Compared to other prokaryotes, the photosynthetic machinery in cyanobacteria imposes higher demand for metals. Thus, Fe is necessary for all three photosynthetic electron transfer chain complexes, Mn for the water-splitting complex, Cu for plastocyanin and cytochrome c oxidase, Mg for chlorophylls, and Zn for carbonic anhydrase [
12]. Nickel, along with Zn, Cu, Mo, and Fe, are essential nutrients for microorganisms since they participate in various cellular processes [
25]. According to literature data, nickel is present in the main part of commercially available spirulina products. Thus, in 23 samples of spirulina produced in Australia, the USA, Great Britain, Japan, India, Canada, and New Zealand, its content varied from 0.21 to 4.6 mg/kg d.w. [
8]. In the present study, nickel content in control spirulina biomass was 11.8 mg/kg.
3.1. Ni System
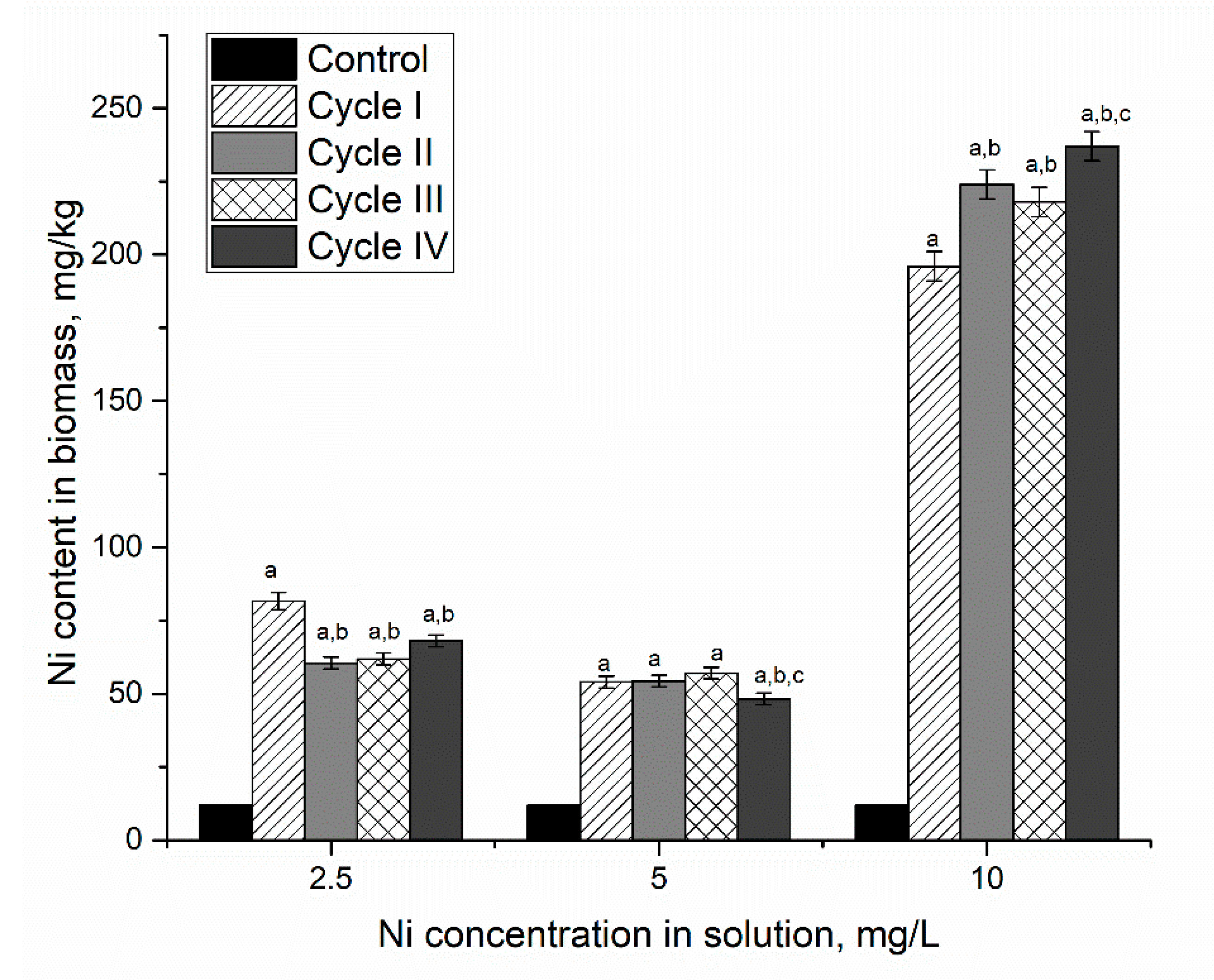
The addition of nickel ions at different concentrations (according to
Table 1) to the cultivation medium increased their content in the biomass (
Figure 1). That is in agreement with Rugnini et al. [
21]. However, the efficiency of metal uptake depended on the cycle of cultivation. Thus, in the system containing 2.5 mg/L of nickel ions, the highest accumulation of nickel ions was noticed during the first cultivation cycle, when the biomass accumulated 7 times more nickel ions than control. Over the next three cultivation cycles, their content in biomass was 5.1–5.7 times higher than control but lower than in the first cycle. At nickel concentration in solution 5 mg/L, its content in biomass was 4.6–4.7 times higher than in control. Thus, the accumulation of nickel ions by
Spirulina platensis biomass at concentrations in the solution of 2.5 and 5 mg/L was almost at the same level. Increased nickel concentration up to 10 mg/L in solution led to a significant increase (
p < 0.001) of its content in biomass, 17–20 times in comparison with control. The most pronounced increase was observed in the first cycle of cultivation, by 1560%, with respect to control. In the next three cultivation cycles, the increase in nickel uptake was less significant. Thus, in the second cycle, it was 14% higher than in the first cycle. In the third cycle, a slight decrease (by 2.3%) was noticed, while in the fourth cycle, an increase of 8.7% took place.
Cyanobacteria
Anabaena cylindrica,
Anabaena flos-aquae, and
Nostoc sp. accumulated 2–6 mg/g of nickel at a 10 mg/L concentration in solution [
26]. The uptake of nickel ions in the present study was lower compared to values presented by Rugnini et al. [
21] for two cyanobacteria,
Chlorella vulgaris and
Desmodesmus sp., although the range of the studied concentrations in both studies was very similar. The maximum accumulation of nickel by
Synechococcus sp. IU 625 occurred on the fifth day of biomass growth; however, nickel was undetectable inside the cells on day 11. The release of nickel ions from biomass can be explained by overexpression of the smtA gene and other alternative mechanisms. At the same time, it should be mentioned that
Synechococcus sp. IU 625 demonstrated high resistance to nickel ions, maintaining high biomass growth rates at nickel concentrations in the cultivation medium of 10 and 25 mg/L [
27].
