
Effects of Spore-Displayed p75 Protein from Lacticaseibacillus rhamnosus GG on the Transcriptional Response of HT-29 Cells


Abstract
:1. Introduction
2. Materials and Methods
2.1. Spore Preparation
2.2. Cell Culture and Treatment
2.3. Cell Viability Assay
2.4. Total RNA Isolation, cDNA Library Construction, and RNA-Sequencing
2.5. Identification of Differentially Expressed Genes
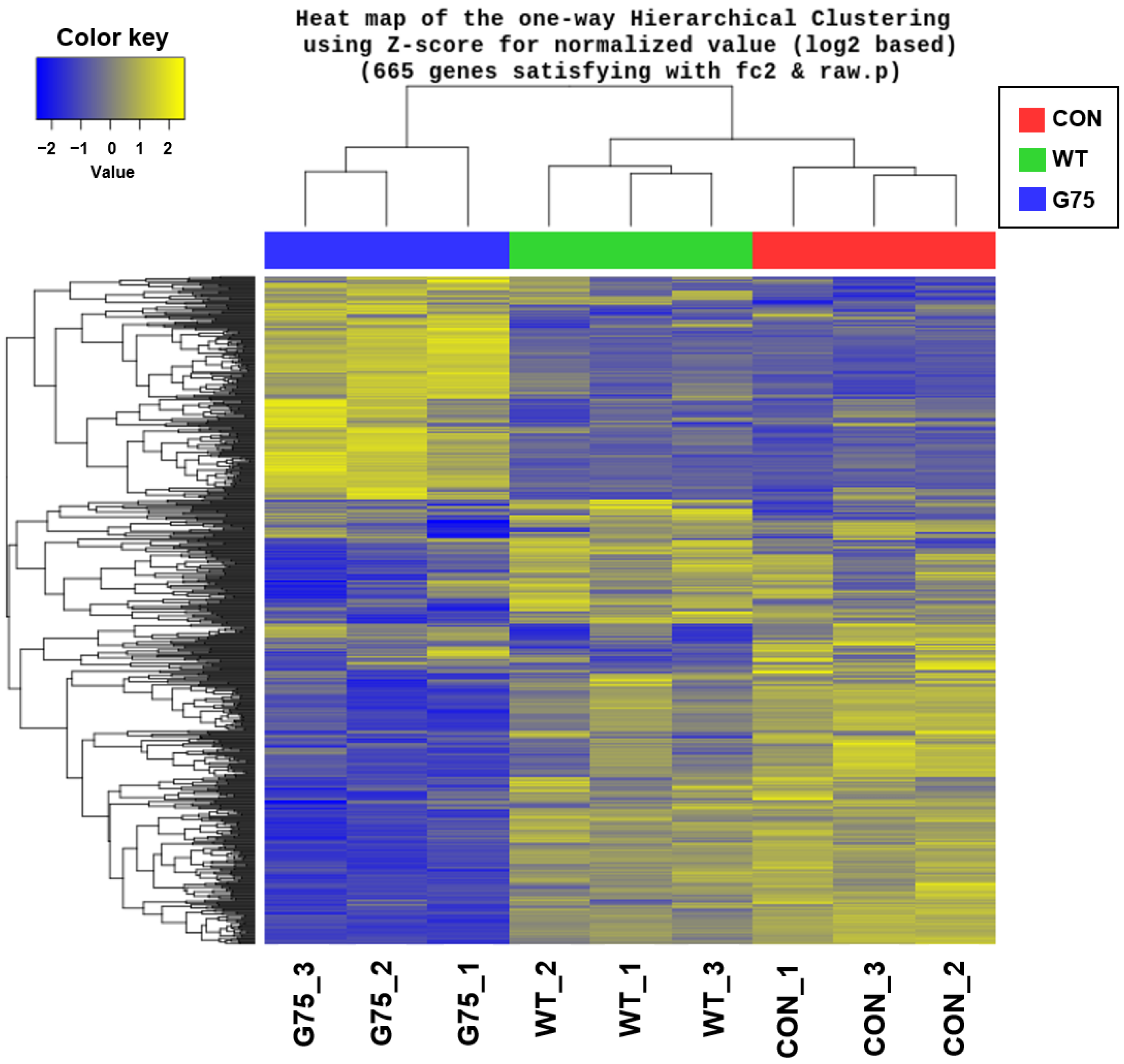
2.6. Hierarchical Clustering
2.7. Gene Ontology (GO) Enrichment Analysis and Gene–Gene Interaction Network Analysis
2.8. Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR) Confirmation
