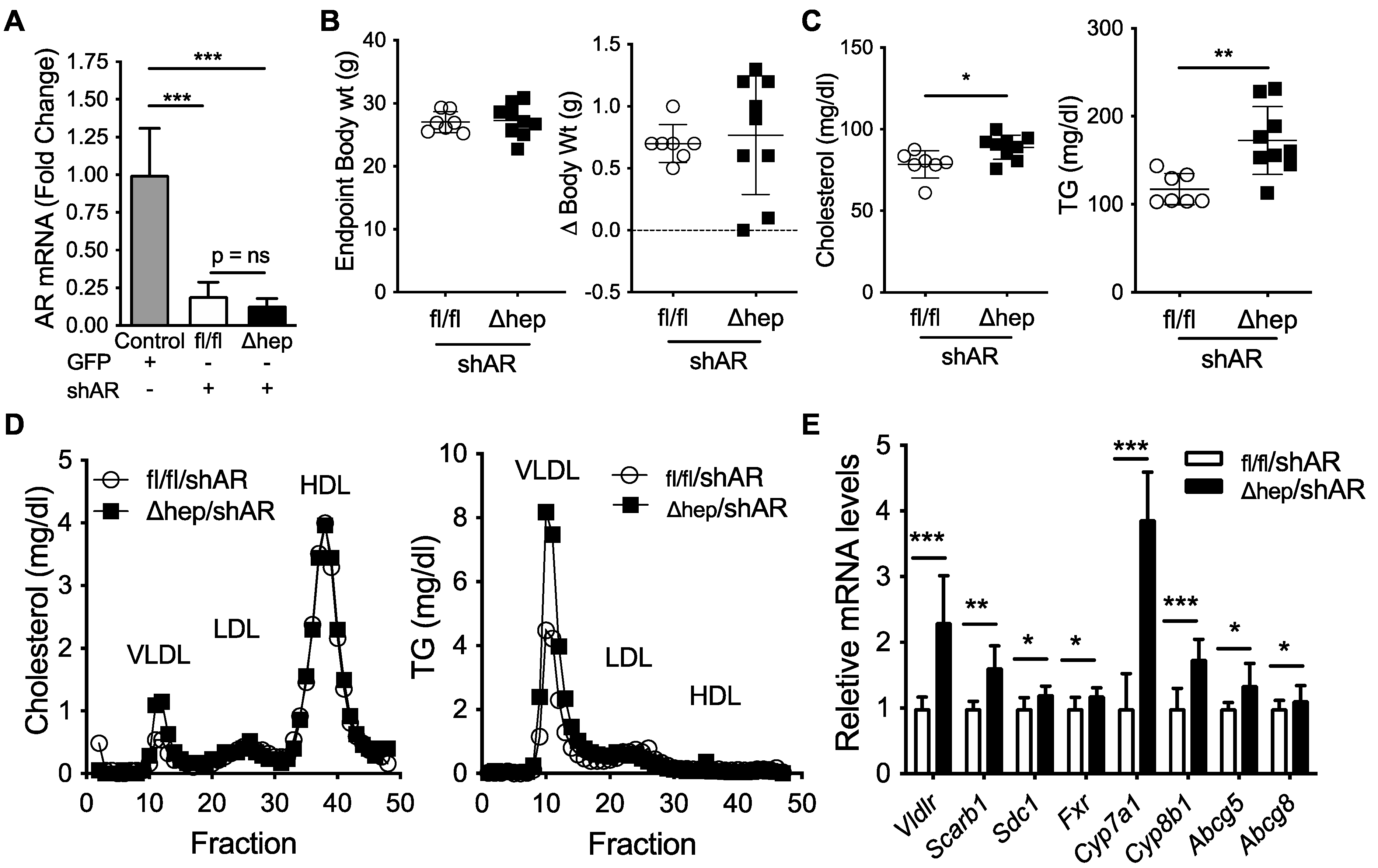
2.5. Hepatic SHP Regulates Plasma TGs through Androgen Receptor
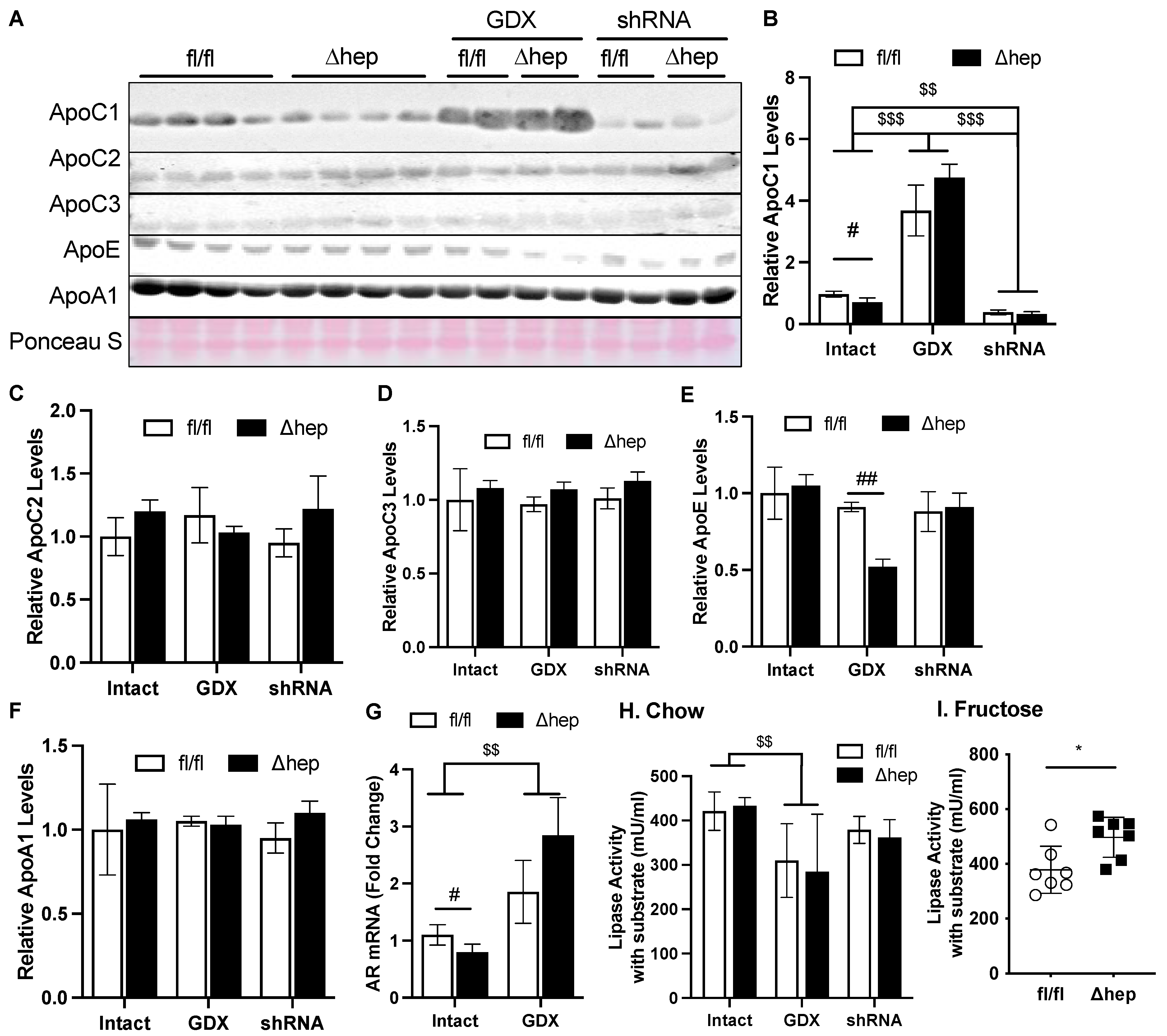
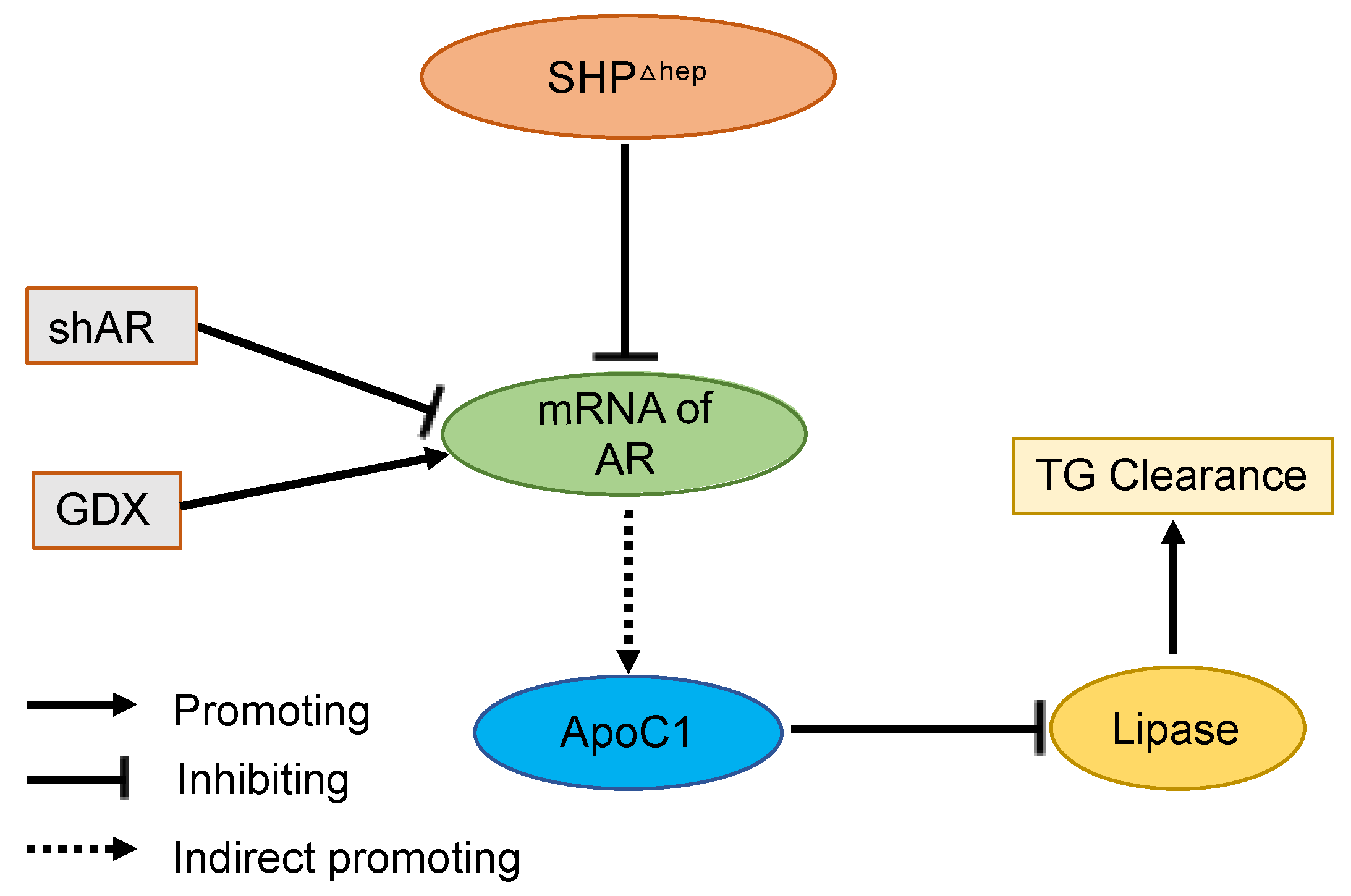
2.6. Hepatic SHP Regulates TG Clearance via ApoC1 and Involves Regulation by Liver AR
3. Discussion
4. Materials and Methods
4.1. Animals, Genotyping, and Viral Knockdown
4.2. Gonadectomy and Ovariectomy Surgery
4.3. Lipid and Lipoprotein Analysis
4.4. In Vivo TG Production and Clearance
4.5. Western Blotting
4.6. Blood Lipase Activities
4.7. Liver and Adipose mRNA Quantification
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frick, M.H.; Elo, O.; Haapa, K.; Heinonen, O.P.; Heinsalmi, P.; Helo, P.; Huttunen, J.K.; Kaitaniemi, P.; Koskinen, P.; Manninen, V.; et al. Helsinki Heart Study: Primary-Prevention Trial with Gemfibrozil in Middle-Aged Men with Dyslipidemia. N. Engl. J. Med. 1987, 317, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Keech, A.C.; Simes, R.J.; Barter, P.J.; Best, J.D.; Scott, R.A.P.; Taskinen, M.R.; Forder, P.M.; Pillai, A.; Davis, T.M.; Glasziou, P.; et al. Effects of long-term fenofibrate therapy on cardiovascular events in 9795 people with type 2 diabetes mellitus (the FIELD study): Randomised controlled trial. Lancet 2005, 366, 1849–1861. [Google Scholar] [CrossRef]
- Rubins, H.B.; Robins, S.J.; Collins, D.; Fye, C.L.; Anderson, J.W.; Elam, M.B.; Faas, F.H.; Linares, E.; Schaefer, E.J.; Schectman, G.; et al. Gemfibrozil for the Secondary Prevention of Coronary Heart Disease in Men with Low Levels of High-Density Lipoprotein Cholesterol. N. Engl. J. Med. 1999, 341, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Bezafibrate Infarction Prevention Study Group. Secondary prevention by raising HDL cholesterol and reducing triglycerides in patients with coronary artery disease. Circulation 2000, 102, 21–27. [Google Scholar] [CrossRef]
- Holmes, M.V.; Asselbergs, F.W.; Palmer, T.M.; Drenos, F.; Lanktree, M.B.; Nelson, C.P.; Dale, C.E.; Padmanabhan, S.; Finan, C.; Swerdlow, D.I.; et al. Mendelian randomization of blood lipids for coronary heart disease. Eur. Heart J. 2015, 36, 539–550. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.J.; Peloso, G.M.; Yu, H.; Butterworth, A.S.; Wang, X.; Mahajan, A.; Saleheen, D.; Emdin, C.; Alam, D.; Alves, A.C.; et al. Exome-wide association study of plasma lipids in >300,000 individuals. Nat. Genet. 2017, 49, 1758–1766. [Google Scholar] [CrossRef] [Green Version]