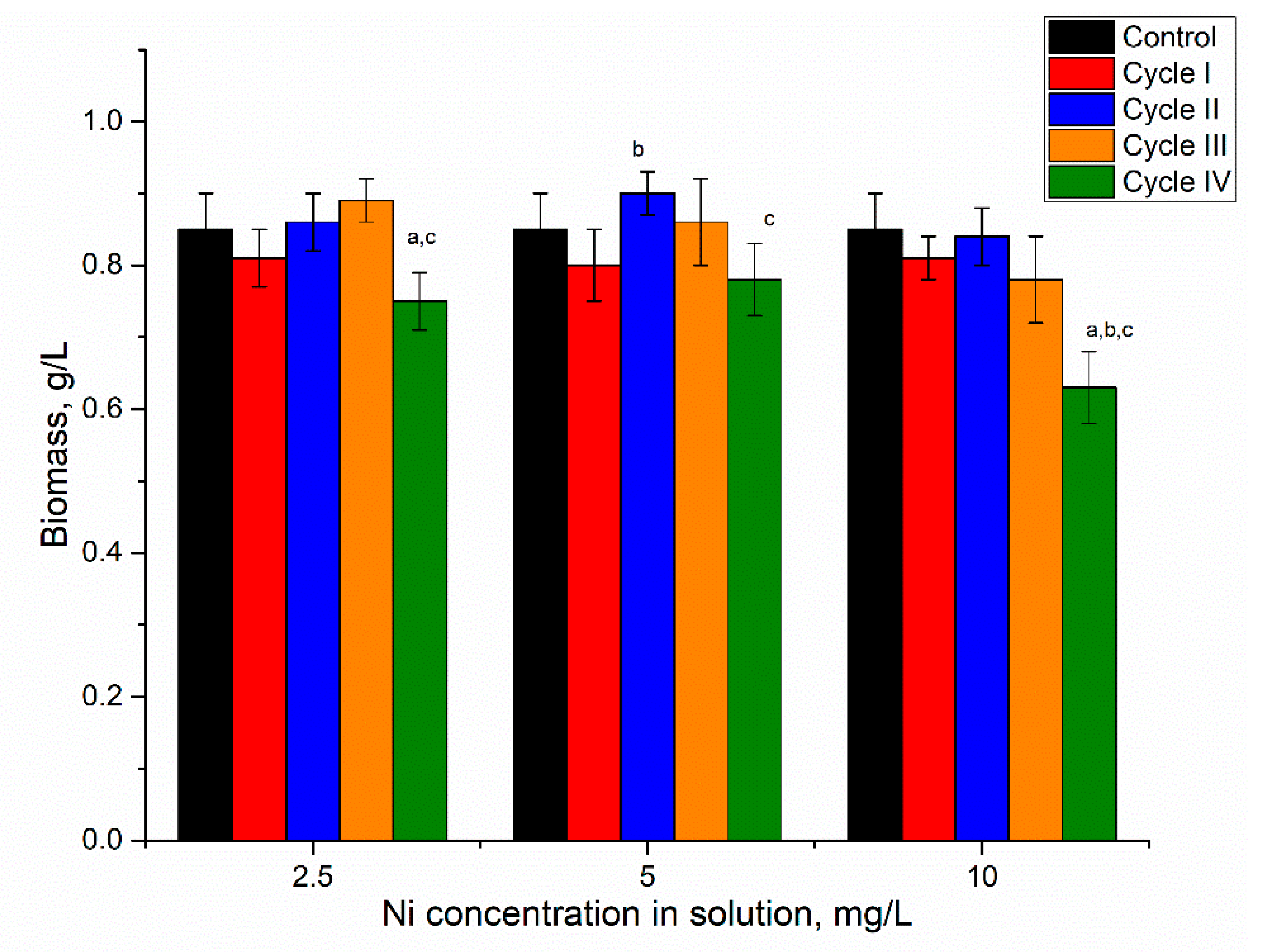
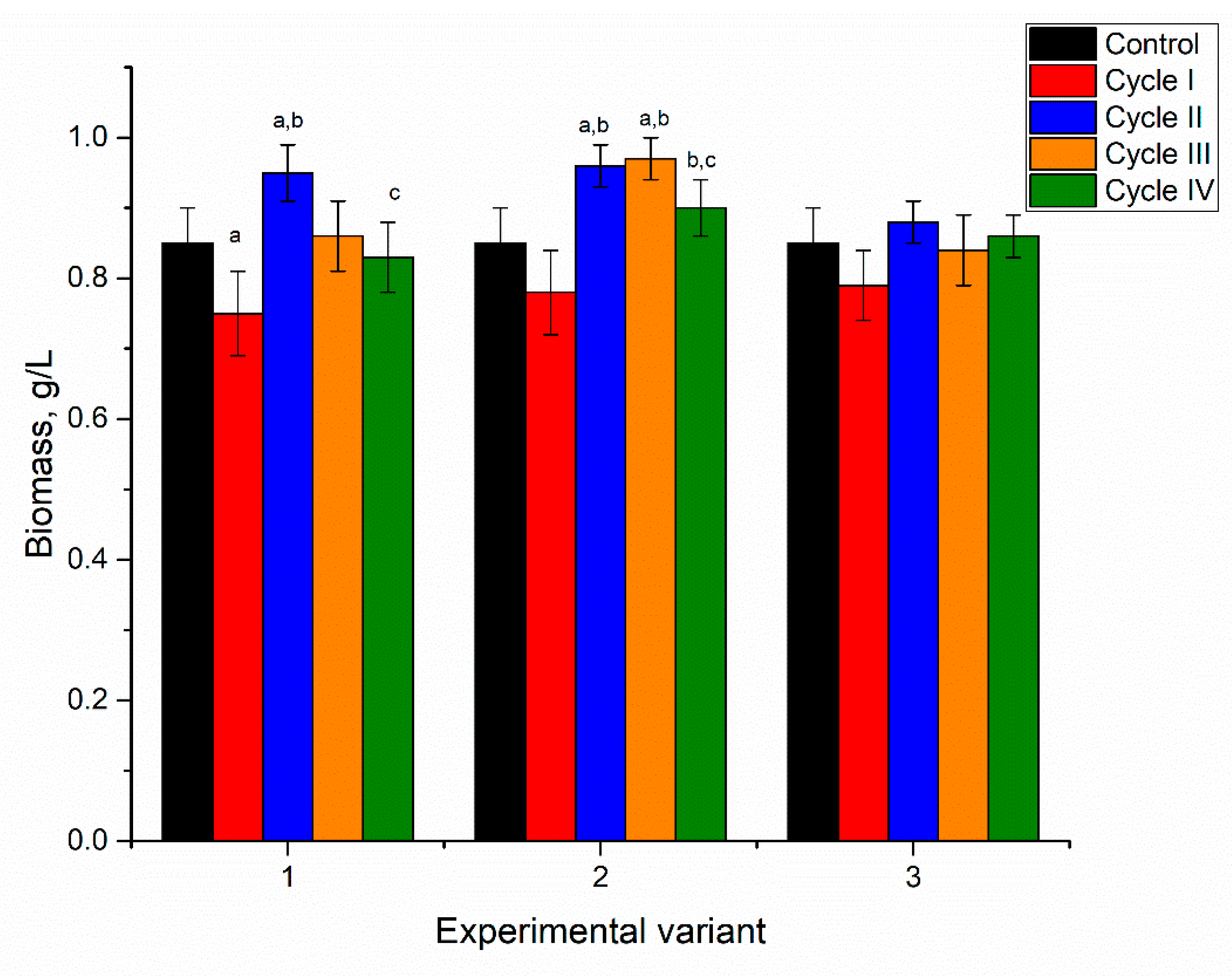
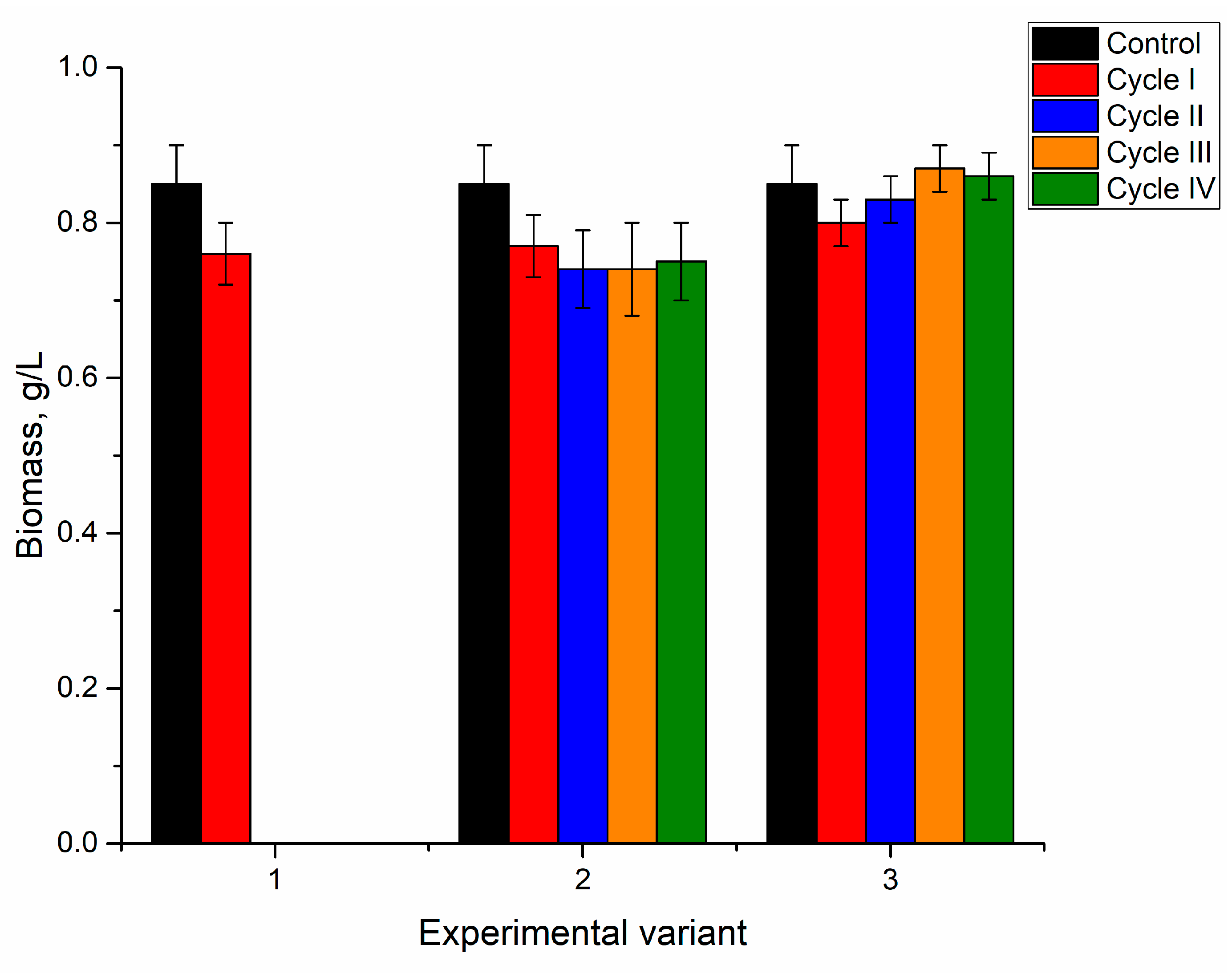
The addition of nickel ions to the medium in concentrations of 2.5–10 mg/L for the first cultivation cycle did not significantly affect spirulina biomass accumulation (
Figure 2).
