
Retrospective Proteomic Analysis of Cellular Immune Responses and Protective Correlates of p24 Vaccination in an HIV Elite Controller Using Antibody Arrays


Abstract
:1. Introduction
2. Methods and Materials
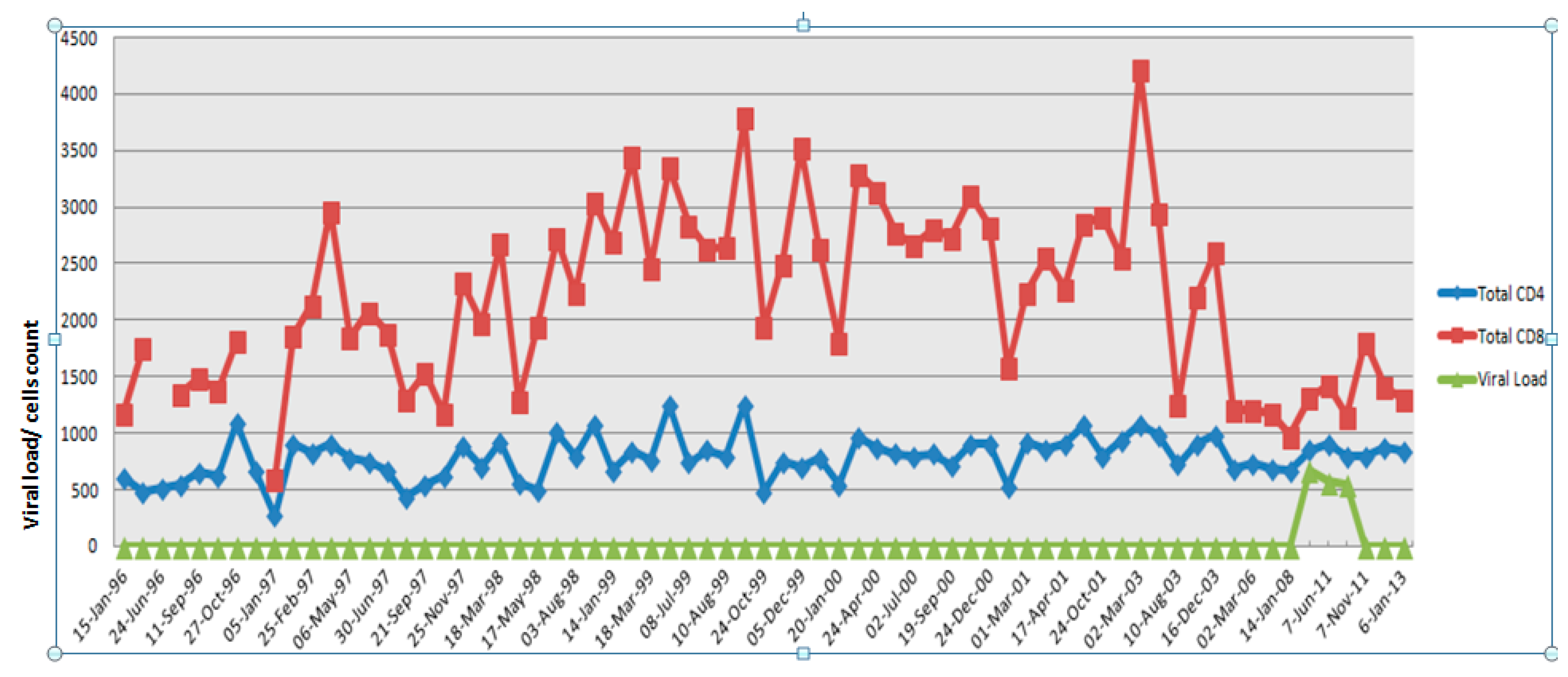
2.1. Clinical History of the Study Subject
2.2. PBMCs Separation and Sample Preparation
2.3. Macs Column Separation of CD4+, CD8+ T Cells and Monocytes
2.4. Protein Extraction
2.5. Quantification Using the BioRad DC-Protein Assay
2.6. RayBio® Combination of Human Cytokine Antibody Array G Series
2.7. Data Normalization and Analysis
2.8. Statistical Analysis Using K-Way and Log Linear Models to Discover K–Gene Interactions
2.9. Follow-up Tests
3. Results
3.1. Overlapping Proteins that Functionally Bind CD4+, CD8+ T Cells,and CD14+Monocytes: Evidence for Crosstalk between Cell Types
3.2. HIV Aviremic Phase
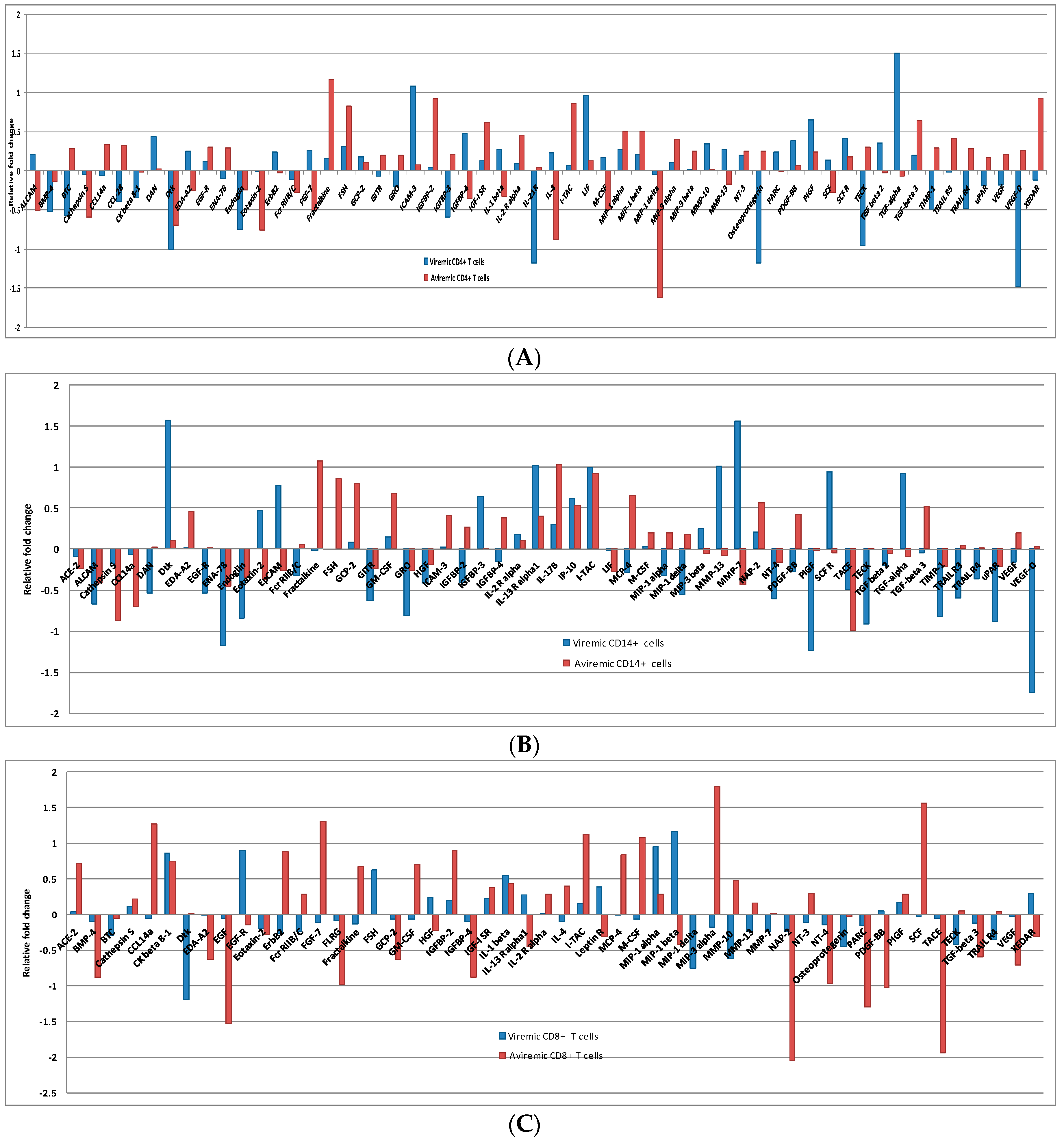
3.3. Cell-Specific Differentially Expressed Proteins during Viremic and Aviremic Phases
3.3.1. Proteins Segregating Viremic and Aviremic Phases Based on CD4+ T Cells
3.3.2. Proteins Segregating Viremic and Aviremic Phases Based on CD14+ Monocytes
3.3.3. Proteins Segregating Viremic and Aviremic Phases Based on CD8+ T Cells
3.4. CD4+ T Cells and Monocytes Are Key Players during Viremia, Whereas CD4+, CD8+ T Cells and Monocytes Are Equal Partners during the Aviremic Phase
3.5. Q-RTPCR Validation of Proteomic Expression: High Concordance between Protein and Genomic Expression Trends
3.6. K-Way and Log-Linear Model Analysis Reveal Joint Expression of Genes from All Three Cell Types Significant in Two- and Three-Way Interactions
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dyer, W.B.; Zaunders, J.J.; Yuan, F.F.; Wang, B.; Learmont, J.C.; Geczy, A.F.; Saksena, N.K.; McPhee, D.A.; Gorry, P.R.; Sullivan, J.S. Mechanisms of HIV non-progression; robust and sustained CD4+ T-cell proliferative responses to p24 antigen correlate with control of viremia and lack of disease progression after long-term transfusion-acquired HIV-1 infection. Retrovirology 2008, 5. [Google Scholar] [CrossRef] [PubMed]