- Thomsen, M.; Varbo, A.; Tybjærg-Hansen, A.; Nordestgaard, B.G. Low Nonfasting Triglycerides and Reduced All-Cause Mortality: A Mendelian Randomization Study. Clin. Chem. 2014, 60, 737–746. [Google Scholar] [CrossRef] [Green Version]
- Sarwar, N.; Sandhu, M.S.; Ricketts, S.L.; Butterworth, A.S.; Di Angelantonio, E.; Boekholdt, S.M.; Ouwehand, W.H.; Watkins, H.; Samani, N.J.; Saleheen, D.; et al. Triglyceride-mediated pathways and coronary disease: Collaborative analysis of 101 studies. Lancet 2010, 375, 1634–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adiels, M.; Olofsson, S.-O.; Taskinen, M.-R.; Borén, J. Overproduction of Very Low–Density Lipoproteins Is the Hallmark of the Dyslipidemia in the Metabolic Syndrome. Arter. Thromb. Vasc. Biol. 2008, 28, 1225–1236. [Google Scholar] [CrossRef]
- Sparks, J.D.; Sparks, C.E.; Adeli, K. Selective Hepatic Insulin Resistance, VLDL Overproduction, and Hypertriglyceridemia. Arter. Thromb. Vasc. Biol. 2012, 32, 2104–2112. [Google Scholar] [CrossRef] [Green Version]
- Chandler, C.E.; Wilder, D.E.; Pettini, J.L.; Savoy, Y.E.; Petras, S.F.; Chang, G.; Vincent, J.; Harwood, H.J. CP-346086: An MTP inhibitor that lowers plasma cholesterol and triglycerides in experimental animals and in humans. J. Lipid Res. 2003, 44, 1887–1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raal, F.J.; Santos, R.D.; Blom, D.J.; Marais, A.D.; Charng, M.-J.; Cromwell, W.C.; Lachmann, R.H.; Gaudet, D.; Tan, J.L.; Chasan-Taber, S.; et al. Mipomersen, an apolipoprotein B synthesis inhibitor, for lowering of LDL cholesterol concentrations in patients with homozygous familial hypercholesterolaemia: A randomised, double-blind, placebo-controlled trial. Lancet 2010, 375, 998–1006. [Google Scholar] [CrossRef]
- Nordestgaard, B.G.; Benn, M.; Schnohr, P.; Tybjaerg-Hansen, A. Nonfasting Triglycerides and Risk of Myocardial Infarction, Ischemic Heart Disease, and Death in Men and Women. JAMA 2007, 298, 299–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boquist, S.; Ruotolo, G.; Tang, R.; Björkegren, J.; Bond, M.G.; de Faire, U.; Karpe, F.; Hamsten, A. Alimentary Lipemia, Postprandial Triglyceride-Rich Lipoproteins, and Common Carotid Intima-Media Thickness in Healthy, Middle-Aged Men. Circulation 1999, 100, 723–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carstensen, M.; Thomsen, C.; Gotzsche, O.; Holst, J.J.; Schrezenmeir, J.; Hermansen, K. Differential Postprandial Lipoprotein Responses in Type 2 Diabetic Men with and without Clinical Evidence of a Former Myocardial Infarction. Rev. Diabet. Stud. 2004, 1, 175. [Google Scholar] [CrossRef] [Green Version]