At the end of the first cultivation cycle, the biomass was reduced insignificantly by 5–6%, and this response was the same for all applied nickel concentrations. In the next two cycles, in all analyzed systems, the amount of biomass returned to control values. At the end of the fourth cycle, the amount of biomass was reduced in all analyzed systems, as the resulting values were 0.73, 0.78, and 0.75 g/L for nickel concentrations at 2.5, 5, and 10 mg/L in the medium, respectively. For a nickel concentration of 2.5 mg/L, biomass reduction in the fourth cultivation cycle was significant (
p < 0.001) compared to the second and third cycles. At a nickel concentration of 10 mg/L, the decrease in the fourth cycle was significant (
p < 0.001) compared to control and other cycles of biomass growth, while at a concentration of 5 mg/L the reduction was insignificant. Thus, the reported in [
28] dose-dependent decrease of
Spirulina indica,
Spirulina maxima, and
Spirulina platensis growth with the increase of nickel concentration in the medium was not observed in the present work even at repeated cultivation.
Spirulina platensis is one of the richest protein sources of microbial origin containing 46–63% dry biomass (DB) of this nutrient [
29]. Proteins serve multiple purposes as structural components, enzymes, membrane transporters, signaling molecules, or regulatory factors. In cyanobacteria, some proteins as, for example Psb27, are important in photosystem II (PSII) repair, while others, like PetP, are involved in the stress adaptation of the photosynthetic electron transport [
30].
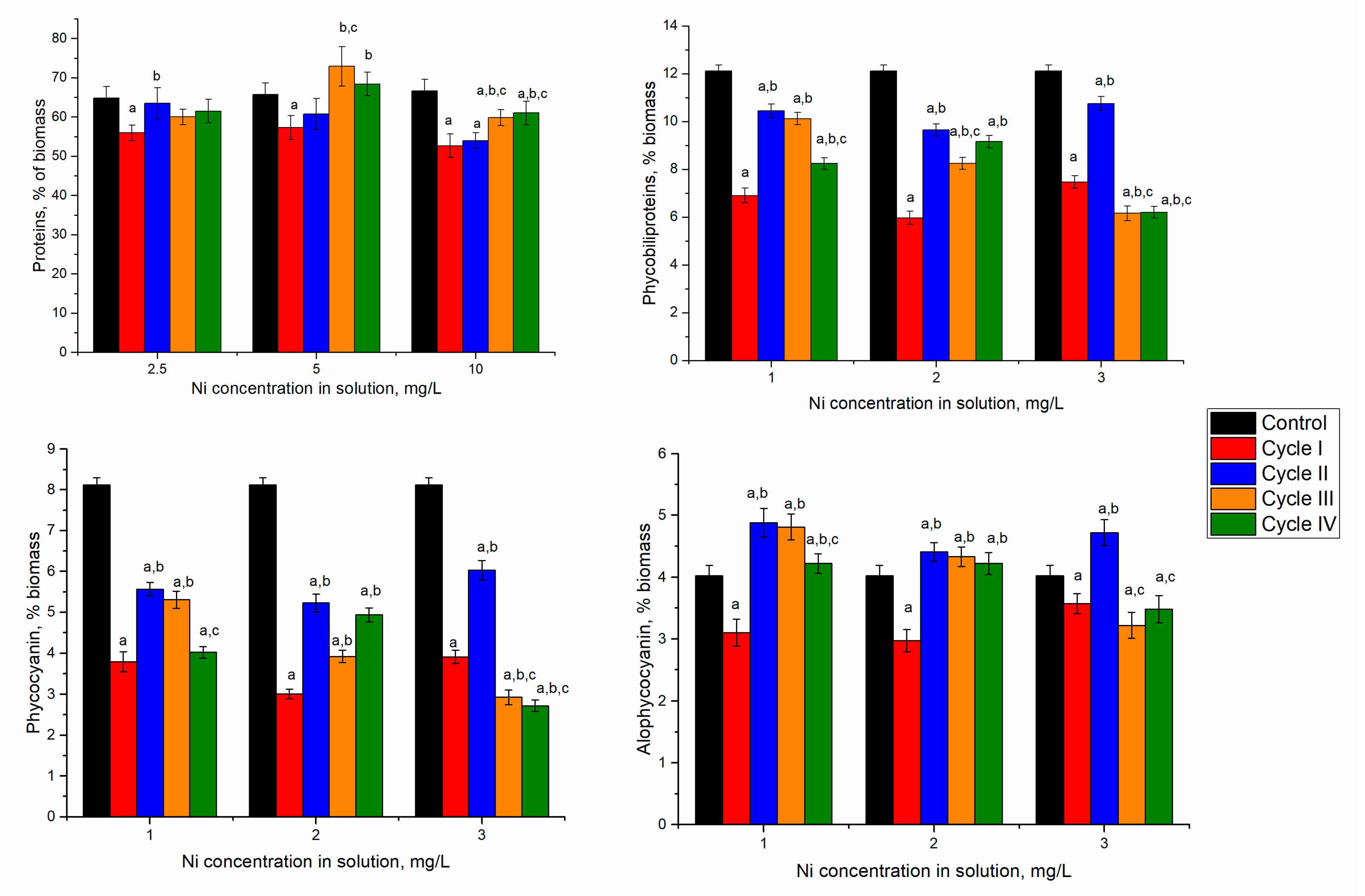
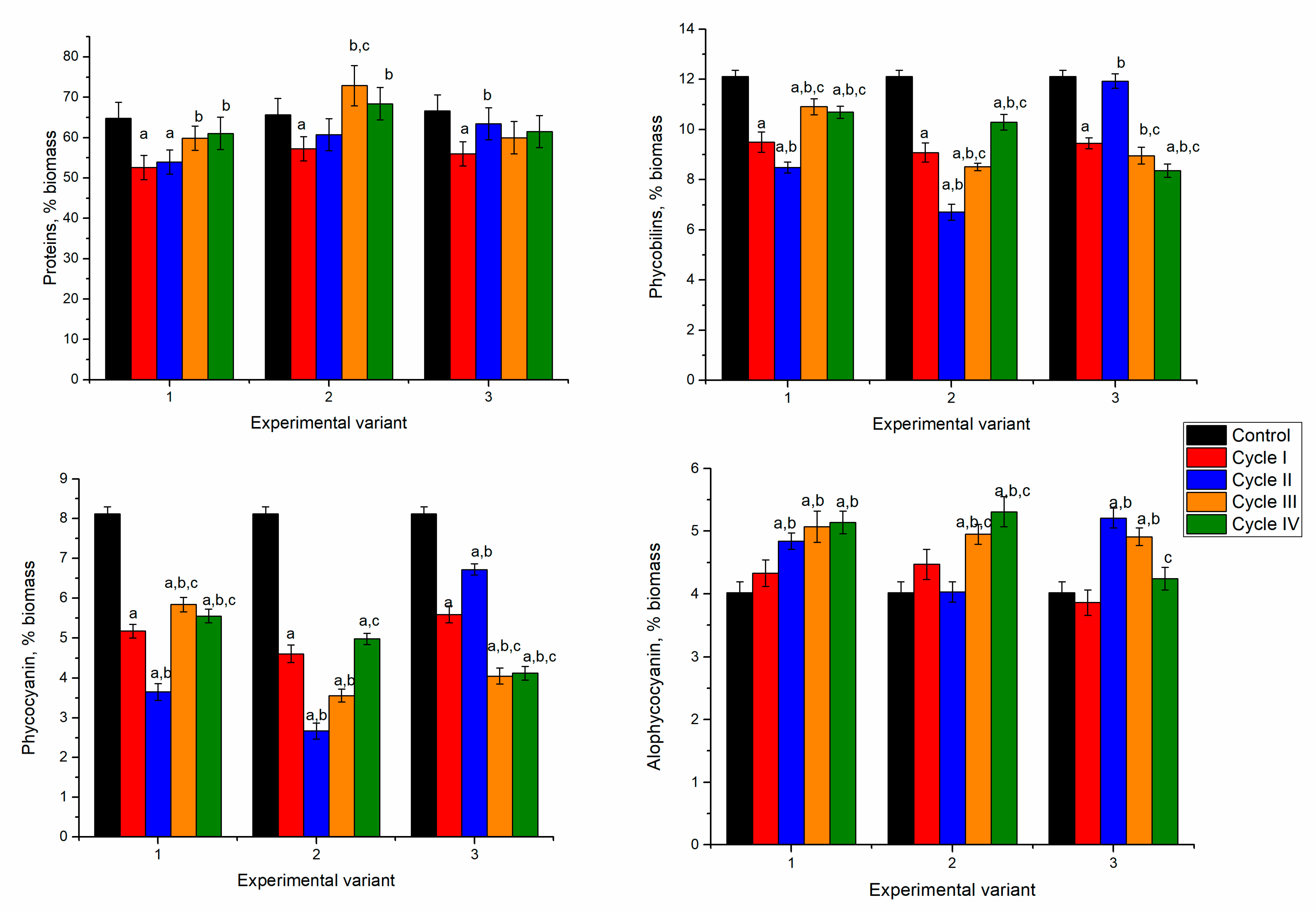
Spirulina’s response to nickel ion effects in the first cultivation cycle was manifested by reducing biomass and protein content. However, in the next cycles, spirulina adaptation to the new conditions of growth was observed (
Figure 3).
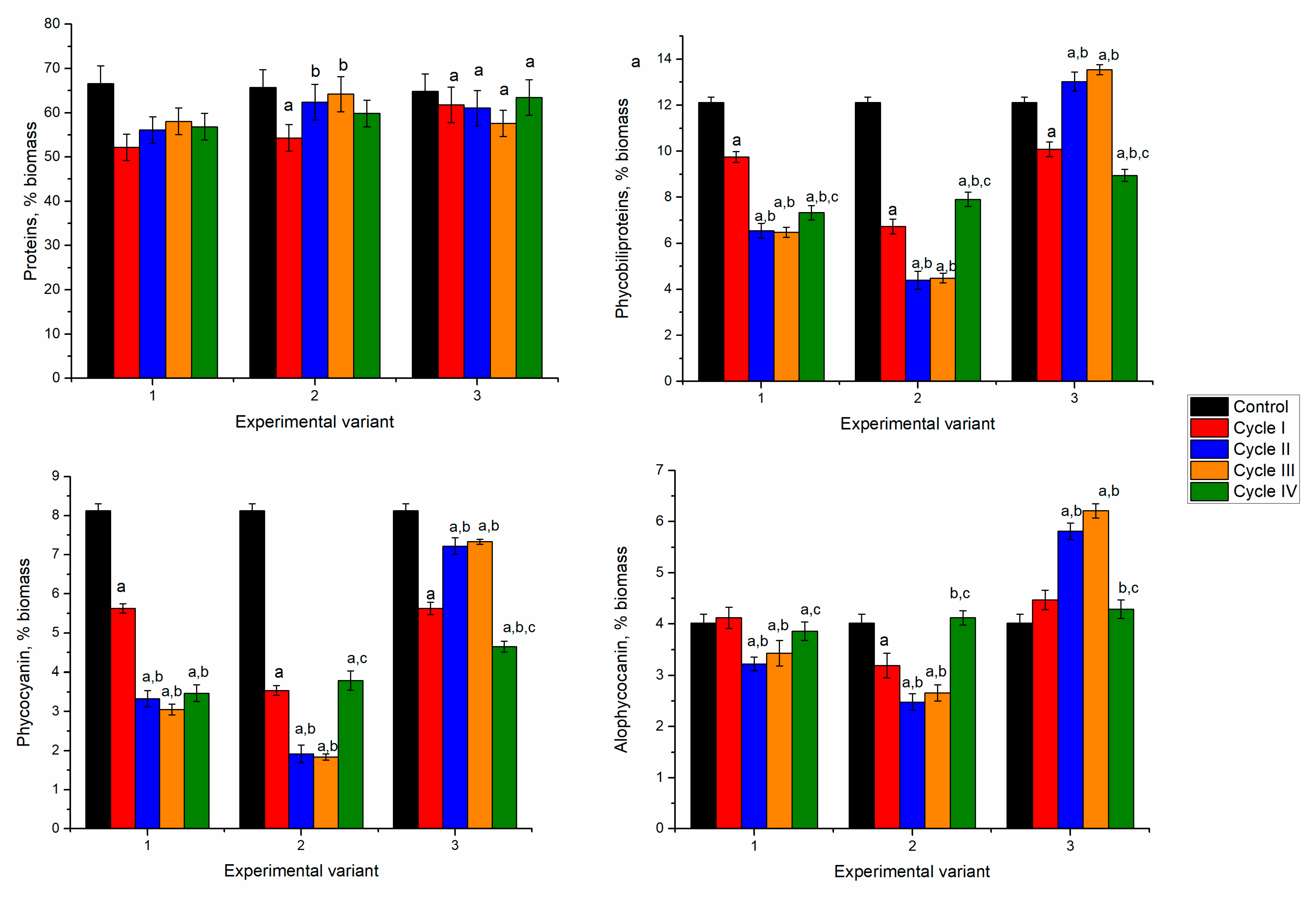