- Pontesilli, O.; Carotenuto, P.; Kerkhof-Garde, S.R.; Roos, M.T.; Keet, I.P.; Coutinho, R.A.; Goudsmit, J.; Miedema, F. Lymphoproliferative response to HIV type 1 p24 in long-term survivors of HIV type 1 infection is predictive of persistent AIDS-free infection. AIDS Res. Hum. Retroviruses 1999, 15, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Kiepiela, P.; Ngumbela, K.; Thobakgale, C.; Ramduth, D.; Honeyborne, I.; Moodley, E.; Reddy, S.; de Pierres, C.; Mncube, Z.; Mkhwanazi, N.; et al. CD8+ T-cell responses to different HIV proteins have discordant associations with viral load. Nat. Med. 2007, 13, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, F.O.; Tasken, K.; Kvale, D. Combined Env- and Gag-specific T cell responses in relation to PD-1 and CD4+ T cell loss rates in HIV-1 infection. Clin. Exp. Immunol. 2010, 161, 315–323. [Google Scholar] [PubMed]
- Rolland, M.; Heckerman, D.; Deng, W.; Rousseau, C.M.; Coovadia, H.; Bishop, K.; Goulder, P.J.; Walker, B.D.; Brander, C.; Mullins, J.I. Broad and Gag-biased HIV-1 epitope repertoires are associated with lower viral loads. PLoS ONE 2008, 3, e1424. [Google Scholar] [CrossRef] [PubMed]
- Zuniga, R.; Lucchetti, A.; Galvan, P.; Sanchez, S.; Sanchez, C.; Hernandez, A.; Sanchez, H.; Frahm, N.; Linde, C.H.; Hewitt, H.S.; et al. Relative dominance of Gag p24-specific cytotoxic T lymphocytes is associated with human immunodeficiency virus control. J. Virol. 2006, 80, 3122–3125. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Gow, I.; Colebunders, R.; Weller, I.; Tchamouroff, S.; Weber, J.; Boag, F.; Hales, G.; Adams, S.; Patou, G.; et al. Therapeutic vaccination (p24-VLP) of patients with advanced HIV-1 infection in the pre-HAART era does not alter CD4 cell decline. HIV Med. 2001, 2, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Altfeld, M.; Addo, M.M.; Kreuzer, K.A.; Rockstroh, J.K.; Dumoulin, F.L.; Schliefer, K.; Leifeld, L.; Sauerbruch, T.; Spengler, U. TH1 to TH2 shift of cytokines in peripheral blood of HIV-infected patients is detectable by reverse transcriptase polymerase chain reaction but not by enzyme-linked immunosorbent assay under nonstimulated conditions. J. Acquir. Immune Defic. Syndr. 2000, 23, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Babu, C.K.; Suwansrinon, K.; Bren, G.D.; Badley, A.D.; Rizza, S.A. HIV Induces TRAIL Sensitivity in Hepatocytes. PLoS ONE 2009, 4. [Google Scholar] [CrossRef] [PubMed]
- Barat, C.; Gervais, P.; Tremblay, M.J. Engagement of ICAM-3 provides a costimulatory signal for human immunodeficiency virus type 1 replication in both activated and quiescent CD4+ T lymphocytes: Implications for virus pathogenesis. J. Virol. 2004, 78, 6692–6697. [Google Scholar] [CrossRef] [PubMed]