- Ginsberg, H.N.; Jones, J.; Blaner, W.S.; Thomas, A.; Karmally, W.; Fields, L.; Blood, D.; Begg, M.D. Association of Postprandial Triglyceride and Retinyl Palmitate Responses with Newly Diagnosed Exercise-Induced Myocardial Ischemia in Middle-Aged Men and Women. Arter. Thromb. Vasc. Biol. 1995, 15, 1829–1838. [Google Scholar] [CrossRef]
- Groot, P.H.; Van Stiphout, W.A.; Krauss, X.H.; Jansen, H.; Van Tol, A.; Van Ramshorst, E.; Chin-On, S.; Hofman, A.; Cresswell, S.R.; Havekes, L. Postprandial lipoprotein metabolism in normolipidemic men with and without coronary artery disease. Arter. Thromb. A J. Vasc. Biol. 1991, 11, 653–662. [Google Scholar] [CrossRef] [Green Version]
- Sharrett, A.R.; Chambless, L.E.; Heiss, G.; Paton, C.C.; Patsch, W. Association of Postprandial Triglyceride and Retinyl Palmitate Responses With Asymptomatic Carotid Artery Atherosclerosis in Middle-aged Men and Women. Arter. Thromb. Vasc. Biol. 1995, 15, 2122–2129. [Google Scholar] [CrossRef]
- Gaudet, D.; Gipe, D.A.; Pordy, R.; Ahmad, Z.; Cuchel, M.; Shah, P.K.; Chyu, K.-Y.; Sasiela, W.J.; Chan, K.-C.; Brisson, D.; et al. ANGPTL3 Inhibition in Homozygous Familial Hypercholesterolemia. N. Engl. J. Med. 2017, 377, 296–297. [Google Scholar] [CrossRef]
- Graham, M.J.; Lee, R.G.; Brandt, T.A.; Tai, L.-J.; Fu, W.; Peralta, R.; Yu, R.; Hurh, E.; Paz, E.; McEvoy, B.W.; et al. Cardiovascular and Metabolic Effects of ANGPTL3 Antisense Oligonucleotides. N. Engl. J. Med. 2017, 377, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, D.; Alexander, V.J.; Baker, B.F.; Brisson, D.; Tremblay, K.; Singleton, W.; Geary, R.S.; Hughes, S.G.; Viney, N.J.; Graham, M.J.; et al. Antisense Inhibition of Apolipoprotein C-III in Patients with Hypertriglyceridemia. N. Engl. J. Med. 2015, 373, 438–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaefer, E.J.; Lamon-Fava, S.; Cohn, S.D.; Schaefer, M.M.; Ordovas, J.M.; Castelli, W.P.; Wilson, P.W. Effects of age, gender, and menopausal status on plasma low density lipoprotein cholesterol and apolipoprotein B levels in the Framingham Offspring Study. J. Lipid Res. 1994, 35, 779–792. [Google Scholar] [CrossRef]
