
Metabolomics Insights into Chemical Convergence in Xanthomonas perforans and Metabolic Changes Following Treatment with the Small Molecule Carvacrol

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Metabolomic Profiling of X. perforans, and Annotation
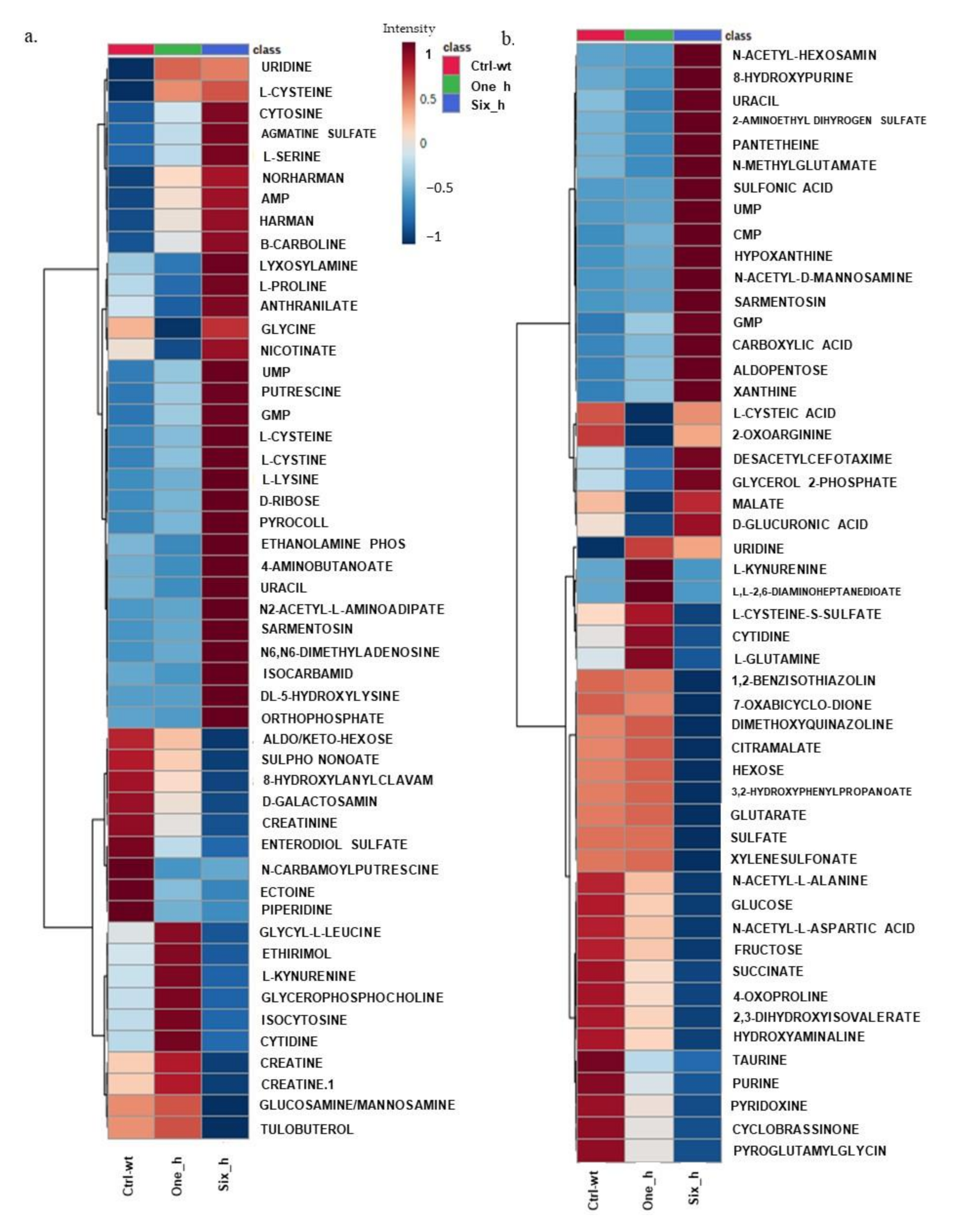
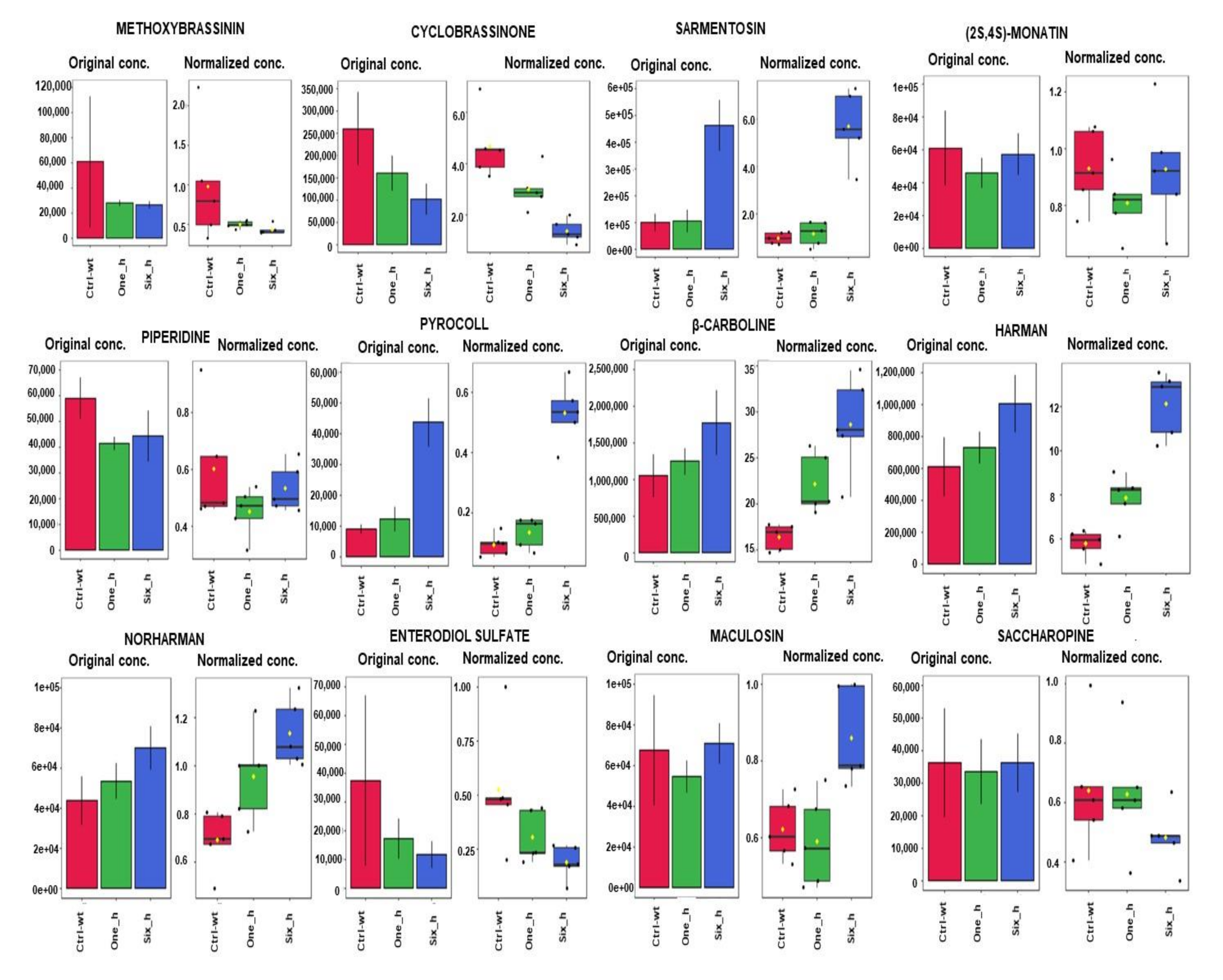
2.2. Effect of Carvacrol on Metabolic Changes in a Time Course Experiment in X. perforans
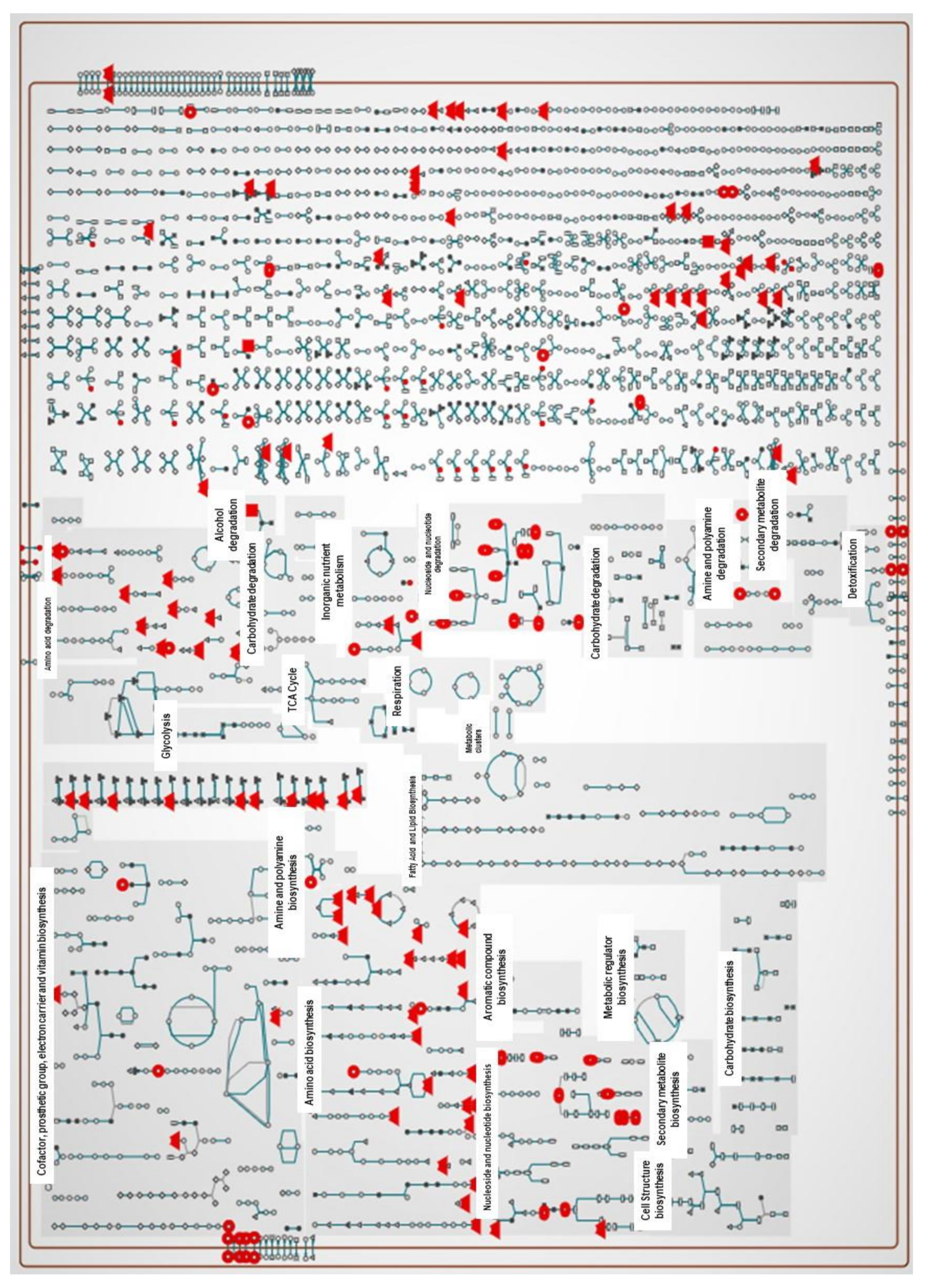
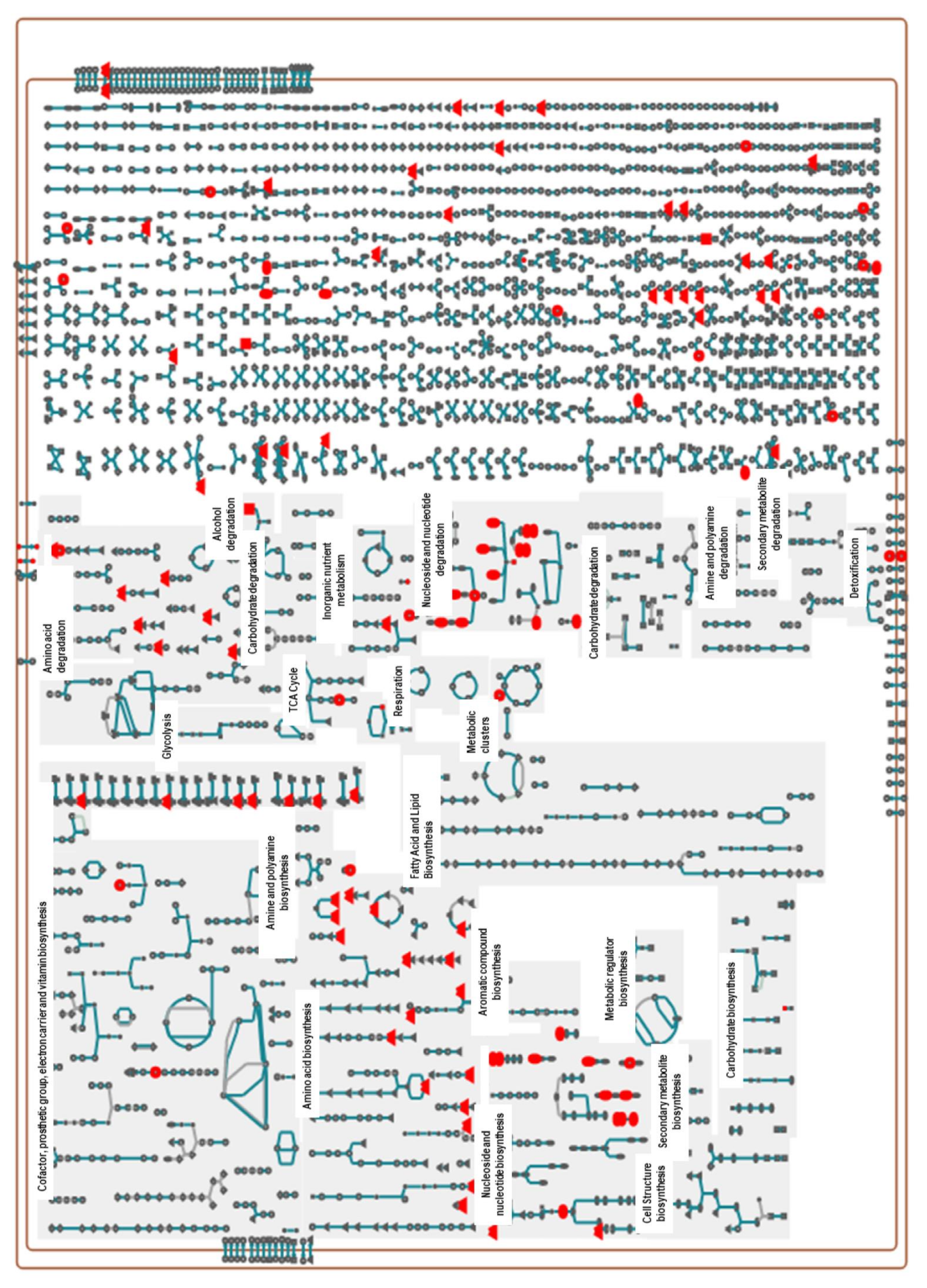
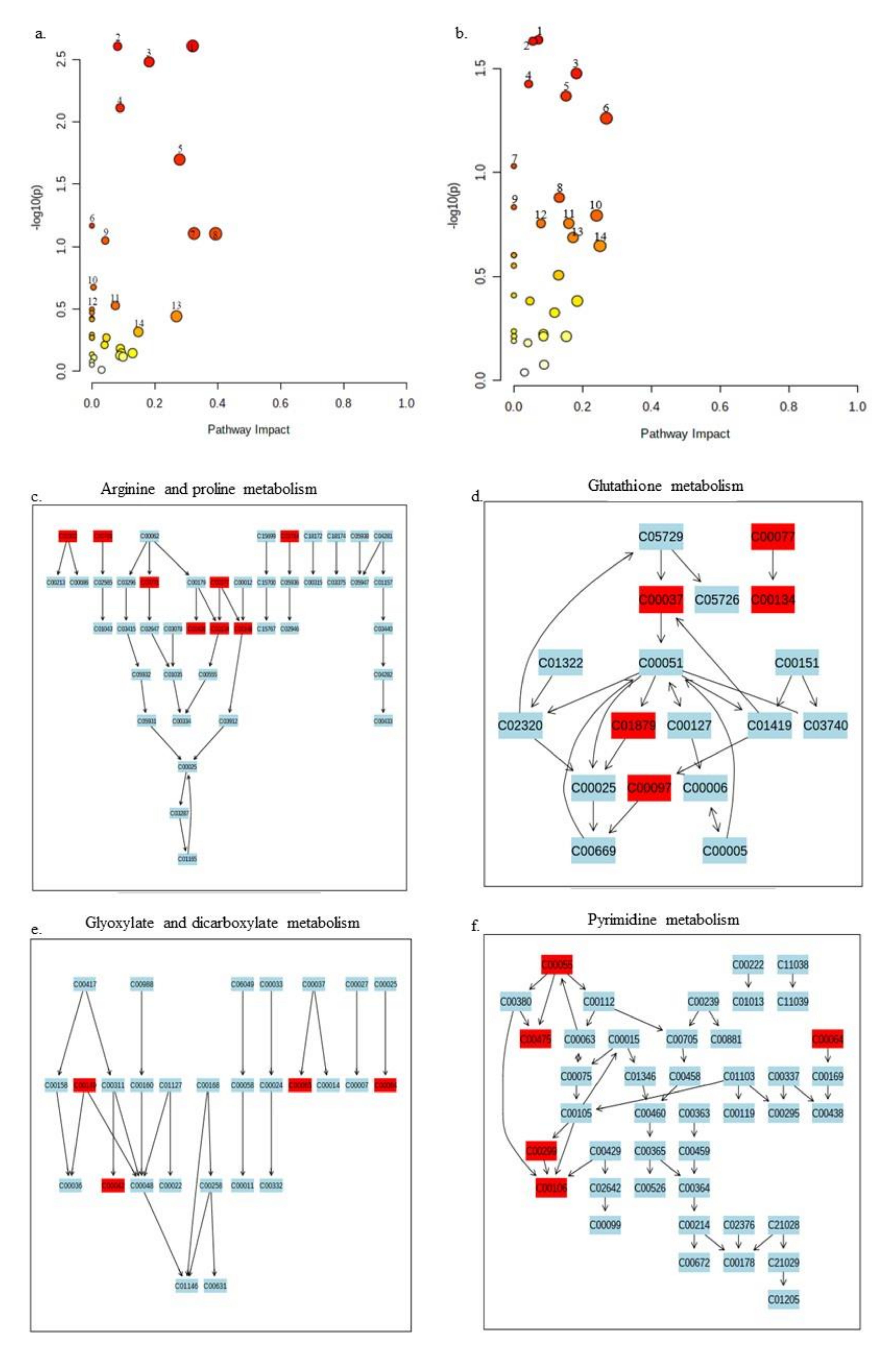
2.3. Pathway Analysis of Significant Metabolites of X. perforans
3. Discussion
3.1. LC-MS Identified Convergence of Chemicals from Diverse Systems in the Metabolome of X. perforans
3.2. Identified Metabolites as Novel Management Strategies for Xanthomonas Plant Pathogens
3.3. The Diversity of Metabolites in X. perforans Suggests the Possibility of Alternative Ecological Niches for Bacterial Spot Pathogens
3.4. Pathway Analyses Suggest That Carvacrol Elicits Global Metabolic Changes in X. perforans
4. Materials and Methods
4.1. Sample Preparation
4.2. Protein Precipitation and Metabolomics Profiling
4.3. Preliminary Data Analysis and Annotation
4.4. Univariate and Multivariate Analysis of Metabolic Pathway
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vincent, I.M.; Ehmann, D.E.; Mills, S.D.; Perros, M.; Barrett, M.P. Untargeted metabolomics to ascertain antibiotic modes of action. Antimicrob. Agents Chemother. 2016, 60, 2281–2291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiffman, C.; Petrick, L.; Perttula, K.; Yano, Y.; Carlsson, H.; Whitehead, T.; Metayer, C.; Hayes, J.; Rappaport, S.; Dudoit, S. Filtering procedures for untargeted LC-MS metabolomics data. BMC Bioinform. 2019, 20, 334. [Google Scholar] [CrossRef] [Green Version]
- Kellogg, J.; Kang, S. Metabolomics, an Essential Tool in Exploring and Harnessing Microbial Chemical Ecology. Phytobiomes J. 2020, 4, 195–210. [Google Scholar] [CrossRef]
- Sindelar, M.; Patti, G.J. Chemical Discovery in the Era of Metabolomics. J. Am. Chem. Soc. 2020, 142, 9097–9105. [Google Scholar] [CrossRef] [PubMed]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D.; McLean, J.A. Untargeted Metabolomics Strategies—Challenges and Emerging Directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manier, S.K.; Keller, A.; Schäper, J.; Meyer, M.R. Untargeted metabolomics by high resolution mass spectrometry coupled to normal and reversed phase liquid chromatography as a tool to study the in vitro biotransformation of new psychoactive substances. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Salem, M.A.; De Souza, L.P.; Serag, A.; Fernie, A.R.; Farag, M.A.; Ezzat, S.M.; Alseekh, S. Metabolomics in the Context of Plant Natural Products Research: From Sample Preparation to Metabolite Analysis. Metabolites 2020, 10, 37. [Google Scholar] [CrossRef] [Green Version]
- Domingo-Almenara, X.; Montenegro-Burke, J.R.; Benton, H.P.; Siuzdak, G. Annotation: A Computational Solution for Streamlining Metabolomics Analysis. Anal. Chem. 2018, 90, 480–489. [Google Scholar] [CrossRef] [Green Version]
- Pichersky, E.; Lewinsohn, E. Convergent Evolution in Plant Specialized Metabolism. Annu. Rev. Plant Biol. 2011, 62, 549–566. [Google Scholar] [CrossRef] [Green Version]
- Washburn, J.; Bird, K.; Conant, G.; Pires, J.C. Convergent Evolution and the Origin of Complex Phenotypes in the Age of Systems Biology. Int. J. Plant Sci. 2016, 177, 305–318. [Google Scholar] [CrossRef]
- Jensen, N.B.; Zagrobelny, M.; Hjernø, K.; Olsen, C.E.; Houghton-Larsen, J.; Borch, J.; Møller, B.L.; Bak, S. Convergent evolution in biosynthesis of cyanogenic defence compounds in plants and insects. Nat. Commun. 2011, 2, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, D. The genetic causes of convergent evolution. Nat. Rev. Genet. 2013, 14, 751–764. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; O’Donnell, A.J.; Barboline, J.J.; Barkman, T.J. Convergent evolution of caffeine in plants by co-option of exapted ancestral enzymes. Proc. Natl. Acad. Sci. USA 2016, 113, 10613–10618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beran, F.; Köllner, T.G.; Gershenzon, J.; Tholl, D. Chemical convergence between plants and insects: Biosynthetic origins and functions of common secondary metabolites. New Phytol. 2019, 223, 52–67. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Takahashi, E. Caffeine biosynthesis in Camellia sinensis. Phytochemistry 1976, 15, 1235–1239. [Google Scholar] [CrossRef]
- Ashihara, H.; Monteiro, A.M.; Gillies, F.M.; Crozier, A. Biosynthesis of Caffeine in Leaves of Coffee. Plant Physiol. 1996, 111, 747–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grenade, N.L.; Howe, G.W.; Ross, A.C. The convergence of bacterial natural products from evolutionarily distinct pathways. Curr. Opin. Biotechnol. 2021, 69, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Stringlis, I.A.; Zhang, H.; Pieterse, C.M.J.; Bolton, M.D.; de Jonge, R. Microbial small molecules—Weapons of plant subversion. Nat. Prod. Rep. 2018, 35, 410–433. [Google Scholar] [CrossRef] [Green Version]
- Pang, Z.; Chen, J.; Wang, T.; Gao, C.; Li, Z.; Guo, L.; Xu, J.; Cheng, Y. Linking Plant Secondary Metabolites and Plant Microbiomes: A Review. Front. Plant Sci. 2021, 12, 300. [Google Scholar] [CrossRef]
- Verpoorte, R.; Choi, Y.H.; Kim, H.K. Metabolomics: What’s new? Flavour Fragr. J. 2010, 25, 128–131. [Google Scholar] [CrossRef]
- Turi, C.; Finley, J.; Shipley, P.; Murch, S.J.; Brown, P.N. Metabolomics for Phytochemical Discovery: Development of Statistical Approaches Using a Cranberry Model System. J. Nat. Prod. 2015, 78, 953–966. [Google Scholar] [CrossRef]