In spirulina biomass growing in the media containing 2.5 and 5 mg/L of nickel ions, at the end of the first cultivation cycle, the content of proteins in biomass significantly decreased by 12–15% (
p < 0.001). Nickel concentration of 10 mg/L led to the reduction of protein content by 19%. This could be due to the stress caused by the nickel ions present in the culture medium. In microorganisms, nickel toxicity is expressed in the inhibition of metalloproteins, interaction with enzyme active site His or Cys residues, and enhancement of oxidative stress [
31]. Starting with the second cultivation cycle, at all nickel concentrations, the protein content of biomass increased in comparison with the first cycle. At a nickel concentration of 2.5 mg/L in the medium, the content of proteins over the next three cycles almost reached control values. The difference between the second–fourth cycles and the first cycle was statistically significant (
p < 0.001). At a nickel concentration of 5 mg/L in the medium, the maximum amount of proteins was attained in the third cultivation cycle. This value was significantly higher (
p < 0.001) than the first two cycles, but it did not differ essentially from control.
The increase in protein content can be explained by the excretion of nickel ions from the cell. Huertas et al. [
27] showed that an excess of nickel that could cause a perturbation of protein function is counterbalanced by the presence of efflux pumps that expel it out of the cell. The slight increase or decrease of nickel content compared to the first cycle confirmed this fact.
In cyanobacteria, phycobiliproteins, large water-soluble supramolecular protein aggregates serve as major accessory pigments in photosynthesis. They can be divided broadly into three classes: phycoerythrin, phycocyanin, and allophycocyanin, based on their spectral properties [
32]. Spirulina contains two of these pigments—phycocyanin and allophycocyanin. In all experimental variants, the amount of phycobiliproteins was significantly lower (
p < 0.001) compared to control (
Figure 3).