- Couillard, C.; Bergeron, N.; Prud’homme, D.; Bergeron, J.; Tremblay, A.; Bouchard, C.; Mauriege, P.; Despres, J.P. Gender difference in postprandial lipemia: Importance of visceral adipose tissue accumulation. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 2448–2455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittendorfer, B.; Patterson, B.W.; Klein, S. Effect of sex and obesity on basal VLDL-triacylglycerol kinetics. Am. J. Clin. Nutr. 2003, 77, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Cianflone, K.; Zakarian, R.; Couillard, C.; Delplanque, B.; Despres, J.-P.; Sniderman, A. Fasting acylation-stimulating protein is predictive of postprandial triglyceride clearance. J. Lipid Res. 2004, 45, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Horton, T.J.; Commerford, S.R.; Pagliassotti, M.J.; Bessesen, D.H. Postprandial leg uptake of triglyceride is greater in women than in men. Am. J. Physiol. Metab. 2002, 283, E1192–E1202. [Google Scholar] [CrossRef] [Green Version]
- Båvner, A.; Sanyal, S.; Gustafsson, J.-Å.; Treuter, E. Transcriptional corepression by SHP: Molecular mechanisms and physiological consequences. Trends Endocrinol. Metab. 2005, 16, 478–488. [Google Scholar] [CrossRef]
- Zhang, Y.; Hagedorn, C.H.; Wang, L. Role of nuclear receptor SHP in metabolism and cancer. Biochim. Biophys. Acta Mol. Basis Dis. 2011, 1812, 893–908. [Google Scholar] [CrossRef] [Green Version]
- Lu, T.T.; Makishima, M.; Repa, J.J.; Schoonjans, K.; Kerr, T.A.; Auwerx, J.; Mangelsdorf, D.J. Molecular Basis for Feedback Regulation of Bile Acid Synthesis by Nuclear Receptors. Mol. Cell 2000, 6, 507–515. [Google Scholar] [CrossRef]
- Park, Y.J.; Kim, S.C.; Kim, J.; Anakk, S.; Lee, J.M.; Tseng, H.-T.; Yechoor, V.; Park, J.; Choi, J.-S.; Jang, H.C.; et al. Dissociation of diabetes and obesity in mice lacking orphan nuclear receptor small heterodimer partner. J. Lipid Res. 2011, 52, 2234–2244. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Iqbal, J.; Saha, P.K.; Liu, J.; Chan, L.; Hussain, M.M.; Moore, D.D.; Wang, L. Molecular characterization of the role of orphan receptor small heterodimer partner in development of fatty liver. Hepatology 2007, 46, 147–157. [Google Scholar] [CrossRef]
- Wang, L.; Liu, J.; Saha, P.; Huang, J.; Chan, L.; Spiegelman, B.; Moore, D.D. The orphan nuclear receptor SHP regulates PGC-1α expression and energy production in brown adipocytes. Cell Metab. 2005, 2, 227–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, M.; Houten, S.; Wang, L.; Moschetta, A.; Mangelsdorf, D.J.; Heyman, R.A.; Moore, D.D.; Auwerx, J. Bile acids lower triglyceride levels via a pathway involving FXR, SHP, and SREBP-1c. J. Clin. Investig. 2004, 113, 1408–1418. [Google Scholar] [CrossRef] [Green Version]