- Casartelli, N.; Di Matteo, G.; Potesta, M.; Rossi, P.; Doria, M. CD4 and major histocompatibility complex class I down-regulation by the human immunodeficiency virus type 1 Nef protein in pediatric AIDS progression. J. Virol. 2003, 77, 11536–11545. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Dyer, W.B.; Zaunders, J.J.; Mikhail, M.; Sullivan, J.S.; Williams, L.; Haddad, D.N.; Harris, G.; Holt, J.A.; Cooper, D.A.; et al. Comprehensive analyses of a unique HIV-1-infected nonprogressor reveal a complex association of immunobiological mechanisms in the context of replication-incompetent infection. Virology 2002, 304, 246–264. [Google Scholar] [CrossRef] [PubMed]
- Zaunders, J.J.; Dyer, W.B.; Wang, B.; Munier, M.L.; Miranda-Saksena, M.; Newton, R.; Moore, J.; Mackay, C.R.; Cooper, D.A.; Saksena, N.K.; et al. Identification of circulating antigen-specific CD4+ T lymphocytes with a CCR5+, cytotoxic phenotype in an HIV-1 long-term nonprogressor and in CMV infection. Blood 2004, 103, 2238–2247. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, D.; Hertzog, P.; Tomkinson, E.; Couldwell, D.; McCarville, S.; Parrish, S.; Cunningham, P.; Newell, M.; Owens, M.; Cooper, D. Aadministration of imiquimod, an interferon inducer, in asymptomatic human immunodeficiency virus-infected persons to determine safety and biologic response modification. J. Infect. Dis. 1998, 178, 858–861. [Google Scholar] [CrossRef] [PubMed]
- Kelleher, A.D.; Roggensack, M.; Jaramillo, A.B.; Smith, D.E.; Walker, A.; Gow, I.; McMurchie, M.; Harris, J.; Patou, G.; Cooper, D.A. Safety and immunogenicity of a candidate therapeutic vaccine, p24 virus-like particle, combined with zidovudine, in asymptomatic subjects. Community HIV Research Network Investigators. AIDS 1998, 12, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.J.; Lemey, P.; Dyer, W.B.; Sullivan, J.S.; Chew, C.B.; Vandamme, A.M.; Dwyer, D.E.; Saksena, N.K. Genetic analyses reveal structured HIV-1 populations in serially sampled T lymphocytes of patients receiving HAART. Virology 2006, 348, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Howell, D.C. Statistical Methods for Psychology, 7th ed.; Cengage Learning: Belmot, CA, USA, 2009; pp. 630–655. [Google Scholar]
- Field, A. Discovering Statistics Using SPSS, 2nd ed.; Sage Publications: Thousand Oaks, CA, USA, 2005; pp. 695–718. [Google Scholar]