- Caesar, L.K.; Kellogg, J.J.; Kvalheim, O.M.; Cech, N.B. Opportunities and Limitations for Untargeted Mass Spectrometry Metabolomics to Identify Biologically Active Constituents in Complex Natural Product Mixtures. J. Nat. Prod. 2019, 82, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Schirawski, J.; Perlin, M.H. Plant–Microbe Interaction 2017—The Good, the Bad and the Diverse. Int. J. Mol. Sci. 2018, 19, 1374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacoby, R.P.; Koprivova, A.; Kopriva, S. Pinpointing secondary metabolites that shape the composition and function of the plant microbiome. J. Exp. Bot. 2021, 72, 57–69. [Google Scholar] [CrossRef]
- Mine, A.; Sato, M.; Tsuda, K. Toward a systems understanding of plant–microbe interactions. Front. Plant Sci. 2014, 5, 423. [Google Scholar] [CrossRef]
- Rose, L.E.; Overdijk, E.J.R.; Van Damme, M. Small RNA molecules and their role in plant disease. Eur. J. Plant Pathol. 2019, 154, 115–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazarov, P.A.; Baleev, D.N.; Ivanova, M.I.; Sokolova, L.M.; Karakozova, M.V. Infectious plant diseases: Etiology, current status, problems and prospects in plant protection. Acta Nat. 2020, 12, 46–59. [Google Scholar] [CrossRef] [PubMed]
- Speth, E.B.; Lee, Y.N.; He, S.Y. Pathogen virulence factors as molecular probes of basic plant cellular functions. Curr. Opin. Plant Biol. 2007, 10, 580–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Baarlen, P.; Van Belkum, A.; Summerbell, R.C.; Crous, P.W.; Thomma, B.P. Molecular mechanisms of pathogenicity: How do pathogenic microorganisms develop cross-kingdom host jumps? FEMS Microbiol. Rev. 2007, 31, 239–277. [Google Scholar] [CrossRef] [Green Version]
- Neuhauser, S.; Kirchmair, M.; Bulman, S.; Bass, D. Cross-kingdom host shifts of phytomyxid parasites. BMC Evol. Biol. 2014, 14, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.; Bhatt, D. The circadian clock and defence signalling in plants. Mol. Plant Pathol. 2014, 16, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Mhlongo, M.I.; Piater, L.A.; Madala, N.E.; Labuschagne, N.; Dubery, I.A. The Chemistry of Plant–Microbe Interactions in the Rhizosphere and the Potential for Metabolomics to Reveal Signaling Related to Defense Priming and Induced Systemic Resistance. Front. Plant Sci. 2018, 9, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, V.S. Interrogating Plant–Microbe Interactions with Chemical Tools: Click Chemistry Reagents for Metabolic Labeling and Activity-Based Probes. Molecules 2021, 26, 243. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.; Vorhölter, F.-J.; Potnis, N.; Jones, J.B.; Van Sluys, M.-A.; Bogdanove, A.J.; Dow, J.M. Pathogenomics of Xanthomonas: Understanding bacterium–plant interactions. Nat. Rev. Genet. 2011, 9, 344–355. [Google Scholar] [CrossRef]
- Timilsina, S.; Potnis, N.; Newberry, E.A.; Liyanapathiranage, P.; Iruegas-Bocardo, F.; White, F.F.; Goss, E.M.; Jones, J.B. Xanthomonas diversity, virulence and plant–pathogen interactions. Nat. Rev. Genet. 2020, 18, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Büttner, D.; Bonas, U. Regulation and secretion of Xanthomonasvirulence factors. FEMS Microbiol. Rev. 2010, 34, 107–133. [Google Scholar] [CrossRef] [Green Version]
- Royer, M.; Koebnik, R.; Marguerettaz, M.; Barbe, V.; Robin, G.P.; Brin, C.; Carrere, S.; Gomez, C.; Hügelland, M.; Völler, G.H.; et al. Genome mining reveals the genus Xanthomonas to be a promising reservoir for new bioactive non-ribosomally synthesized peptides. BMC Genom. 2013, 14, 658. [Google Scholar] [CrossRef] [Green Version]
- Katzen, F.; Ferreiro, D.U.; Oddo, C.G.; Ielmini, M.V.; Becker, A.; Pühler, A.; Ielpi, L. Xanthomonas campestris pv campestris gum Mutants: Effects on Xanthan Biosynthesis and Plant Virulence. J. Bacteriol. 1998, 180, 1607–1617. [Google Scholar] [CrossRef] [Green Version]
- Royer, M.; Costet, L.; Vivien, E.; Bes, M.; Cousin, A.; Damais, A.; Pieretti, I.; Savin, A.; Megessier, S.; Viard, M.; et al. Albicidin Pathotoxin produced by Xanthomonas albilineans is encoded by three large PKS and NRPS genes present in a gene cluster also containing several putative modifying, regulatory, and resistance genes. Mol. Plant Microbe Interact. 2004, 17, 414–427. [Google Scholar] [CrossRef] [Green Version]
- Hashimi, S.M.; Wall, M.K.; Smith, A.B.; Maxwell, A.; Birch, R.G. The Phytotoxin Albicidin is a Novel Inhibitor of DNA Gyrase. Antimicrob. Agents Chemother. 2007, 51, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Kretz, J.; Kerwat, D.; Schubert, V.; Grätz, S.; Pesic, A.; Semsary, S.; Cociancich, S.; Royer, M.; Süssmuth, R.D. Total Synthesis of Albicidin: A Lead Structure from Xanthomonas albilineansfor Potent Antibacterial Gyrase Inhibitors. Angew. Chem. Int. Ed. 2015, 54, 1969–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimi, S.M. Albicidin, a potent DNA gyrase inhibitor with clinical potential. J. Antibiot. 2019, 72, 785–792. [Google Scholar] [CrossRef] [PubMed]
- Pohronezny, K.; Volin, R.B. The effect of bacterial spot on yield and quality of fresh market tomatoes. Hort Sci. 1983, 18, 59–70. [Google Scholar]
- Stall, R.E.; Jones, J.B.; Minsavage, G.V. Durability of Resistance in Tomato and Pepper to Xanthomonads Causing Bacterial Spot. Annu. Rev. Phytopathol. 2009, 47, 265–284. [Google Scholar] [CrossRef] [PubMed]
- Potnis, N.; Timilsina, S.; Strayer, A.; Shantharaj, D.; Barak, J.D.; Paret, M.L.; Vallad, G.E.; Jones, J.B. Bacterial spot of tomato and pepper: Diverse Xanthomonasspecies with a wide variety of virulence factors posing a worldwide challenge. Mol. Plant Pathol. 2015, 16, 907–920. [Google Scholar] [CrossRef]