- Seol, W.; Hanstein, B.; Brown, M.; Moore, D.D. Inhibition of Estrogen Receptor Action by the Orphan Receptor SHP (Short Heterodimer Partner). Mol. Endocrinol. 1998, 12, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Gobinet, J.; Auzou, G.; Nicolas, J.-C.; Sultan, C.; Jalaguier, S. Characterization of the Interaction between Androgen Receptor and a New Transcriptional Inhibitor, SHP. Biochem. Us 2001, 40, 15369–15377. [Google Scholar] [CrossRef]
- Tseng, H.-T.; Park, Y.J.; Lee, Y.K.; Moore, D.D. The orphan nuclear receptor small heterodimer partner is required for thiazolidinedione effects in leptin-deficient mice. J. Biomed. Sci. 2015, 22, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, A.; Magee, N.; Deng, F.; Lehn, S.; Zhong, C.; Zhang, Y. Hepatocyte nuclear receptor SHP suppresses inflammation and fibrosis in a mouse model of nonalcoholic steatohepatitis. J. Biol. Chem. 2018, 293, 8656–8671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magee, N.; Zou, A.; Ghosh, P.; Ahamed, F.; Delker, D.; Zhang, Y. Disruption of hepatic small heterodimer partner induces dissociation of steatosis and inflammation in experimental nonalcoholic steatohepatitis. J. Biol. Chem. 2020, 295, 994–1008. [Google Scholar] [CrossRef]
- Kerr, T.A.; Saeki, S.; Schneider, M.; Schaefer, K.; Berdy, S.; Redder, T.; Shan, B.; Russell, D.W.; Schwarz, M. Loss of Nuclear Receptor SHP Impairs but Does Not Eliminate Negative Feedback Regulation of Bile Acid Synthesis. Dev. Cell 2002, 2, 713–720. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Han, Y.; Kim, C.-S.; Lee, Y.-K.; Moore, D.D. Resistance of SHP-null Mice to Bile Acid-induced Liver Damage. J. Biol. Chem. 2003, 278, 44475–44481. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.J.; Qatanani, M.; Chua, S.S.; LaRey, J.L.; Johnson, S.A.; Watanabe, M.; Moore, D.D.; Lee, Y.K. Loss of orphan receptor small heterodimer partner sensitizes mice to liver injury from obstructive cholestasis. Hepatology 2008, 47, 1578–1586. [Google Scholar] [CrossRef]
- Anakk, S.; Watanabe, M.; Ochsner, S.A.; McKenna, N.J.; Finegold, M.J.; Moore, D.D. Combined deletion of Fxr and Shp in mice induces Cyp17a1 and results in juvenile onset cholestasis. J. Clin. Investig. 2011, 121, 86–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, G.; Zhang, T.; Yu, S.; Lee, S.; Calabuig-Navarro, V.; Yamauchi, J.; Ringquist, S.; Dong, H.H. ATF4 Protein Deficiency Protects against High Fructose-induced Hypertriglyceridemia in Mice. J. Biol. Chem. 2013, 288, 25350–25361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmisano, B.T.; Le, T.D.; Zhu, L.; Lee, Y.K.; Stafford, J.M. Cholesteryl ester transfer protein alters liver and plasma triglyceride metabolism through two liver networks in female mice. J. Lipid Res. 2016, 57, 1541–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenney, R.M.; Bell, C.L.; Wilson, J.M. AAV8 capsid variable regions at the two-fold symmetry axis contribute to high liver transduction by mediating nuclear entry and capsid uncoating. Virology. 