- Agresti, A. An Introduction to Categorical Data Analysis, 2nd ed.; Wiley Inter-Science: Hoboken, NJ, USA, 2007; p. 212. [Google Scholar]
- Poropatich, K.; Sullivan, D.J., Jr. Human immunodeficiency virus type 1 long-term nonprogressors: The viral, genetic and immunological basis for disease non-progression. J. General. Virol. 2011, 92, 247–268. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Hieshima, K.; Haskell, C.; Baba, M.; Nagira, M.; Nishimura, M.; Kakizaki, M.; Takagi, S.; Nomiyama, H.; Schall, T.J.; et al. Identification and molecular characterization of fractalkine receptor CX3CR1, which mediates both leukocyte migration and adhesion. Cell 1997, 91, 521–530. [Google Scholar] [CrossRef]
- Imami, N.; Pires, A.; Hardy, G.; Wilson, J.; Gazzard, B.; Gotch, F. A balanced type 1/type 2 response is associated with long-term nonprogressive human immunodeficiency virus type 1 infection. J. Virol. 2002, 76, 9011–9023. [Google Scholar] [CrossRef] [PubMed]
- White, G.E.; Greaves, D.R. Fractalkine: A survivor’s guide: Chemokinesas antiapoptotic mediators. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Cotter, R.; Williams, C.; Ryan, L.; Erichsen, D.; Lopez, A.; Peng, H.; Zheng, J. Fractalkine (CX3CL1) and brain inflammation: Implications for HIV-1-associated dementia. J. Neurovirol. 2002, 8, 585–598. [Google Scholar] [CrossRef] [PubMed]
- Landsman, L.; Bar-On, L.; Zernecke, A.; Kim, K.W.; Krauthgamer, R.; Shagdarsuren, E.; Lira, S.A.; Weissman, I.L.; Weber, C.; Jung, S. CX3CR1 is required for monocyte homeostasis and atherogenesis by promoting cell survival. Blood 2009, 113, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Karlmark, K.R.; Zimmermann, H.W.; Roderburg, C.; Gassler, N.; Wasmuth, H.E.; Luedde, T.; Trautwein, C.; Tacke, F. The fractalkine receptor CXCR1 protects against liver fibrosis by controlling differentiation and survival of infiltrating hepatic monocytes. Hepatology 2010, 52, 1769–1782. [Google Scholar] [CrossRef] [PubMed]
- Geijtenbeek, T.B.; van Kooyk, Y. DC-SIGN: A novel HIV receptor on DCs that mediates HIV-1 transmission. Curr. Top. Microbiol. Immunol. 2003, 276, 31–54. [Google Scholar] [PubMed]
- Gallo, R.C.; Garzino-Demo, A.; DeVico, A.L. HIV infection and pathogenesis: What about chemokines? J. Clin. Immunol. 1999, 19, 293–299. [Google Scholar] [CrossRef]