- Tudor-Nelson, S.M.; Minsavage, G.V.; Stall, R.E.; Jones, J.B. Bacteriocin-Like Substances from Tomato Race 3 Strains of Xanthomonas campestris pv vesicatoria. Phytopathology 2003, 93, 1415–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, D.M.; Stall, R.E.; Jones, J.B.; Pauly, M.H.; Vallad, G.E.; Dahlbeck, D.; Staskawicz, B.J.; Scott, J.W. Transgenic Resistance Confers Effective Field Level Control of Bacterial Spot Disease in Tomato. PLoS ONE 2012, 7, e42036. [Google Scholar] [CrossRef] [PubMed]
- Klein-Gordon, J.M.; Xing, M.Y.; Garrett, K.A.; Abrahamian, P.; Paret, M.L.; Minsavage, G.V.; Strayer-Scherer, A.L.; Fulton, J.C.; Timilsina, S.; Jones, J.B.; et al. Assessing Changes and Associations in the Xanthomonas perforans Population Across Florida Commercial Tomato Fields Via a Statewide Survey. Phytopathology 2021. [Google Scholar] [CrossRef]
- Torelli, E.; Aiello, D.; Polizzi, G.; Firrao, G.; Cirvilleri, G. Draft genome of a Xanthomonas perforans strain associated with pith necrosis. FEMS Microbiol. Lett. 2015, 362, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Osdaghi, E.; Taghavi, S.M.; Hamzehzarghani, H.; Fazliarab, A.; Lamichhane, J.R. Monitoring the occurrence of tomato bacterial spot and range of the causal agent Xanthomonas perforans in Iran. Plant Pathol. 2017, 66, 990–1002. [Google Scholar] [CrossRef]
- Jibrin, M.O.; Potnis, N.; Timilsina, S.; Minsavage, G.V.; Vallad, G.E.; Roberts, P.D.; Jones, J.B.; Goss, E.M. Genomic Inference of Recombination-Mediated Evolution in Xanthomonas euvesicatoria and X. perforans. Appl. Environ. Microbiol. 2018, 84, e00136-18. [Google Scholar] [CrossRef] [Green Version]
- Abrahamian, P.; Klein-Gordon, J.M.; Jones, J.B.; Vallad, G.E. Epidemiology, diversity, and management of bacterial spot of tomato caused by Xanthomonas perforans. Appl. Microbiol. Biotechnol. 2021, 105, 6143–6158. [Google Scholar] [CrossRef]
- Liu, Q.; Qiao, K.; Zhang, S. Potential of a Small Molecule Carvacrol in Management of Vegetable Diseases. Molecules 2019, 24, 1932. [Google Scholar] [CrossRef] [Green Version]
- Qiao, K.; Liu, Q.; Huang, Y.; Xia, Y.; Zhang, S. Management of bacterial spot of tomato caused by copper-resistant Xanthomonas perforans using a small molecule compound carvacrol. Crop. Prot. 2020, 132, 105114. [Google Scholar] [CrossRef]
- Monde, K.; Sasaki, K.; Shirata, A.; Takasugi, M. 4-Methoxybrassinin, a sulphur-containing phytoalexin from Brassica oleracea. Phytochemistry 1990, 29, 1499–1500. [Google Scholar] [CrossRef]
- Vicente, J.G.; Holub, E.B. Xanthomonas campestris pv Campestris (cause of black rot of crucifers) in the genomic era is still a worldwide threat to brassica crops. Mol. Plant Pathol. 2013, 14, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Yannai, S. Dictionary of Food Compounds with CD-ROM: Additives, Flavors, and Ingredients; Chapman & Hall/CRC: Boca Raton, FL, USA, 2004. [Google Scholar]
- Kirsch, G.; El-Sawy, E.; Abdelwahab, A.B. Utilization of 1H-Indole-3-carboxaldehyde as a Precursor for the Synthesis of Bioactive Indole Alkaloids. Synthesis 2018, 50, 4525–4538. [Google Scholar] [CrossRef]
- Bjarnholt, N.; Rook, F.; Motawia, M.S.; Cornett, C.; Jørgensen, C.; Olsen, C.E.; Jaroszewski, J.W.; Bak, S.; Møller, B.L. Diversification of an ancient theme: Hydroxynitrile glucosides. Phytochemistry 2008, 69, 1507–1516. [Google Scholar] [CrossRef] [PubMed]
- De Castro, É.C.P.; Zagrobelny, M.; Zurano, J.P.; Cardoso, M.Z.; Feyereisen, R.; Bak, S. Sequestration and biosynthesis of cyanogenic glucosides in passion vine butterflies and consequences for the diversification of their host plants. Ecol. Evol. 2019, 9, 5079–5093. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, E.R.; Rosato, Y.B. Genotypic characterization of xanthomonad strains isolated from passion fruit plants (Passiflora spp.) and their relatedness to different Xanthomonas species. Int. J. Syst. Evol. Microbiol. 2000, 50, 811–821. [Google Scholar] [CrossRef] [Green Version]
- Fry, J.C.; Yurttas, N.; Biermann, K.L.; Lindley, M.G.; Goulson, M.J. The Sweetness Concentration-Response of R,R-Monatin, a Naturally Occurring High-Potency Sweetener. J. Food Sci. 2012, 77, S362–S364. [Google Scholar] [CrossRef] [PubMed]
- Maharaj, V.; Moodley, N.; Vahrmeijer, H. Characterization of natural monatin isomers, a high intensity sweetener from the plant Sclerochiton ilicifolius from South Africa. South Afr. J. Bot. 2018, 115, 37–43. [Google Scholar] [CrossRef]
- Li, S.; Lei, Y.; Jia, Y.; Li, N.; Wink, M.; Ma, Y. Piperine, a piperidine alkaloid from Piper nigrum re-sensitizes P-gp, MRP1 and BCRP dependent multidrug resistant cancer cells. Phytomedicine 2011, 19, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Schnabel, A.; Athmer, B.; Manke, K.; Schumacher, F.; Cotinguiba, F.; Vogt, T. Identification and characterization of piperine synthase from black pepper, Piper nigrum L. Commun. Biol. 2021, 4, 1–10. [Google Scholar] [CrossRef]
- Leyns, F.; Marcel, D.C.; Jean-Guy, S.; De Ley, J. The Host Range of the Genus Xanthomonas. Bot. Rev. 1984, 50, 308–356. [Google Scholar] [CrossRef]
- Saarinen, N.M.; Smeds, A.I.; Peñalvo, J.L.; Nurmi, T.; Adlercreutz, H.; Makela, S. Flaxseed Ingestion Alters Ratio of Enterolactone Enantiomers in Human Serum. J. Nutr. Metab. 2010, 2010, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Wrońska, A.K.; Boguś, M.I. Harman and norharman, metabolites of the entomopathogenic fungus Conidiobolus coronatus (Entomophthorales), affect the serotonin levels and phagocytic activity of hemocytes, insect immunocompetent cells, in Galleria mellonella (Lepidoptera). Cell BioSci. 2019, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, B.; Zhang, Q.; Zhang, J.; Wang, J.; Wu, W. Synthesis and bioactivity of beta-carboline derivatives. Nat. Prod. Commun. 2010, 5, 1591–1596. [Google Scholar]
- Nenaah, G. Antibacterial and antifungal activities of (beta)-carboline alkaloids of Peganum harmala (L) seeds and their combination effects. Fitoterapia 2010, 81, 779–782. [Google Scholar] [CrossRef]