2014, 454–455, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Fuior, E.V.; Gafencu, A.V. Apolipoprotein C1: Its Pleiotropic Effects in Lipid Metabolism and Beyond. Int. J. Mol. Sci. 2019, 20, 5939. [Google Scholar] [CrossRef] [Green Version]
- Jong, M.C.; Dahlmans, V.E.; Van Gorp, P.J.; Breuer, M.L.; Mol, M.J.; Van Der Zee, A.; Frants, R.R.; Hofker, M.H.; Havekes, L.M. Both Lipolysis and Hepatic Uptake of VLDL Are Impaired in Transgenic Mice Coexpressing Human Apolipoprotein E*3Leiden and Human Apolipoprotein C1. Arter. Thromb. Vasc. Biol. 1996, 16, 934–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, L.; Brown, W.C.; Cai, Q.; Krust, A.; Chambon, P.; McGuinness, O.P.; Stafford, J.M. Estrogen Treatment After Ovariectomy Protects Against Fatty Liver and May Improve Pathway-Selective Insulin Resistance. Diabetes 2012, 62, 424–434. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lu, Y.; Wang, E.; Zhang, Z.; Xiong, X.; Zhang, H.; Lu, J.; Zheng, S.; Yang, J.; Xia, X.; et al. Hepatic estrogen receptor α improves hepatosteatosis through upregulation of small heterodimer partner. J. Hepatol. 2015, 63, 183–190. [Google Scholar] [CrossRef]
- Thompson, P.D.; Cullinane, E.M.; Sady, S.P.; Chenevert, C.; Saritelli, A.L.; Sady, M.A.; Herbert, P.N. Contrasting Effects of Testosterone and Stanozolol on Serum Lipoprotein Levels. JAMA 1989, 261, 1165–1168. [Google Scholar] [CrossRef]
- Snyder, P.J.; Peachey, H.; Berlin, J.A.; Hannoush, P.; Haddad, G.; Dlewati, A.; Santanna, J.; Loh, L.; Lenrow, D.A.; Holmes, J.H.; et al. Effects of Testosterone Replacement in Hypogonadal Men. J. Clin. Endocrinol. Metab. 2000, 85, 2670–2677. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.B.; Hsia, S.; Alaupovic, P.; Sinha-Hikim, I.; Woodhouse, L.; Buchanan, T.A.; Shen, R.; Bross, R.; Berman, N.; Bhasin, S. The Effects of Varying Doses of T on Insulin Sensitivity, Plasma Lipids, Apolipoproteins, and C-Reactive Protein in Healthy Young Men. J. Clin. Endocrinol. Metab. 