- Mikhail, M.; Bin, W.; Saksena, N.K. Mechanisms involved in non-progressive HIV disease. AIDS Rev. 2003, 3, 230–230. [Google Scholar]
- Kornbluth, R.S.; Kee, K.; Richman, D.D. CD40 Ligand (CD154) stimulation of macrophages to produce HIV-1-suppressive β-chemokines. Proc. Natl. Acad. Sci. USA 1998, 95, 5205–5210. [Google Scholar] [CrossRef] [PubMed]
- Hieshima, K.; Imai, T.; Opdenakker, G.; van Damme, J.; Kusuda, J.; Tei, H.; Sakaki, Y.; Takatsuki, K.; Miura, R.; Yoshie, O.; et al. Molecular cloning of a novel human CC chemokine liver and activation-regulated chemokine (LARC) expressed in liver. Chemotactic activity for lymphocytes and gene localization on chromosome 2. J. Biol. Chem. 1997, 272, 5846–5853. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, T.; Kuwabara, T.; Nakano, H.; Aritomi, K.; Onodera, T.; Lipp, M.; Takahama, Y.; Kakiuchi, T. Chemokines CCL19 and CCL21 promote activation-induced cell death of antigen-responding T cells. Blood 2007, 109, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Ferris, R.L.; Whiteside, T.L. Chemokine C receptor 7 expression and protection of circulating CD8+ T lymphocytes from apoptosis. Clin. Cancer Res. 2005, 11, 7901–7910. [Google Scholar] [CrossRef] [PubMed]
- Kannanganat, S.; Ibegbu, C.; Cheenreddi, L.; Robinson, H.L.; Amara, R.R. Multiple-cytokine-producing antiviral CD4 T cells are functionally superior to single-cytokine-producing cells. J. Virol. 2007, 81, 8468–8476. [Google Scholar] [CrossRef] [PubMed]
- Muller, M.; Carter, S.; Hofer, M.J.; Campbell, I.L. Review: The chemokine receptor CXCR3 and its ligands CXCL9, CXCL10 and CXCL11 in neuroimmunity-a tale of conflict and conundrum. Neuropathol. Appl. Neurobiol. 2010, 36, 368–387. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Rottman, J.B.; Myers, P.; Kassam, N.; Weinblatt, M.; Loetscher, M.; Koch, A.E.; Moser, B.; Mackay, C.R. The chemokine receptors CXCR3 and CCR5 mark subsets of T cells associated with certain inflammatory reactions. J. Clin. Investig. 1998, 101, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Sallusto, F.; Lenig, D.; Mackay, C.R.; Lanzavecchia, A. Flexible programs of chemokine receptor expression on human polarized T helper 1 and 2 lymphocytes. J. Exp. Med. 1998, 187, 875–883. [Google Scholar] [CrossRef] [PubMed]