In the experimental variants with nickel concentrations of 2.5 and 5 mg/L, at the end of the first cultivation cycle the content of total phycobiliproteins in biomass was reduced by 43–51%. Consecutive cultivation in the second cycle was beneficial for the culture. The amount of phycobilin increased by 51% and 61% compared to the first cycle. However, it was lower by 13 and 20% compared to control biomass. The tendency to restore phycobiliprotein content was maintained for two further cultivation cycles. At the end of the fourth cycle, the phycobiliprotein content increased by 19 and 53%, respectively, compared to values obtained in the first cycle. At the same time, it was lesser than control by 32% and 24%, respectively.
In the case of the application of 10 mg/L, the decrease of the phycobiliprotein content by 38% in the first cycle was followed by an increase of 44% in the second cycle, and in the next two cycles it remained slightly lower compared to the first cycle. Thus, the introduction of nickel ions in the culture medium induced the inhibition processes of phycobiliprotein synthesis, regardless of the applied concentration. A dose-dependent decrease of protein content with an increase in nickel concentration was observed for Spirulina strains [
33].
Significant reduction in phycobiliprotein content was mainly determined by decreasing phycocyanin by more than 50% in the first and fourth cycles of biomass growth (
Figure 3). By adding 2.5 mg/L of nickel in the first cycle, the phycocyanin content decreased by 53%, compared to 23% in control and allophycocyanin. A recovery attempt was observed in the second and third cycles, but phycocyanin values remained low (30–35% below the control values). In the case of allophycocyanin, its content in the second cycle increased significantly by 21% (
p < 0.001) compared to control and was maintained in the third cycle. In the last cycle, the allophycocyanin content was at the control level.
At the introduction of 5 mg/L of nickel in the cultivation medium and the end of the first cultivation cycle, phycocyanin content decreased by 63% and allophycocyanin by 26% compared to control. The tendency to rectorate the phycocyanin content during repeated cultivation in the medium containing nickel ions failed. Its content in the fourth cycle was 39% lower than the control value. At the same time, the content of allophycocyanin returned to the control values. The least favorable situation was observed at a nickel concentration of 10 mg/L when in the first cycle of cultivation, the phycocyanin content was reduced by 52% and at the end of the fourth cycle by 66%. The content of allophycocyanin was significantly lower (p < 0.001) compared to control in the first, third, and fourth cycles and higher in the second cycle. Thus, upon repeated contact with the metal in the second cycle, the spirulina culture restored its allophycocyanin level, but the repeated action of the pollutant eventually leads to a stable reduction of the pigment content.
It is known that both phycocyanin and allophycocyanin bind heavy metal ions differentially, whereas phycocyanin has a higher affinity for them. Thus, metal binding preferentially to phycocyanin can be one of the reasons explaining the significant decrease in their content [
34]. The same authors reported that silver and copper ions show minimal binding to allophycocyanin, as they induce structural changes such as a decrease in absorbance and fluorescence of phycocyanin.
3.2. Ni/Cr/Fe System
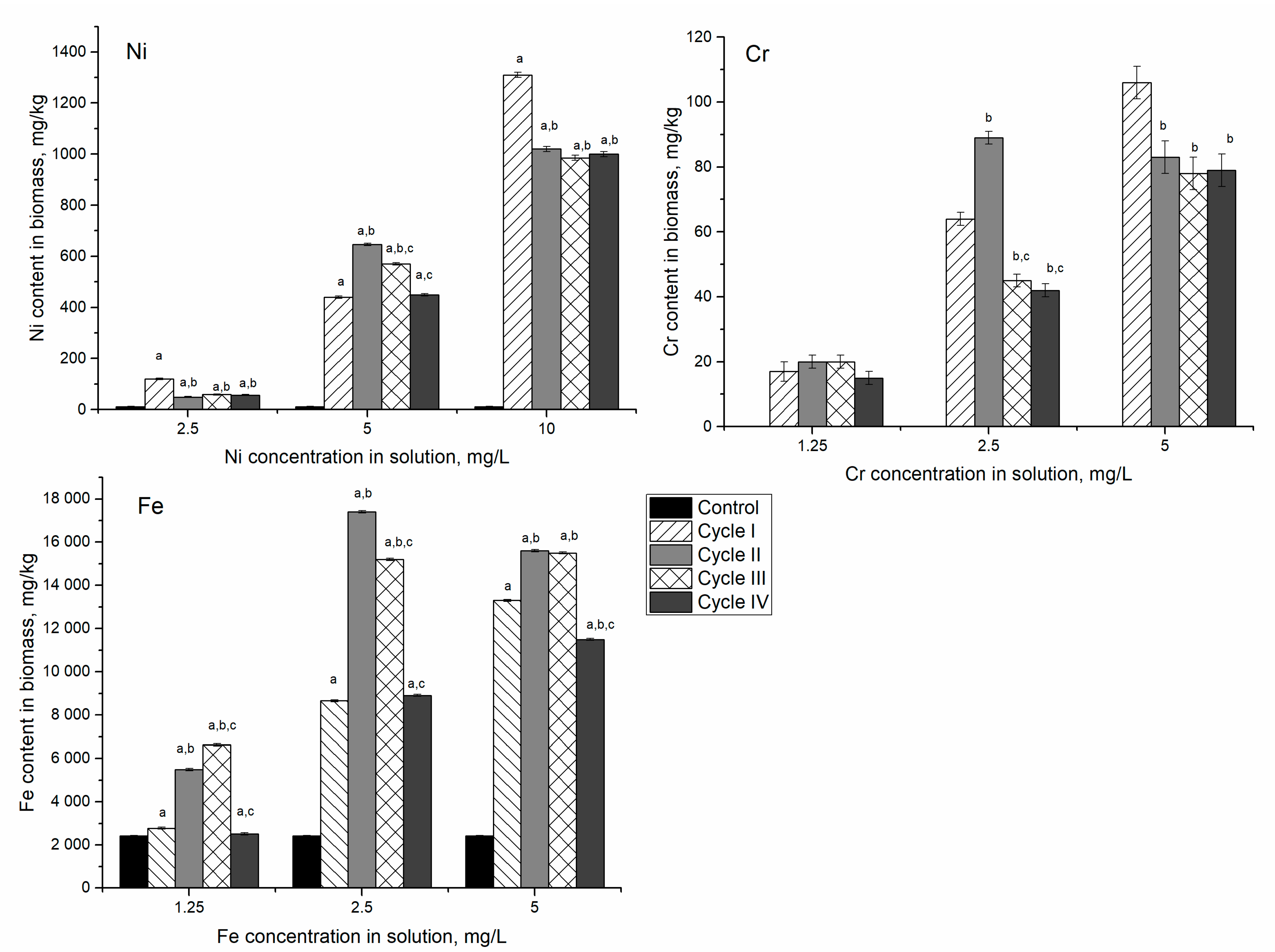
In the Ni/Cr/Fe system at nickel concentration in the solution of 2.5 mg/L, after the first cycle of cultivation, nickel content in biomass was 10 times higher than control (
Figure 4). In the next three cycles, its content in biomass decreased approximately twice and was almost on the same level. At nickel concentration in solution 5 mg/L, its content in biomass increased 37–55 times.
The most significant increase was noticed in the second cycle when nickel content in biomass exceeded control value by 83–111 times. At nickel concentration of 10 mg/L, the most pronounced increase was observed in the first cultivation cycle, then in the next three cycles, it decreased by 22–25% compared to the first cycle. The amount of Ni accumulated by the biomass in Ni/Cr/Fe systems was 1.5–11 times higher than in the system containing only nickel ions.
Chromium content in control biomass was below the detection limit of NAA. However, its content in the system containing Cr(VI) ions varied from 17 to 110 mg/kg. At a chromium concentration of 1.25 mg/L, the content of the element accumulated in biomass was approximately the same during all cultivation cycles. At a concentration of 2.5 mg/L in the first cycle of cultivation, the spirulina biomass accumulated 3 times more chromium compared to the medium with a concentration of 1.25 mg/L. In the second cycle, this amount increased to 90 mg/kg. In the next two cycles, the amount of chromium accumulated decreased twice compared to the second cycle. At chromium concentration of 5.0 mg/L, the highest amount of the element was accumulated in the first cycle of cultivation, after which it decreased and remained at the same level for the next three cycles.