2002, 87, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Conde-Knape, K.; Bensadoun, A.; Sobel, J.H.; Cohn, J.S.; Shachter, N.S. Overexpression of apoC-I in apoE-null mice. J. Lipid Res. 2002, 43, 2136–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, K.; Harnish, D.C.; Evans, M.J. Estrogen Receptor α Regulates Expression of the Orphan Receptor Small Heterodimer Partner. J. Biol. Chem. 2003, 278, 36418–36429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poensgen, J. Apolipoprotein C-1 inhibits the hydrolysis by phospholipase A2 of phospholipids in liposomes and cell membranes. Biochim. Biophys. Acta Lipids Lipid Metab. 1990, 1042, 188–192. [Google Scholar] [CrossRef]
- Pan, X.; Zhang, Y.; Wang, L.; Hussain, M.M. Diurnal Regulation of MTP and Plasma Triglyceride by CLOCK Is Mediated by SHP. Cell Metab. 2010, 12, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Berbée, J.F.P.; van der Hoogt, C.C.; Sundararaman, D.; Havekes, L.M.; Rensen, P.C.N. Severe hypertriglyceridemia in human APOC1 transgenic mice is caused by apoC-I-induced inhibition of LPL. J. Lipid Res. 2005, 46, 297–306. [Google Scholar] [CrossRef] [Green Version]
- Rosenson, R.S.; Davidson, M.H.; Hirsh, B.J.; Kathiresan, S.; Gaudet, D. Genetics and Causality of Triglyceride-Rich Lipoproteins in Atherosclerotic Cardiovascular Disease. J. Am. Coll. Cardiol. 2014, 64, 2525–2540. [Google Scholar] [CrossRef] [Green Version]
- Marcel, Y.; Bergseth, M.; Nestruck, A. Preparative isoelectric focussing of apolipoproteins C and E from human very low density lipoproteins. Biochim. Biophys. Acta Lipids Lipid Metab. 1979, 573, 175–183. [Google Scholar] [CrossRef]
- Simonet, W.; Bucay, N.; Pitas, R.; Lauer, S.; Taylor, J. Multiple tissue-specific elements control the apolipoprotein E/C-I gene locus in transgenic mice. J. Biol. Chem. 1991, 266, 8651–8654. [Google Scholar] [CrossRef]
- Palmisano, B.T.; Anozie, U.; Yu, S.; Neuman, J.C.; Zhu, L.; Edington, E.M.; Luu, T.; Stafford, J.M. Cholesteryl Ester Transfer Protein Impairs Triglyceride Clearance via Androgen Receptor in Male Mice. Lipids 2021, 56, 17–29. [Google Scholar] [CrossRef]
- Zhu, L.; Luu, T.; Emfinger, C.H.; Parks, B.A.; Shi, J.; Trefts, E.; Zeng, F.; Kuklenyik, Z.; Harris, R.C.; Wasserman, D.H.; et al. CETP Inhibition Improves HDL Function but Leads to Fatty Liver and Insulin Resistance in CETP-Expressing Transgenic Mice on a High-Fat Diet. Diabetes 2018, 67, 2494–2506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Ruijter, J.M.; Ramakers, C.; Hoogaars, W.M.H.; Karlen, Y.; Bakker, O.; Hoff, M.J.B.V.D.; Moorman, A.F.M. Amplification efficiency: Linking baseline and bias in the analysis of quantitative PCR data. Nucleic Acids Res. 2009, 37, e45. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| Abcg5 | CCTGCAGAGCGACGTTTTTC | GCATCGCTGTGTATCGCAAC |