- Foley, J.F.; Yu, C.R.; Solow, R.; Yacobucci, M.; Peden, K.W.; Farber, J.M. Roles for CXC chemokine ligands 10 and 11 in recruiting CD4+ T cells to HIV-1-infected monocyte-derived macrophages, dendritic cells, and lymph nodes. J. Immunol. 2005, 174, 4892–4900. [Google Scholar] [CrossRef] [PubMed]
- Petkovic, V.; Moghini, C.; Paoletti, S.; Uguccioni, M.; Gerber, B. ITAC/CXCL11 is a natural antagonist for CCR5. J. Leukoc. Biol. 2004, 76, 701–708. [Google Scholar] [CrossRef] [PubMed]
- Lajoie, J.; Juno, J.; Burgener, A.; Rahman, S.; Mogk, K.; Wachihi, C.; Mwanjewe, J.; Plummer, F.A.; Kimani, J.; Ball, T.B.; et al. A distinct cytokine and chemokine profile at the genital mucosa is associated with HIV-1 protection among HIV-exposed sero-negative commercial sex workers. Mucosal Immunol. 2012, 5, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Osakwe, C.E.; Bleotu, C.; Chifiriuc, M.C.; Crancea, C.; Otelea, D.; Paraschiv, S.; Petrea, S.; Dinu, M.; Baicus, C.; Streinu-Cercel, A.; et al. TH1/TH2 cytokine levels as an indicator for disease progression in human immunodeficiency virus type 1 infection and response to antiretroviral therapy. Roum. Arch. Microbiol. Immunol. 2010, 69, 24–34. [Google Scholar] [PubMed]
- Ogilvie, P.; Bardi, G.; Clark-Lewis, I.; Baggiolini, M.; Uguccioni, M. Eotaxin is a natural antagonist for CCR2 and an agonist for CCR5. Blood 2001, 97, 1920–1924. [Google Scholar] [CrossRef] [PubMed]
- Ogilvie, P.; Paoletti, S.; Clark-Lewis, I.; Uguccioni, M. Eotaxin-3 is a natural antagonist for CCR2 and exerts a repulsive effect on human monocytes. Blood 2003, 102, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Uguccioni, M.; Loetscher, P.; Forssmann, U.; Dewald, B.; Li, H.; Lima, S.H.; Li, Y.; Kreider, B.; Garotta, G.; Thelen, M.; et al. Monocyte chemotactic protein 4 (MCP-4), a novel structural and functional analogue of MCP-3 and eotaxin. J. Exp. Med. 1996, 183, 2379–2384. [Google Scholar] [CrossRef] [PubMed]
- Schnepple, D.J.; Shepard, B.; Bren, G.D.; Cummins, N.W.; Natesampillai, S.; Trushin, S.; Algeciras-Schimnich, A.; Meng, X.W.; Sainski, A.M.; Rizza, S.A.; et al. Isolation of a TRAIL antagonist from the serum of HIV-infected patients. J. Biol. Chem. 2011, 286, 35742–35754. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Erdmann, N.; Peng, H.; Herek, S.; Davis, J.S.; Luo, X.; Ikezu, T.; Zheng, J. TRAIL-mediated apoptosis in HIV-1-Infected macrophages is dependent on the inhibition of Akt-1 phosphorylation. J. Immunol. 2006, 177, 2304–2313. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.H.; Johnson, H.; Shu, H.B. Tumor necrosis factor-related apoptosis-inducing ligand receptors signal NF-κB and JNK activation and apoptosis through distinct pathways. J. Biol. Chem. 1999, 274, 30603–30610. [Google Scholar] [CrossRef] [PubMed]