Iron accumulation in biomass also increased with the increase of its concentration in the solution. With the addition of 1.25 mg/L of iron ions, the content in biomass continuously increased over three cultivation cycles, and then it decreased to the level of control. At higher Fe concentrations, 2.5 and 5 mg/L, the same pattern was observed. However, in these experimental variants, after four cultivation cycles, Fe content in biomass was 3.7 and 4.7 times higher than in control, respectively. As in the case of the Ni system, the removal of metal ions from cells to decrease the toxic effect on spirulina was observed.
For spirulina grown in the medium containing ions of three metals, nickel, chromium, and iron, at the end of the first cycles in all experimental variants, the biomass decreased by 7–12%. However, only in the case of the first experimental variant was this difference significant (
p < 0.001) compared to control. In the other cases, there was a tendency for reduced biomass productivity (
Figure 5). In the next three cycles, biomass was on the level of control or overpassed it. Thus, in the first experimental variant, the amount of biomass in the second cycle was significantly higher than control. In the second experimental variant, the amount of biomass was higher than control in the second and third cycles. In the rest of the cases, the obtained values were at the level of control.
While assessing the effect of nickel concentration on biomass, it was noticed that at concentrations of 2.5 mg/L and 10 mg/L, the maximum metal accumulation took place in the first cycle of cultivation, leading to decreased spirulina productivity. In the next two cycles, nickel content in biomass was reduced by 2 and 1.3 times, respectively, while biomass increased by 12–14% compared to control. The highest accumulation of iron, in all experimental variants, was observed in the second and third cycles. Thus, dependence between the accumulation of Fe and biomass production can be assumed, although it occurred at levels characteristic of the strain under study. At the same time, it should be noted that in the nickel mono-system, in the first and third experimental variants, the amount of biomass was significantly lower than control, while in the analyzed poly-systems, this effect was not observed.
Protein content in the first experimental variant (at a Ni concentration of 2.5 mg/L) was not significantly affected by metal ions in the cultivation medium during repeated cultivation (
Figure 6).
For the second and third variants (5 mg/L and 10 mg/L), the content of proteins at the end of the first cycle was significantly reduced by 17–20% (p < 0.001). A tendency to restore the protein content towards the end of the fourth cycle was observed. In the second experimental variant, the amount of protein in spirulina biomass at the end of the second–fourth cycles did not differ significantly from control. The situation was different in the third experimental variant. Although there was a tendency to restore the protein content in cycles II-IV, it remained significantly lower than control.
In the first experimental variant, the content of phycobiliproteins decreased insignificantly, by 19% in the first cycle of cultivation and by 39–46% in the next three cycles (
Figure 6). The phycocyanin content decreased drastically, by 31–62%, compared to control. The allophycocyanin content did not change significantly. The maximum decrease of 19% took place in the second cycle.
In the second experimental variant, a critical decrease in the content of phycobiliproteins took place in the second and third cycles. Their value was 63% lower than the control. At the end of the fourth cultivation cycle, the content of phycobiliproteins remained low, the value 35% lower than control. The phycocyanin content decreased by 56% in the first cycle and 76–77% in the second and third cycles. The allophycocyanin content was reduced by 20–38% during the first three cultivation cycles and returned to normal values in the fourth cycle.
It is interesting that in the third experimental variant, the content of phycobiliproteins changed less during repeated cultivation and decreased by 26% compared to control in the fourth cycle. Content of phycocyanin in the first and fourth cycles was also significantly reduced, by 31–43% (
p < 0.001), while the allophycocyanin content remained almost unchanged. In the second and third cycles, the content of phycobiliproteins and phycocyanin did not change, and an increase in the content of allophycocyanin by 45% was noted. In our previous study, we showed that in spirulina biomass grown in a Cr/Fe system, the content of phycobiliproteins was reduced by approximately 70% [
18]. Since Bellamy-Carter and co-authors [
34] reported little to no binding of iron ions by phycobilins, it can be suggested that nickel and chromium are responsible for the decrease of phycobiliproteins.
3.3. Ni/Cu/Sr/Zn System
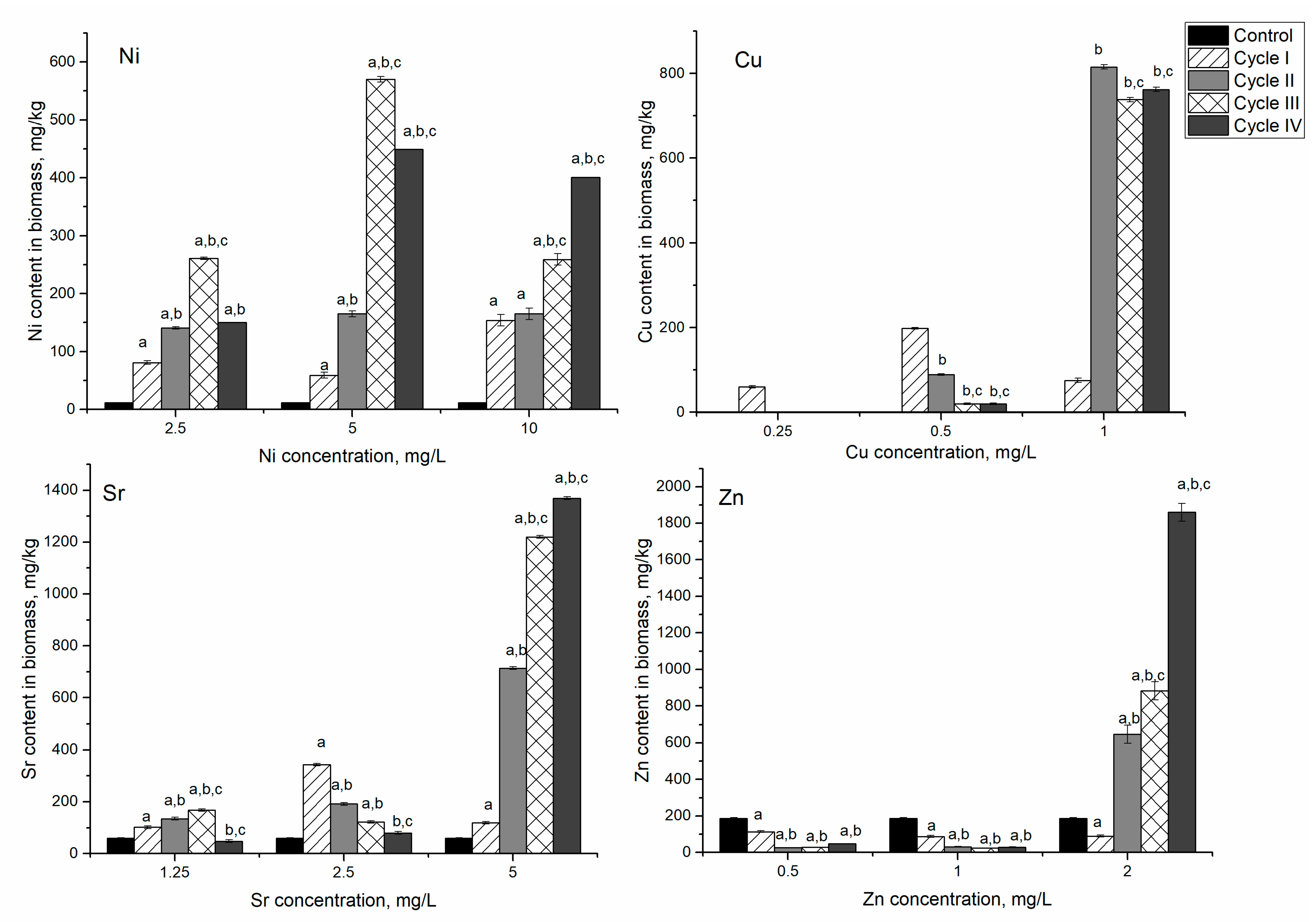
In the Ni-Cu-Sr-Zn system in the first experimental variant, the nickel content in biomass increased during three cycles of cultivation by 6.9–22 times compared to control. In the fourth cycle, it declined to the level of the second cycle (
Figure 7). The level of nickel accumulation was higher compared to the monometallic system, where the maximum amount of nickel, 80 mg/kg, was accumulated in the first cycle; and compared to the Ni/Cr/Fe system, where biomass accumulated up to 100 mg/kg of nickel in the first cycle of cultivation. In the analyzed system, maximum accumulation of nickel (250 mg/kg) was achieved in the third cycle, and it was 2.5–3.0 higher than the Ni and Ni/Cr/Fe systems.