- Dietera, A.; Hamm, A.; Fiedler, H.-P.; Goodfellow, M.; Müller, W.E.G.; Brun, R.; Beil, W.; Bringmann, G. Pyrocoll, an Antibiotic, Antiparasitic and Antitumor Compound Produced by a Novel Alkaliphilic Streptomyces Strain. J. Antibiot. 2003, 56, 639–646. [Google Scholar] [CrossRef] [Green Version]
- Stierle, A.C.; Cardellina, J.H.; Strobel, G.A. Maculosin, a host-specific phytotoxin for spotted knapweed from Alternaria alternata. Proc. Natl. Acad. Sci. USA 1988, 85, 8008–8011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saunders, P.; Broquist, H.P. Saccharopine, an Intermediate of the Aminoadipic Acid Pathway of Lysine Biosynthesis. J. Biol. Chem. 1966, 241, 3435–3440. [Google Scholar] [CrossRef]
- Liu, H.-W.; Ji, Q.-T.; Ren, G.-G.; Wang, F.; Su, F.; Wang, P.-Y.; Zhou, X.; Wu, Z.-B.; Li, Z.; Yang, S. Antibacterial Functions and Proposed Modes of Action of Novel 1,2,3,4-Tetrahydro-β-carboline Derivatives that Possess an Attractive 1,3-Diaminopropan-2-ol Pattern against Rice Bacterial Blight, Kiwifruit Bacterial Canker, and Citrus Bacterial Canker. J. Agric. Food Chem. 2020, 68, 12558–12568. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Xiang, M.; Luo, M.; Liu, H.; Zhou, X.; Wu, Z.; Liu, L.; Li, Z.; Yang, S. Novel piperazine-tailored ursolic acid hybrids as significant antibacterial agents targeting phytopathogens Xanthomonas oryzae pv oryzae and X. axonopodis pv. citri probably directed by activation of apoptosis. Pest Manag. Sci. 2020, 76, 2746–2754. [Google Scholar] [CrossRef]
- Xiang, M.; Song, Y.; Ji, J.; Zhou, X.; Liu, L.; Wang, P.; Wu, Z.; Li, Z.; Yang, S. Synthesis of novel 18 β-glycyrrhetinic piperazine amides displaying significant in vitro and in vivo antibacterial activities against intractable plant bacterial diseases. Pest Manag. Sci. 2020, 76, 2959–2971. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, C.; Bermúdez, M.Á.; Romero, L.C.; Gotor, C.; García, I. Cysteine homeostasis plays an essential role in plant immunity. New Phytol. 2012, 193, 165–177. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-C.; Holmes, E.C.; Rajniak, J.; Kim, J.-G.; Tang, S.; Fischer, C.R.; Mudgett, M.B.; Sattely, E.S. N-hydroxy-pipecolic acid is a mobile metabolite that induces systemic disease resistance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E4920–E4929. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, M.; Zeier, T.; Bernsdorff, F.; Reichel-Deland, V.; Kim, D.; Hohmann, M.; Scholten, N.; Schuck, S.; Bräutigam, A.; Hölzel, T.; et al. Flavin monooxygenase-generated N-hydroxypipecolic acid is a critical element of plant systemic immunity. Cell 2018, 173, 456–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Liu, R.; Lim, G.-H.; de Lorenzo, L.; Yu, K.; Zhang, K.; Hunt, A.G.; Kachroo, A.; Kachroo, P. Pipecolic acid confers systemic immunity by regulating free radicals. Sci. Adv. 2018, 4, eaar4509. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Brumos, J.; Li, H.; Ji, Y.; Ke, M.; Gong, X.; Zeng, Q.; Li, W.; Zhang, X.; An, F.; et al. A Small-Molecule Screen Identifies l-Kynurenine as a Competitive Inhibitor of TAA1/TAR Activity in Ethylene-Directed Auxin Biosynthesis and Root Growth in Arabidopsis. Plant Cell 2011, 23, 3944–3960. [Google Scholar] [CrossRef] [Green Version]
- Dekker, J. L-methionine induced inhibition of powdery mildew and its reversal by folic acid. Eur. J. Plant Pathol. 1969, 75, 182–185. [Google Scholar] [CrossRef]
- Joshi, J.R.; Yao, L.; Charkowski, A.O.; Heuberger, A.L. Metabolites from Wild Potato Inhibit Virulence Factors of the Soft Rot and Blackleg Pathogen Pectobacterium brasiliense. Mol. Plant Microbe Interact. 2021, 34, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Cowley, T.; Walters, D.R. Polyamine metabolism in barley reacting hypersensitively to the powdery mildew fungus Blumeria graminisf. sp hordei. Plant Cell Environ. 2002, 25, 461–468. [Google Scholar] [CrossRef]
- El Ghachtouli, N.; Martin-Tanguy, J.; Paynot, M.; Gianinazzi, S. First-report of the inhibition of arbuscular mycorrhizal infection of Pisum sativumby specific and irreversible inhibition of polyamine biosynthesis or by gibberellic acid treatment. FEBS Lett. 1996, 385, 189–192. [Google Scholar] [CrossRef]
- Walters, D.; Meurer-Grimes, B.; Rovira, I. Antifungal activity of three spermidine conjugates. FEMS Microbiol. Lett. 2001, 201, 255–258. [Google Scholar] [CrossRef]
- Walters, D.R. Polyamines and plant disease. Phytochemistry 2003, 64, 97–107. [Google Scholar] [CrossRef]
- Lowe-Power, T.M.; Hendrich, C.G.; von Roepenack-Lahaye, E.; Li, B.; Wu, D.; Mitra, R.; Dalsing, B.L.; Ricca, P.; Naidoo, J.; Cook, D.; et al. Metabolomics of tomato xylem sap during bacterial wilt reveals Ralstonia solanacearum produces abundant putrescine, a metabolite that accelerates wilt disease. Environ. Microbiol. 2018, 20, 1330–1349. [Google Scholar] [CrossRef]
- Liu, C.; Atanasov, K.E.; Arafaty, N.; Murillo, E.; Tiburcio, A.F.; Zeier, J.; Alcázar, R. Putrescine elicits ROS -dependent activation of the salicylic acid pathway in Arabidopsis thaliana. Plant Cell Environ. 2020, 43, 2755–2768. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Alcázar, R. A new insight into the contribution of putrescine to defense in Arabidopsis thaliana. Plant Signal. Behav. 2021, 16, 1885187. [Google Scholar] [CrossRef]
- Walters, D.R.; Mackintosh, C.A. Control of plant disease by perturbation of fungal polyamine metabolism. Physiol. Plant. 1997, 100, 689–695. [Google Scholar] [CrossRef]
- Chen, M.; Herde, M.; Witte, C.-P. Of the nine cytidine deaminase like genes in Arabidopsis thaliana eight are pseudogenes and only one is required to maintain pyrimidine homeostasis in vivo. Plant Physiol. 2016, 171, 799–809. [Google Scholar] [CrossRef] [Green Version]