| Abcg8 | TGGATAGTGCCTGCATGGATC | AATTGAATCTGCATCAGCCCC |
| Apob | GCCCATTGTGGACAAGTTGATC | CCAGGACTTGGAGGTCTTGGA |
| Ar | GGACCATGTTTTACCCATCG | TCGTTTCTGCTGGCACATAG |
| Cd36 | TGGCCTTACTTGGGATTGG | CCAGTGTATATGTAGGCTCATCCA |
| Cyp7a1 | AGCAACTAAACAACCTGCCAGTACTA | GTCCGGATATTCAAGGATGCA |
| Cyp7b1 | TAGCCCTCTTTCCTCCACTCATA | GAACCGATCGAACCTAAATTCCT |
| Cyp8b1 | GCCTTCAAGTATGATCGGTTCCT | GATCTTCTTGCCCGACTTGTAGA |
| Fxr | TCCGGACATTCAACCATCAC | TCACTGCACATCCCAGATCTC |
| Gpihbp1 | TACCTACTCCATGTGGTGTACTG | AGGATGTCTAGTCCCACTTTCC |
| Ldlr | GCATCAGCTTGGACAAGGTGT | GGGAACAGCCACCATTGTTG |
| Lipc | GACGGGAAGAACAAGATTGGAA | GCATCATCAGGAGAAAGG |
| Lrp1 | TCAGACGAGCCTCCAGACTGT | ACAGATGAAGGCAGGGTTGGT |
| Mttp | CAAGCTCACGTACTCCACTGAAG | TCATCATCACCATCAGGATTCCT |
| P4hb | GCCGCAAAACTGAAGGCAG | GGTAGCCACGGACACCATAC |
| Pdia3 | CGCCTCCGATGTGTTGGAA | CAGTGCAATCCACCTTTGCTAA |
| Pdia4 | TCCCATTGCTGTAGCGAAGAT | GGGGTAGCCACTCACATCAAAT |
| Pparα | TATTCGGCTGAAGCTGGTGTAC | CTGGCATTTGTTCCGGTTCT |
| Ppia | CGATGACGAGCCCTTGG | TCTGCTGTCTTTGGAACTTTGTC |
| Scarb1 | TCAGAAGCTGTTCTTGGTCTGAAC | GTTCATGGGGATCCCAGTGA |
| Sdc1 | CTTTGTCACGGCAGACACCTT | GACAGAGGTAAAAGCAGTCTCG |
| Shp | CGATCCTCTTCAACCCAGATG | AGGGCTCCAAGACTTCACACA |
| Vldlr | CCACAGCAGTATCAGAAGTCAGTGT | CACCTACTGCTGCCATCACTAAGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palmisano, B.T.; Zhu, L.; Litts, B.; Burman, A.; Yu, S.; Neuman, J.C.; Anozie, U.; Luu, T.N.; Edington, E.M.; Stafford, J.M. Hepatocyte Small Heterodimer Partner Mediates Sex-Specific Effects on Triglyceride Metabolism via Androgen Receptor in Male Mice. Metabolites 2021, 11, 330. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11050330
Palmisano BT, Zhu L, Litts B, Burman A, Yu S, Neuman JC, Anozie U, Luu TN, Edington EM, Stafford JM. Hepatocyte Small Heterodimer Partner Mediates Sex-Specific Effects on Triglyceride Metabolism via Androgen Receptor in Male Mice. Metabolites. 2021; 11(5):330. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11050330
Chicago/Turabian StylePalmisano, Brian T., Lin Zhu, Bridget Litts, Andreanna Burman, Sophia Yu, Joshua C. Neuman, Uche Anozie, Thao N. Luu, Emery M. Edington, and John M. Stafford. 2021. "Hepatocyte Small Heterodimer Partner Mediates Sex-Specific Effects on Triglyceride Metabolism via Androgen Receptor in Male Mice" Metabolites 11, no. 5: 330. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11050330