Copper was detected in biomass only after the first cycle of cultivation. Accumulation of strontium by biomass was similar to the nickel uptake. It increased over three cultivation cycles, and then, in the fourth cycle, it decreased below the control level. Zinc was the only element whose content in biomass continuously decreased from 186 mg/kg in control biomass to 23 mg/kg after four cycles of cultivation.
In the second experimental variant, maximum metal accumulation was achieved in different cycles: nickel (646 mg/kg) in the third cycle of cultivation, and copper (192 mg/kg) and strontium (343 mg/kg) in the first cycle. Then their content in biomass decreased in comparison to the maximum accumulated amount, but it was higher than in control biomass. For zinc, it was noted the same pattern as in the first experimental variant, as its content in biomass decreased by 86% compared to control. The decrease of zinc content in biomass can be explained through replacement by other metal ions, in particular copper, present in the analyzed system. Copper displays a high affinity for metalloproteins, and if equivalent quantities of all divalent metals were present, proteins would probably all bind copper [
35]. The decrease in zinc content can also be explained by the inhibition of zinc metalloenzymes by nickel [
36]. The reduction of strontium content can also be explained by its replacement with nickel ions [
33]. In general, the content of nickel in biomass increased 5–55 times, of Cu 20–198 times, and of Sr 1.3–5.7 times.
An interesting pattern was observed in the third experimental variant. Nickel, strontium, and zinc content in biomass continuously increased over the cycles. Thus, the amount of nickel accumulated by biomass increased from 1200% in the first cycle to 3300% in the fourth cycle compared to control. Zinc behavior in the third experimental variant was different from the other experiments. It was characterized by the increase of its content in biomass by a factor of 10 with respect to control. The maximum amount of copper was accumulated by the biomass in the second cycle of cultivation (815 mg/kg), and then it slightly decreased in the next two cycles of cultivation (approximately 9%). The possible mechanisms of cyanobacteria resistance to heavy metals include complexation and active efflux [
37]. Accumulation of lesser nickel content compared to the second experimental variant indicated that its efflux is the main mechanism of spirulina resistance. Continuous accumulation of strontium and zinc suggested their binding to metallothioneins, which are synthesized under heavy metal stress [
37]. In the case of copper, both mechanisms of cyanobacteria resistance are possible.
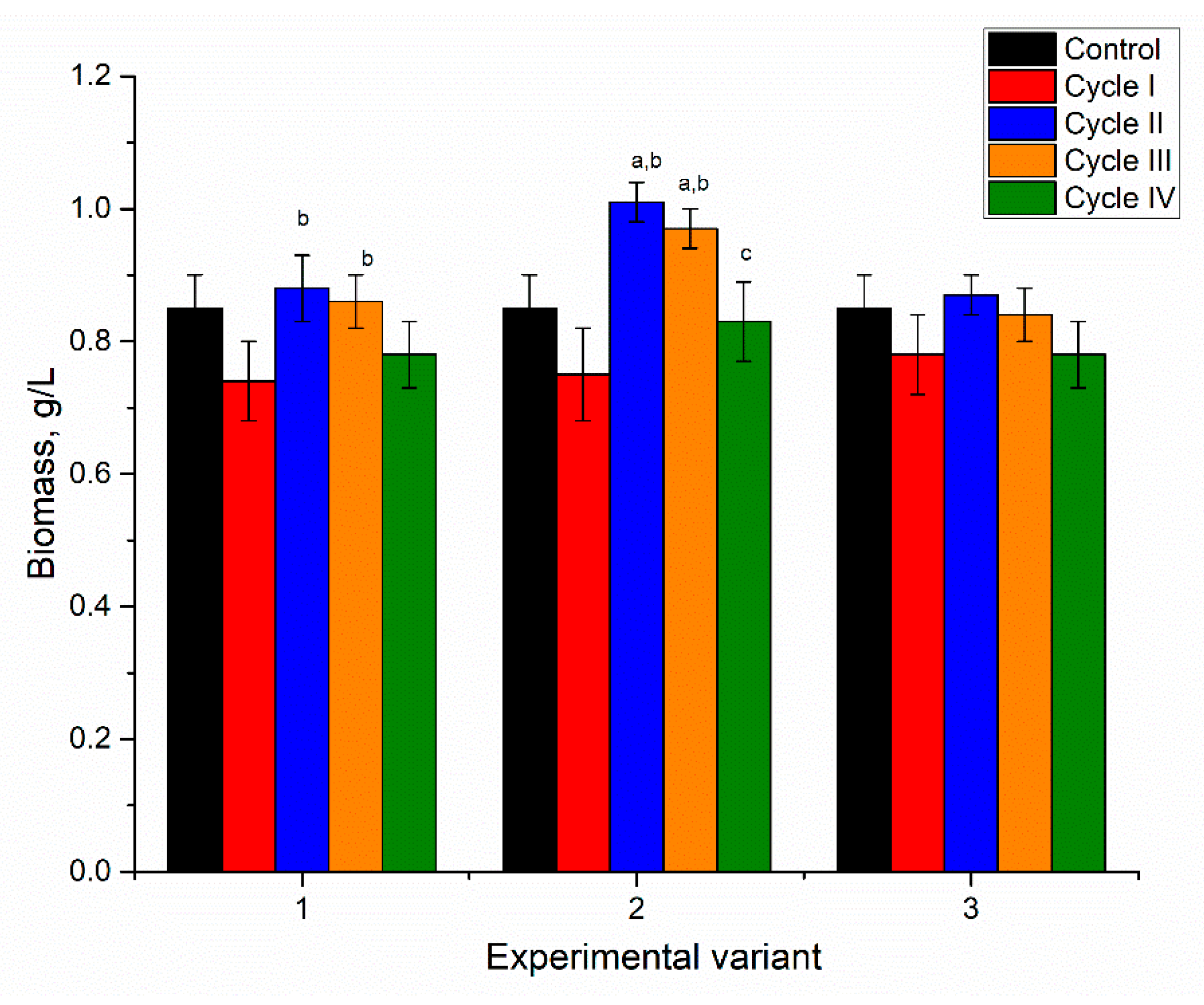
Accumulation of nickel, copper, zinc, and strontium in spirulina did not have a negative impact on biomass production during consecutive cultivation in a metal-loaded medium (
Figure 8). In all experimental variants, the culture’s response to the introduction of metals in the cultivation medium was similar to the Ni system. It was characterized by a decrease in biomass by 10% compared to control at the end of the first cycle. This difference was statistically insignificant.
In the first and third experimental variants over the next three cycles, the biomass was maintained at the level of control. In the second experimental variant, there was an increase in biomass productivity by 14–18% at the end of the second and third cultivation cycles. The maintenance of high biomass productivity under loading of chemical elements indicated adaptation of spirulina biomass to the new growth conditions.
The content of proteins in spirulina grown with the addition of Ni/Cu/Sr/Zn was also insignificantly affected by the presence of metal ions (
Figure 9). As in the case of biomass productivity, in all experimental variants, the content of proteins in biomass at the end of the first cycle decreased by 13–15% with respect to control. Restoration of the protein content took place in consecutive cultivation cycles and reached the level of control.
The addition of elements according to the first experimental variant resulted in decreased content of phycobiliproteins at the end of the first cycle by 21%. It maintained its low in the second cycle, reaching values approximately equal to control, but in the third and fourth cycles it was significantly lower. In the second experimental variant, the critical content of phycobiliproteins was obtained during the second cycle of cultivation, when their content was 44% lower than control. At the end of the fourth cycle of cultivation, the phycobiliprotein content was restored, and the values were 15% lower than control. In the case of the third experimental variant, in the first cycle, the phycobilin content decreased by 22% compared to control. In the second cycle, their content was approximately equal to control. However, in the next cycles, it was reduced again to the level of the first cycle.
The significant reduction in phycobiliprotein content was caused by decreased phycocyanin quantity. In the first experimental variant, during the first two cultivation cycles, the phycocyanin content decreased by 63% compared to the control, while in the third and fourth cycles, its content increased and reached a value 32% lower than control. The allophycocyanin content increased over the four cultivation cycles by 20–28%.
The same tendency was observed in the second experimental variant. In the first two cycles, the maximum reduction of phycocyanin was 67%. In the next two cycles, it started to increase, and by the end of the fourth cycle, it was lower than control by 39%. The allophycocyanin content increased during cultivation by 23–32%.