- Emery, J.G.; McDonnell, P.; Burke, M.B.; Deen, K.C.; Lyn, S.; Silverman, C.; Dul, E.; Appelbaum, E.R.; Eichman, C.; DiPrinzio, R.; et al. Osteoprotegerin is a receptor for the cytotoxic ligand TRAIL. J. Biol. Chem. 1998, 273, 14363–14367. [Google Scholar] [CrossRef] [PubMed]
- Holen, I.; Croucher, P.I.; Hamdy, F.C.; Eaton, C.L. Osteoprotegerin (OPG) is a survival factor for human prostate cancer cells. Cancer Res. 2002, 62, 1619–1623. [Google Scholar] [PubMed]
- Qin, S.; Sui, Y.; Murphey-Corb, M.A.; Reinhart, T.A. Association between decreased CXCL12 and CCL25 expression and increased apoptosis in lymphoid tissues of cynomolgus macaques during SIV infection. J. Med. Primatol. 2008, 37, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Vicari, A.P.; Figueroa, D.J.; Hedrick, J.A.; Foster, J.S.; Singh, K.P.; Menon, S.; Copeland, N.G.; Gilbert, D.J.; Jenkins, N.A.; Bacon, K.B.; et al. TECK: A novel CC chemokine specifically expressed by thymic dendritic cells and potentially involved in T cell development. Immunity 1997, 7, 291–301. [Google Scholar] [CrossRef]
- Wurbel, M.A.; McIntire, M.G.; Dwyer, P.; Fiebiger, E. CCL25/CCR9 interactions regulate large intestinal inflammation in a murine model of acute colitis. PLoS ONE 2011, 6, e16442. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| K-Way and Higher-Order Effects | K | df | Likelihood Ratio Chi-Square | Sig. | Pearson Chi-Square |
|---|---|---|---|---|---|
| K-way and Higher Order Effects a | 1 | 7 | 616.1 | 0 | 1056.511 |
| 2 | 4 | 104.972 | 0 | 147.811 | |
| 3 | 1 | 22.957 | 0 | 25.08 | |
| K-way Effects b | 1 | 3 | 511.128 | 0 | 908.7 |
| 2 | 3 | 82.015 | 0 | 122.731 | |
| 3 | 1 | 22.957 | 0 | 25.08 |
| Cytokines | PCR Array Expression | Protein Array Expression | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Viremic CD4+ | Aviremic CD4+ | Viremic CD14+ | Aviremic CD14+ | Viremic CD8+ | Aviremic CD8+ | Viremic CD4+ | Aviremic CD4+ | Viremic CD14+ | Aviremic CD14+ | Viremic CD8+ | Aviremic CD8+ | |
| CCL18/PARC | 156.28 | – | – | – | – | – | 1.72 | – | – | – | – | −19.78 |