- Martín, S.; Cuevas, J.M.; Grande-Pérez, A.; Elena, S.F. A putative antiviral role of plant cytidine deaminases. F1000Research 2017, 6, 622. [Google Scholar] [CrossRef]
- Ma, X.; Wang, W.-M.; Bittner, F.; Schmidt, N.; Berkey, R.; Zhang, L.; King, H.; Zhang, Y.; Feng, J.; Wen, Y.; et al. Dual and Opposing Roles of Xanthine Dehydrogenase in Defense-Associated Reactive Oxygen Species Metabolism in Arabidopsis. Plant Cell 2016, 28, 1108–1126. [Google Scholar] [CrossRef] [Green Version]
- Stasolla, C.; Loukanina, N.; Yeung, E.C.; Thorpe, T.A. Alterations in pyrimidine nucleotide metabolism as an early signal during the execution of programmed cell death in tobacco BY-2 cells. J. Exp. Bot. 2004, 55, 2513–2522. [Google Scholar] [CrossRef] [PubMed]
- Niles, R.M.; Mount, M.S. Failure to Detect Cyclic 3′, 5′-Adenosine Monophosphate in Healthy and Crown Gall Tumorous Tissues of Vicia faba. Plant Physiol. 1974, 54, 372–373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durner, J.; Wendehenne, D.; Klessig, D.F. Defense gene induction in tobacco by nitric oxide, cyclic GMP, and cyclic ADP-ribose. Proc. Natl. Acad. Sci. USA 1998, 95, 10328–10333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, F.; Zhao, S.; Dong, H.; Zhang, H.; Sun, L.; Miao, C. Nia1 and Nia2 are Involved in Exogenous Salicylic Acid-induced Nitric Oxide Generation and Stomatal Closure in Arabidopsis. J. Integr. Plant Biol. 2010, 52, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Gross, I.; Durner, J. In Search of Enzymes with a Role in 3′, 5′-Cyclic Guanosine Monophosphate Metabolism in Plants. Front. Plant Sci. 2016, 7, 576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, F.; Yang, F.; Rong, W.; Wu, X.; Zhang, J.; Chen, S.; He, C.; Zhou, J.-M. A Xanthomonas uridine 5′-monophosphate transferase inhibits plant immune kinases. Nat. Cell Biol. 2012, 485, 114–118. [Google Scholar] [CrossRef]
- Bent, K.J. Fungitoxic action of dimethirimol and ethirimol. Ann. Appl. Biol. 1970, 66, 103–113. [Google Scholar] [CrossRef]
- Vielba-Fernández, A.; Polonio, Á.; Ruiz-Jiménez, L.; De Vicente, A.; Pérez-García, A.; Fernández-Ortuño, D. Fungicide Resistance in Powdery Mildew Fungi. Microorganisms 2020, 8, 1431. [Google Scholar] [CrossRef] [PubMed]
- Miwa, A.; Sawada, Y.; Tamaoki, D.; Hirai, M.Y.; Kimura, M.; Sato, K.; Nishiuchi, T. Nicotinamide mononucleotide and related metabolites induce disease resistance against fungal phytopathogens in Arabidopsis and barley. Sci. Rep. 2017, 7, 6389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, B.; Li, X.; Ouyang, Z.; Huang, L.; Hong, Y.; Zhang, H.; Li, D.; Song, F. The de novo Biosynthesis of Vitamin B6 Is Required for Disease Resistance Against Botrytis cinerea in Tomato. Mol. Plant Microbe Interact. 2014, 27, 688–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samsatly, J.; Bayen, S.; Jabaji, S.H. Vitamin B6 Is Under a Tight Balance During Disease Development by Rhizoctonia solani on Different Cultivars of Potato and on Arabidopsis thaliana Mutants. Front. Plant Sci. 2020, 11, 875. [Google Scholar] [CrossRef]
- Tintor, N.; Paauw, M.; Rep, M.; Takken, F.L.W. The root-invading pathogen Fusarium oxysporum targets pattern-triggered immunity using both cytoplasmic and apoplastic effectors. New Phytol. 2020, 227, 1479–1492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, P.; Meng, Y.; Wise, R.P. Functional Contribution of Chorismate Synthase, Anthranilate Synthase, and Chorismate Mutase to Penetration Resistance in Barley–Powdery Mildew Interactions. Mol. Plant Microbe Interact. 2009, 22, 311–320. [Google Scholar] [CrossRef] [Green Version]
- Chripkova, M.; Drutovic, D.; Pilatova, M.; Mikes, J.; Budovska, M.; Vaskova, J.; Broggini, M.; Mirossay, L.; Mojzis, J. Brassinin and its derivatives as potential anticancer agents. Toxicol. Vitr. 2014, 28, 909–915. [Google Scholar] [CrossRef]
- Klein, A.P.; Sattely, E.S. Biosynthesis of cabbage phytoalexins from indole glucosinolate. Proc. Natl. Acad. Sci. USA 2017, 114, 1910–1915. [Google Scholar] [CrossRef] [Green Version]
- Pedras, M.S.C.; Okinyo, D.P.O. En route to erucalexin: A unique rearrangement in the crucifer phytoalexin biosynthetic pathway. Chem. Commun. 2006, 17, 1848–1850. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Okinyo, D.P.O. Syntheses of perdeuterated indoles and derivatives as probes for the biosyntheses of crucifer phytoalexins. J. Label. Compd. Radiopharm. 2006, 49, 33–45. [Google Scholar] [CrossRef]
- Ahuja, I.; Kissen, R.; Bones, A.M. Phytoalexins in defense against pathogens. Trends Plant Sci. 2012, 17, 73–90. [Google Scholar] [CrossRef]
- Munhoz, C.D.F.; Santos, A.; Arenhart, R.; Santini, L.; Monteiro-Vitorello, C.B.; Vieira, M.L.C. Analysis of plant gene expression during passion fruit- Xanthomonas axonopodis interaction implicates lipoxygenase 2 in host defence. Ann. Appl. Biol. 2015, 167, 135–155. [Google Scholar] [CrossRef]
- Araújo, E.R.; Costa, J.R.; Pontes, N.C.; Quezado-Duval, A.M. Xanthomonas perforans and X. gardneri associated with bacterial leaf spot on weeds in Brazilian tomato fields. Eur. J. Plant Pathol. 2015, 143, 543–548. [Google Scholar] [CrossRef]
- Santos, L.V.S.; Melo, E.A.; Silva, A.M.F.; Félix, K.C.S.; Quezado-Duval, A.M.; Albuquerque, G.M.R.; Gama, M.A.S.; Souza, E.B. Weeds as alternate hosts of Xanthomonas euvesicatoria pv euvesicatoria and X. campestris pv. campestris in vegetable-growing fields in the state of Pernambuco, Brazil. Trop. Plant Pathol. 2020, 45, 1–9. [Google Scholar] [CrossRef]
- Tamura, K.; Takikawa, Y.; Tsuyumu, S.; Goto, M.; Watanabe, M. Coronatine Production by Xanthomonas campestris pv phormiicola. Jpn. J. Phytopathol. 1992, 58, 276–281. [Google Scholar] [CrossRef] [Green Version]