A different response was established for the third experimental variant, in which, in the first cycle of cultivation, the phycocyanin content decreased by 31% below the control. However, it tended to get restored in the second cycle, but it was again reduced in the next two cycles. Allophycocyanin, which content in the first cycles was on the level of control, increased in the next cycles by 30% compared to control. Zinc, copper, and nickel can be responsible for decreasing phycobiliprotein content. Reducing phycobiliprotein content in the presence of zinc, copper, and nickel ions in the cultivation medium were also reported in other studies [
17,
38,
39].
3.4. Ni/Cu/Zn/Mo System
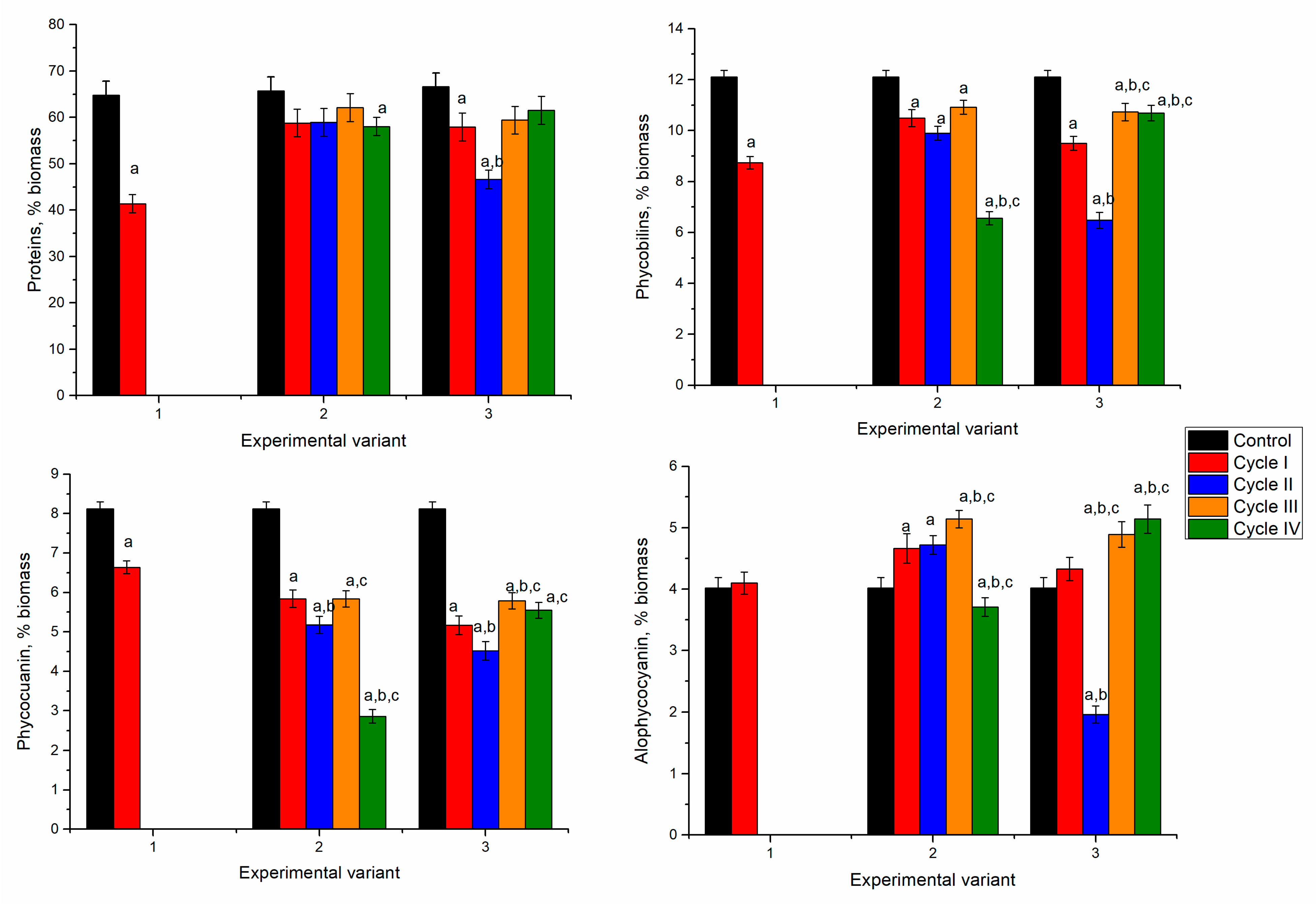
In the Ni-Cu-Zn-Mo system, in the first experimental variant, spirulina biomass survived only for one cycle (
Figure 10). At the end of the first cycle of cultivation, spirulina accumulated 40% of protein, an amount insufficient for biomass growth recovery. One of the main reasons for such an effect can be the low zinc content in the system.
As a result, the nickel content in biomass increased 5 times, the content of copper 59 times, and the quantity of molybdenum was 3.5 mg/kg. The zinc content in biomass decreased by 25% compared to control.
In the second experimental variant, even though the metal concentrations in the solution were higher, the biomass could grow for four cycles. Additionally, the solution of specified chemical composition resulted in increased copper content in biomass by a factor of 1110 in the first cycles of cultivation. In the next cycles, Cu was not detected in biomass. The same pattern was marked for molybdenum, which content was reduced from 4.5 mg/kg (first cycle of cultivation) to an undetectable level (fourth cycle). The zinc content in biomass continuously decreased over the cycles compared to control, while nickel increased (3–5/5.1 times). Adaptation to high metal concentration in cyanobacteria is manifested through several mechanisms: the ability to excrete to the media-heavy metal ligands, like siderophores or extracellular polymeric substances, the production of metallothioneins, or the induction of metal transporters [
28].
In the third experimental variant, maximum accumulation of all metal ions present in the mixture took place in the first cultivation cycle: 40 times for nickel, 2870 times for copper, 9.4 times for zinc, and 4.6 mg/kg for molybdenum. In the next three cycles, a drastic decrease in metal uptake was noticed. By the fourth cycle, copper and molybdenum were not detected in biomass, zinc content was lower by 88% than in control, while nickel was 17 times. Nickel adversity in microorganisms is mostly due to its ability to interfere with or replace other essential minerals, such as iron, zinc, copper, calcium, magnesium, molybdenum, iodine, potassium, and sodium in cells [
33]. The observed reduction of the levels of zinc, copper, and molybdenum in biomass support this fact.
As mentioned previously, in the first experimental variant, the spirulina biomass was able to grow only for one cycle, and the biomass was 12% lower (statistically insignificant) than the control (
Figure 11). In this case, a correlation between productivity and the accumulation of metals in biomass cannot be determined. In the second experimental variant, the biomass productivity was lower by 10–13% (statistically insignificant) than in control. The biomass content obtained in the third experimental variant did not appreciably change over the repeated cultivation in a metal loaded medium.
The biomass obtained in the first experimental variant at the end of the first cycle of cultivation contained 41.4% of proteins, which was 37% less compared to the control sample (
Figure 12). In the second experimental variant, the values of the protein content in biomass over the four cycles of repeated cultivation were relatively stable and slightly below the control level, and the differences were statistically insignificant. In the third experimental variant, the metal ions negatively influenced protein synthesis during two consecutive cultivation cycles. The value initially decreased by 27% but then returned to control during the third and fourth cultivation cycles.
The content of phycobiliproteins in the analyzed system changed differently. Thus, in the first experimental variant, it decreased by 12%. In the second experimental variant, a continuous decrease of phycobiliproteins took place, except in the third cycle, in which the content slightly increased compared to the first and second cycles. The most pronounced decrease was in the fourth cycle, 39% compared to control. In the third experimental variant, the decrease of phycobiliproteins in the first two cycles was accompanied by an increase in the next two cycles. The change in the content of phycocyanin followed the same trend as the case of phycobiliproteins. The most pronounced decrease was noticed in the fourth cycle of the second experimental variant, by 40% compared to control. The allophycocyanin content in the first and second experimental variants was on the level of control or higher (by 14–22%). In the third experimental variant, an abnormal decrease of allophycocyanin by 50% in the second cycle was observed, while in the other cycles, it was higher than control (by 7–22%).
The performed experiments demonstrated that A. platensis could be considered a candidate for wastewater treatment on an industrial scale. The biomass’s ability to grow for several cycles in metal contaminated media allows the reduction of operational costs for the process. Further studies are required to develop equipment that could provide, in automatic mode, the cultivation of A. platensis, wastewater supply, and removal of the treated water.


 ,
, 














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}