| CCL19/MIP-3B | 5.42 | 14.05 | 14.89 | 47.90 | 12.06 | 10.93 | – | 1.77 | 1.76 | – | – | – |
| CCL24/eotaxin-2 | −6.46 | −6.81 | −20.45 | −4.19 | −44.26 | −7.52 | – | −5.71 | −2.96 | −2.37 | −1.58 | −1.91 |
| CCL4/MIP-1- β | 5.97 | 6.16 | – | – | – | – | 1.63 | 3.23 | – | – | 14.75 | −2.67 |
| CCL5/RANTES | – | – | −107.00 | 16.94 | – | – | – | −1.76 | – | 2.73 | – | – |
| CCL8/MCP-2 | 199.19 | 39.18 | 68.40 | 25.49 | 18.40 | 50.56 | – | – | 2.49 | 12.47 | 1.71 | – |
| CD80 | – | – | 4.38 | 4.88 | – | – | – | – | 1.53 | 3.79 | – | – |
| CXCL10/IP-10 | 83.17 | 16.13 | 31.04 | 18.40 | 67.74 | 15.89 | – | – | 4.17 | 3.46 | – | – |
| CXCL11/I TAC | 180.77 | 66.81 | 78.58 | 103.39 | 311.27 | 41.93 | – | 7.14 | 9.88 | 8.26 | – | 13.26 |
| CXCL13/BLC | 6.05 | 26.03 | −5.29 | – | 4.41 | 8.69 | 2.32 | 5.90 | 1.54 | – | – | 1.55 |
| CXCL2/GRO | −10.72 | – | −29.94 | −7.20 | −13.34 | – | −1.58 | 1.59 | −6.47 | −1.82 | – | – |
| CXCL5/ENA-78 | −205.36 | −17.36 | −1041.18 | −37.22 | −14.91 | −18.64 | – | 1.98 | −14.91 | −2.82 | – | – |
| CXCL9/MIG | 79.23 | 28.29 | 11.52 | 43.77 | 94.48 | 34.06 | – | – | 3.44 | 1.61 | – | – |
| IL8 | – | – | −11.66 | – | – | – | – | 1.55 | −2.24 | – | 1.57 | – |
| VCAM1 | 32.85 | 15.89 | 109.29 | 60.38 | – | – | – | – | 1.55 | – | – | – |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perera, S.S.; Wang, B.; Damian, A.; Dyer, W.; Zhou, L.; Conceicao, V.; Saksena, N.K. Retrospective Proteomic Analysis of Cellular Immune Responses and Protective Correlates of p24 Vaccination in an HIV Elite Controller Using Antibody Arrays. Microarrays 2016, 5, 14. https://0-doi-org.brum.beds.ac.uk/10.3390/microarrays5020014
Perera SS, Wang B, Damian A, Dyer W, Zhou L, Conceicao V, Saksena NK. Retrospective Proteomic Analysis of Cellular Immune Responses and Protective Correlates of p24 Vaccination in an HIV Elite Controller Using Antibody Arrays. Microarrays. 2016; 5(2):14. https://0-doi-org.brum.beds.ac.uk/10.3390/microarrays5020014
Chicago/Turabian StylePerera, Suneth S., Bin Wang, Arturo Damian, Wayne Dyer, Li Zhou, Viviane Conceicao, and Nitin K. Saksena. 2016. "Retrospective Proteomic Analysis of Cellular Immune Responses and Protective Correlates of p24 Vaccination in an HIV Elite Controller Using Antibody Arrays" Microarrays 5, no. 2: 14. https://0-doi-org.brum.beds.ac.uk/10.3390/microarrays5020014