- Szabó, T.; Volk, B.; Milen, M. Recent Advances in the Synthesis of β-Carboline Alkaloids. Molecules 2021, 26, 663. [Google Scholar] [CrossRef]
- Piechowska, P.; Zawirska-Wojtasiak, R.; Mildner-Szkudlarz, S. Bioactive β-Carbolines in Food: A Review. Nutrients 2019, 11, 814. [Google Scholar] [CrossRef] [Green Version]
- Spies, C.D.; Rommelspacher, H.; Schnapper, C.; Muller, C.; Marks, C.; Berger, G.; Conrad, C.; Blum, S.; Specht, M.; Hannemann, L.; et al. beta-Carbolines in Chronic Alcoholics Undergoing Elective Tumor Resection. Alcohol. Clin. Exp. Res. 1995, 19, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Rommelspacher, H.; Dufeu, P.; Schmidt, L.G. Harman and Norharman in Alcoholism: Correlations with Psychopathology and Long-Term Changes. Alcohol. Clin. Exp. Res. 1996, 20, 3–8. [Google Scholar] [CrossRef]
- Pfau, W.; Skog, K. Exposure to β-carbolines norharman and harman. J. Chromatogr. B 2004, 802, 115–126. [Google Scholar] [CrossRef]
- Bibi, F. Diversity of antagonistic bacteria isolated from medicinal plant Peganum harmala L. Saudi J. Biol. Sci. 2017, 24, 1288–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cain, C.C.; Lee, D.; Waldo, R.H.; Henry, A.T.; Casida, E.J.; Wani, M.C.; Wall, M.E.; Oberlies, N.H.; Falkinham, J.O. Synergistic Antimicrobial Activity of Metabolites Produced by a Nonobligate Bacterial Predator. Antimicrob. Agents Chemother. 2003, 47, 2113–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puopolo, G.; Cimmino, A.; Palmieri, M.; Giovannini, O.; Evidente, A.; Pertot, I. Lysobacter capsici AZ78 produces cyclo(l -Pro-l -Tyr), a 2,5-diketopiperazine with toxic activity against sporangia of Phytophthora infestans and Plasmopara viticola. J. Appl. Microbiol. 2014, 117, 1168–1180. [Google Scholar] [CrossRef]
- Wattana-Amorn, P.; Charoenwongsa, W.; Williams, C.; Crump, M.; Apichaisataienchote, B. Antibacterial activity of cyclo(L-Pro-L-Tyr) and cyclo(D-Pro-L-Tyr) from Streptomyces sp. strain 22-4 against phytopathogenic bacteria. Nat. Prod. Res. 2016, 30, 1980–1983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Wang, X.; Wang, M.; Chang, Y.; Zhang, F.; Ban, Z.; Tang, R.; Gan, Q.; Wu, S.; Guo, Y.; et al. The lysine catabolite saccharopine impairs development by disrupting mitochondrial homeostasis. J. Biol. 2019, 218, 580–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, P.F.; Constantine, J.W.; Barth, W.E. Pirbuterol, a selective beta-2 adrenergic bronchodilator. J. Pharmacol. Exp. Ther. 1978, 207, 410. [Google Scholar] [PubMed]
- Richards, D.M.; Brogden, R.N. Pirbuterol A Preliminary Review of its Pharmacological Properties and Therapeutic Efficacy in Reversible Bronchospastic Disease. Drugs 1985, 30, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Kittaka, H.; Yamanoi, Y.; Tominaga, M. Transient receptor potential vanilloid 4 (TRPV4) channel as a target of crotamiton and its bimodal effects. Pflüger’s Archiv Gesamte Physiol. Menschen Tiere 2017, 469, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Corsini, E.; Dell’Agli, M.; Facchi, A.; De Fabiani, E.; Lucchi, L.; Boraso, M.S.; Marinovich, M.; Galli, C.L. Enterodiol and Enterolactone Modulate the Immune Response by Acting on Nuclear Factor-κB (NF-κB) Signaling. J. Agric. Food Chem. 2010, 58, 6678–6684. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.E.; Riddick, E.W.; Aldrich, J.R.; Holmes, W.E. Identification of (−)-β-Caryophyllene as a Gender-Specific Terpene Produced by the Multicolored Asian Lady Beetle. J. Chem. Ecol. 2006, 32, 2489–2499. [Google Scholar] [CrossRef]
- Kollner, T.G.; Held, M.; Lenk, C.; Hiltpold, I.; Turlings, T.C.; Gershenzon, J.; Degenhardt, J. A Maize (E)-β-Caryophyllene Synthase Implicated in Indirect Defense Responses against Herbivores Is Not Expressed in Most American Maize Varieties. Plant Cell 2008, 20, 482–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snelders, N.C.; Petti, G.C.; Berg, G.C.M.V.D.; Seidl, M.F.; Thomma, B.P.H.J. An ancient antimicrobial protein co-opted by a fungal plant pathogen for in planta mycobiome manipulation. Proc. Natl. Acad. Sci. USA 2021, 118, 2110968118. [Google Scholar] [CrossRef] [PubMed]
- Jakubiec-Krzesniak, K.; Rajnisz-Mateusiak, A.; Guspiel, A.; Ziemska, J.; Solecka, J. Secondary Metabolites of Actinomycetes and their Antibacterial, Antifungal and Antiviral Properties. Pol. J. Microbiol. 2018, 67, 259–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabral, D.; Penumutchu, S.; Reinhart, E.M.; Zhang, C.; Korry, B.J.; Wurster, J.I.; Nilson, R.; Guang, A.; Sano, W.H.; Rowan-Nash, A.D.; et al. Microbial Metabolism Modulates Antibiotic Susceptibility within the Murine Gut Microbiome. Cell Metab. 2019, 30, 800–823. [Google Scholar] [CrossRef]
- Zou, L.; Hu, Y.-Y.; Chen, W.-X. Antibacterial mechanism and activities of black pepper chloroform extract. J. Food Sci. Technol. 2015, 52, 8196–8203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boriollo, M.F.G.; Marques, M.B.; Da Silva, T.A.; Da Silva, J.J.; Dias, R.A.; Filho, T.H.N.S.; Melo, I.L.R.; Dias, C.T.D.S.; Bernardo, W.L.D.C.; Oliveira, N.D.M.S.; et al. Antimicrobial potential, phytochemical profile, cytotoxic and genotoxic screening of Sedum praealtum A. DC. (balsam). BMC Complement. Med. Ther. 2020, 20, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wink, M. Evolution of secondary metabolites from an ecological and molecular phylogenetic perspective. Phytochemistry 2003, 64, 3–19. [Google Scholar] [CrossRef]
- Jones, J.B.; Lacy, G.H.; Bouzar, H.; Stall, R.E.; Schaad, N.W. Reclassification of the Xanthomonads Associated with Bacterial Spot Disease of Tomato and Pepper. Syst. Appl. Microbiol. 2004, 27, 755–762. [Google Scholar] [CrossRef] [Green Version]
- Bophela, K.N.; Venter, S.N.; Wingfield, M.J.; Duran, A.; Tarigan, M.; Coutinho, T.A. Xanthomonas perforans: A tomato and pepper pathogen associated with bacterial blight and dieback of Eucalyptus pellita seedlings in Indonesia. Australas. Plant Pathol. 2019, 48, 543–551. [Google Scholar] [CrossRef]
- Sbaghi, M.; Jeandet, P.; Bessis, R.; Leroux, P. Degradation of stilbene-type phytoalexins in relation to the pathogenicity of Botrytis cinerea to grapevines. Plant Pathol. 1996, 45, 139–144. [Google Scholar] [CrossRef]
- Jeandet, P.; Hébrard, C.; Deville, M.-A.; Cordelier, S.; Dorey, S.; Aziz, A.; Crouzet, J. Deciphering the Role of Phytoalexins in Plant-Microorganism Interactions and Human Health. Molecules 2014, 19, 18033–18056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raasch-Fernandes, L.D.; Bonaldo, S.M.; Rodrigues, D.; Junior, G.M.V.; Schwan-Estrada, K.R.F.; Da Silva, C.R.; Verçosa, A.G.A.; De Oliveira, D.L.; DeBiasi, B.W. Induction of phytoalexins and proteins related to pathogenesis in plants treated with extracts of cutaneous secretions of southern Amazonian Bufonidae amphibians. PLoS ONE 2019, 14, e0211020. [Google Scholar] [CrossRef] [PubMed]
- Ultee, A.; Smid, E. Influence of carvacrol on growth and toxin production by Bacillus cereus. Int. J. Food Microbiol. 2001, 64, 373–378. [Google Scholar] [CrossRef]
- Xu, J.; Zhou, F.; Ji, B.-P.; Pei, R.-S.; Xu, N. The antibacterial mechanism of carvacrol and thymol against Escherichia coli. Lett. Appl. Microbiol. 2008, 47, 174–179. [Google Scholar] [CrossRef]
- Churklam, W.; Chaturongakul, S.; Ngamwongsatit, B.; Aunpad, R. The mechanisms of action of carvacrol and its synergism with nisin against Listeria monocytogenes on sliced bologna sausage. Food Control. 2020, 108, 106864. [Google Scholar] [CrossRef]
- Alagawany, M.; Farag, M.R.; Abdelnour, S.A.; ElNesr, S.S. A review on the beneficial effect of thymol on health and production of fish. Rev. Aquac. 2021, 13, 632–641. [Google Scholar] [CrossRef]
- Joshi, J.R.; Khazanov, N.; Senderowitz, H.; Burdman, S.; Lipsky, A.; Yedidia, I. Plant phenolic volatiles inhibit quorum sensing in pectobacteria and reduce their virulence by potential binding to ExpI and ExpR proteins. Sci. Rep. 2016, 6, 38126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, J.R.; Khazanov, N.; Khadka, N.; Charkowski, A.O.; Burdman, S.; Carmi, N.; Yedidia, I.; Senderowitz, H. Direct Binding of Salicylic Acid to Pectobacterium N-Acyl-Homoserine Lactone Synthase. ACS Chem. Biol. 2020, 15, 1883–1891. [Google Scholar] [CrossRef]
- Luo, H.-Z.; Guan, Y.; Yang, R.; Qian, G.-L.; Yang, X.-H.; Wang, J.-S.; Jia, A.-Q. Growth inhibition and metabolomic analysis of Xanthomonas oryzae pv oryzae treated with resveratrol. BMC Microbiol. 2020, 20, 1–13. [Google Scholar] [CrossRef]
- Radapong, S.; Sarker, S.D.; Ritchie, K.J. Oxyresveratrol Possesses DNA Damaging Activity. Molecules 2020, 25, 2577. [Google Scholar] [CrossRef]
- Horvathova, E.; Navarova, J.; Galova, E.; Sevcovicova, A.; Chodakova, L.; Snahnicanova, Z.; Melusova, M.; Kozics, K.; Slamenova, D. Assessment of Antioxidative, Chelating, and DNA-Protective Effects of Selected Essential Oil Components (Eugenol, Carvacrol, Thymol, Borneol, Eucalyptol) of Plants and Intact Rosmarinus officinalis Oil. J. Agric. Food Chem. 2014, 62, 6632–6639. [Google Scholar] [CrossRef] [PubMed]
- Aristatile, B.; Al-Numair, K.S.; Al-Assaf, A.H.; Pugalendi, K.V. Pharmacological effect of carvacrol on d-galactosamine-induced mitochondrial enzymes and DNA damage by single-cell gel electrophoresis. J. Nat. Med. 2011, 65, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Melusova, M.; Slamenova, D.; Kozics, K.; Jantova, S.; Horvathova, E. Carvacrol and rosemary essential oil manifest cytotoxic, DNA-protective and pro-apoptotic effect having no effect on DNA repair. Neoplasma 2014, 61, 690–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Ploeg, J.R.; Weiss, M.A.; Saller, E.; Nashimoto, H.; Saito, N.; Kertesz, M.A.; Leisinger, T. Identification of sulfate starvation-regulated genes in Escherichia coli: A gene cluster involved in the utilization of taurine as a sulfur source. J. Bacteriol. 1996, 178, 5438–5446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eichhorn, E.; van der Ploeg, J.R.; Kertesz, M.; Leisinger, T. Characterization of α-Ketoglutarate-dependent Taurine Dioxygenase from Escherichia coli. J. Biol. Chem. 1997, 272, 23031–23036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borchert, A.J.; Gouveia, G.J.; Edison, A.S.; Downs, D.M. Proton Nuclear Magnetic Resonance Metabolomics Corroborates Serine Hydroxymethyltransferase as the Primary Target of 2-Aminoacrylate in a ridA Mutant of Salmonella enterica. mSystems 2020, 5, e00843-19. [Google Scholar] [CrossRef] [Green Version]
- Borchert, A.J.; Walejko, J.M.; Le Guennec, A.; Ernst, D.C.; Edison, A.S.; Downs, D.M. Integrated Metabolomics and Transcriptomics Suggest the Global Metabolic Response to 2-Aminoacrylate Stress in Salmonella enterica. Metabolities 2019, 10, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jibrin, M.O.; Liu, Q.; Jones, J.B.; Zhang, S. Surfactants in plant disease management: A brief review and case studies. Plant Pathol. 2021, 70, 495–510. [Google Scholar] [CrossRef]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [Green Version]
- Goodacre, R.; Broadhurst, D.; Smilde, A.K.; Kristal, B.S.; Baker, J.D.; Beger, R.; Bessant, C.; Connor, S.; Capuani, G.; Craig, A.; et al. Proposed minimum reporting standards for data analysis in metabolomics. Metabolomics 2007, 3, 231–241. [Google Scholar] [CrossRef]
- Salek, R.M.; Haug, K.; Steinbeck, C. Dissemination of metabolomics results: Role of MetaboLights and COSMOS. GigaScience 2013, 2, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, J.; Wishart, D.S.; Xia, J. Using MetaboAnalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef] [PubMed]
- Karp, P.D.; Billington, R.; Caspi, R.; Fulcher, C.A.; Latendresse, M.; Kothari, A.; Keseler, I.M.; Krummenacker, M.; Midford, P.E.; Ong, Q.; et al. The BioCyc collection of microbial genomes and metabolic pathways. Briefings Bioinform. 2019, 20, 1085–1093. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Set | Significant Metabolites (p-Value < 0.05) | ||
|---|---|---|---|
| Identified Metabolites 1 | Unidentified Metabolites | Total Metabolites | |
| Positive Mode | 81 | 559 | 620 |
| Negative Mode | 86 | 353 | 439 |
| Total | 167 2 | 937 | 1059 |
| X. perforans Metabolites | Plant or Microbial Systems Where Metabolite Was Reported with References | Known Xanthomonas Pathogen of Plants with Metabolite with References | This Study as First Report in a Microbe (M) or Xanthomonas Species (X) |
|---|---|---|---|
| Methoxybrassinin A (M/Z = 281.0764; Rt = 10.70 min) 2 | Brassicas [55] | Xanthomonas campestris pv. campestris on crucifers [56] | M |
| Cyclobrassinone (M/Z = 231.0245; Rt = 0.76 min) | Brassicas [57,58] | Xanthomonas campestris pv. campestris on crucifers [56] | M |
| Sarmentosin (M/Z = 276.1071; Rt = 3.26 min.) | Plants in the Passiflora genus and their heliconiine butterfly pests 9 [59,60] | X. axonopodis pv. passiflorae on Passiflora species [61] | M |
| (2s,4s)-monatin (M/Z = 293.1127; Rt = 7.66 min) | Sclerochiton ilicifolius [62,63] | None reported | M |
| Piperidine (M/Z = 86.0964; Rt = 1.07 min) | Piper nigrum [64,65] | X. campestris pv. betlicola on P. nigrum [66] | X |
| Enterodiol sulfate (M/Z = 383.1149; Rt = 0.73 min) | Flax seed [67] | None reported | X |
| β-Carboline (M/Z = 169.0759; Rt = 8.54 min)Harman (M/Z = 183.0913; Rt = 8.80 min)Norharman (M/Z = 157.1082; Rt = 1.35 min) | Many plants, including passion fruits, Peganum harmala, Picrasma quassioides [68,69,70] | X. axonopodis pv. passiflorae on Passiflora species [61] | X |
| Pyrocoll (M/Z = 187.0494; Rt = 6.30 min) | Smoke, Streptomyces [71] | X | |
| Maculosin (M/Z = 261.1224; Rt = 7.69 min) | Alternaria alternata [72] | X | |
| Saccharopine (M/Z = 275.1260; Rt = 0.76 min) | Saccharomyces cerevisiae [73] | X |
| Class, Metabolites | Disease System | References |
|---|---|---|
| Alkaloids | ||
| β-Carboline (M/Z = 169.0759; Rt = 8.54 min) 1 | Rice Bacterial Blight, Kiwifruit Bacterial Canker, and Citrus Bacterial Canker | [74] |
| Piperidine (M/Z = 860964; Rt = 1.07 min) | Xanthomonas oryzae pv. oryzae and X. axonopodis pv. citri | [75,76] |
| Amines | ||
| Amino Acids | ||
| L-cysteine (M/Z = 122.0266; Rt = 0.76 min) | Pseudomonas syringae pv. tomato on Arabidopsis | [77] |
| Pipecolate/L-pipecolic ACID (M/Z = 130.0861; Rt = 1.25 min) | Systemic resistance in Arabidopsis, against bacterial plant pathogens | [78,79,80] |
| L-kynurenine (M/Z = 209.0915; Rt = 6.43 min) | Inhibit ethylene responses in Arabidopsis to disease resistance | [81] |
| L-methionine (M/Z = 150.0581; Rt = 1.43 min) | Powdery mildew on cucumber | [82] |
| Proline (M/Z = 114.0562; Rt = 0.85 min) | Pectobacterium brasiliense | [83] |
| Polyamines | ||
| Spermidine (M/Z = 146.1649; Rt = 0.56 min) | Blumeria graminis f.sp. hordei, Pyrenophora avenae | [84,85,86,87] |
| Putrescine (M/Z = 89.1073; Rt = 0.60 min) | Arabidopsis; virulence factor in Ralstonia solanacearum | [88,89,90] |
| N-acetylputrescine (M/Z = 131.1176; Rt = 1.13 min) | [91] | |
| N-Carbamoylputrescine (M/Z = 132.1130; Rt = 1.47 min) | ||
| Nucleotides/Nucleosides | ||
| Cytidine (M/Z = 244.0922; Rt = 1.66 min) | Cauliflower mosaic virus; | [92,93] |
| Hypoxanthine (M/Z = 137.0456; Rt = 2.11 min) | Powdery mildew in Arabidopsis | [94] |
| Uracil (M/Z = 113.0344; Rt = 1.44 min) | Programmed cell death | [95] |
| Adenosine-5′-Monophosphate (M/Z = 348.0694; Rt = 1.70 min) | Crown gall of Vicia faba | [96] |
| Guanosine-5-Monophosphate (M/Z = 364.0642; Rt = 2.01 min) | Arabidopsis, Tobacco | [97,98,99] |
| Uridine-5-Monophosphate (M/Z = 325.0421; Rt = 1.25 min) | Xanthomonas black rot of crucifers | [100] |
| Dimethirimol (M/Z = 210.1597; Rt = 7.12 min) | Fungicide | [101,102] |
| Vitamins | ||
| Nicotinamide (M/Z = 123.0551; Rt = 1.70 (Vitamin B3) | Fusarium sp. On barley and Arabidopsis | [103] |
| Vitamin B6 | Botrytis cinerea on tomato; Rhizoctonia solani on Potato and Arabidopsis | [104,105] |
| Pyridoxal (M/Z = 168.0649; Rt = 1.94 min (Vitamin B6) | ||
| Pyridoxine (M/Z = 170.0804; Rt = 2.93 min (Vitamin B6) | ||
| Pyridoxine (M/Z = 192.0627; Rt = 2.88 min (Vitamin B6) | ||
| Biotin (M/Z = 245.0946; Rt = 6.58 min) | Fusarium oxysporium in Arabidopsis | [106] |
| Organic Acids | ||
| Anthranilate (M/Z = 120.0443; Rt = 8.74 min) | Powdery mildew on barley; | [107] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jibrin, M.O.; Liu, Q.; Guingab-Cagmat, J.; Jones, J.B.; Garrett, T.J.; Zhang, S. Metabolomics Insights into Chemical Convergence in Xanthomonas perforans and Metabolic Changes Following Treatment with the Small Molecule Carvacrol. Metabolites 2021, 11, 879. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120879
Jibrin MO, Liu Q, Guingab-Cagmat J, Jones JB, Garrett TJ, Zhang S. Metabolomics Insights into Chemical Convergence in Xanthomonas perforans and Metabolic Changes Following Treatment with the Small Molecule Carvacrol. Metabolites. 2021; 11(12):879. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120879
Chicago/Turabian StyleJibrin, Mustafa Ojonuba, Qingchun Liu, Joy Guingab-Cagmat, Jeffrey B. Jones, Timothy J. Garrett, and Shouan Zhang. 2021. "Metabolomics Insights into Chemical Convergence in Xanthomonas perforans and Metabolic Changes Following Treatment with the Small Molecule Carvacrol" Metabolites 11, no. 12: 879. https://0-doi-org.brum.beds.ac.uk/10.3390/metabo11120879