2.9. HBEGF and MUC5AC Protein Expression Verification
2.10. Statistical Analysis
3. Results
3.1. Summary of Transcriptome Sequencing Data
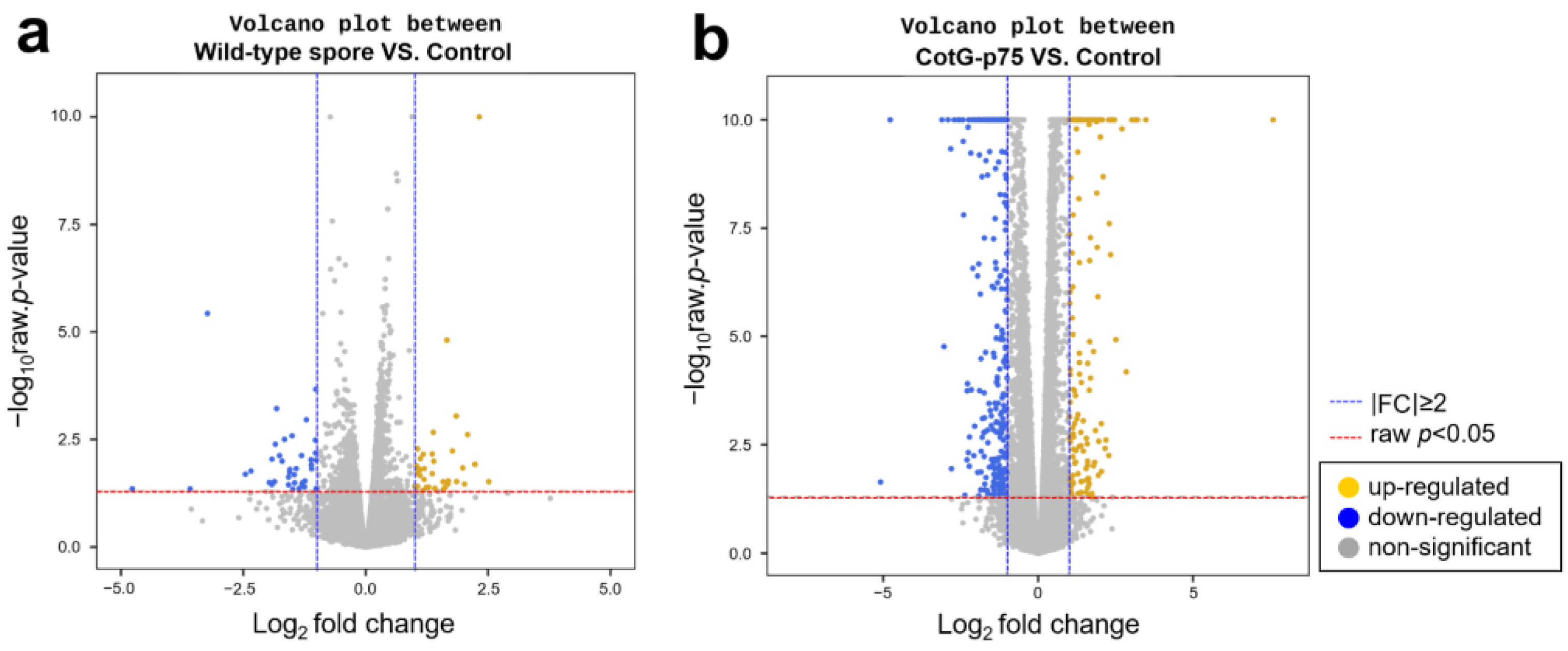
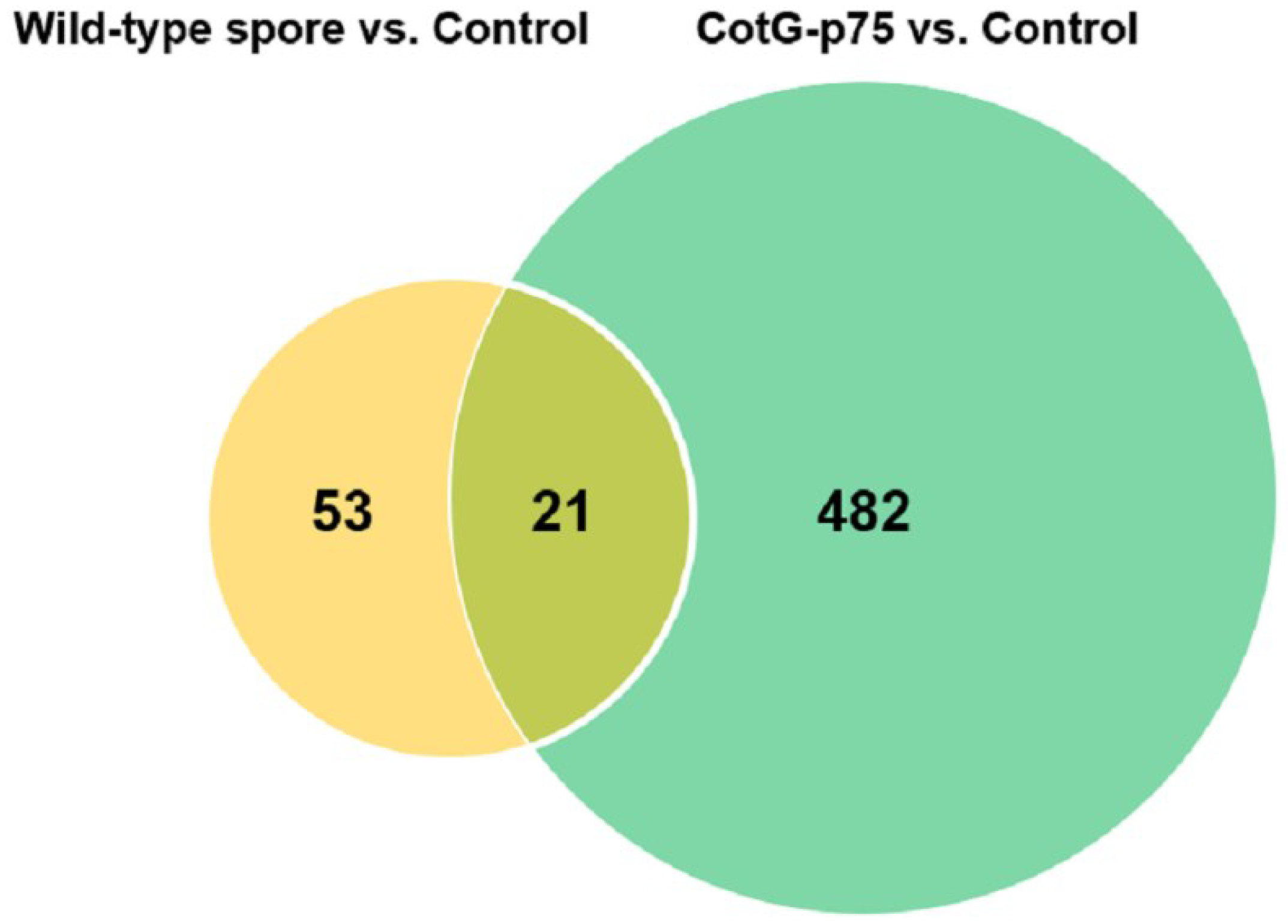
3.2. Differential Gene Expression in HT-29 Cells Stimulated with CotG-p75
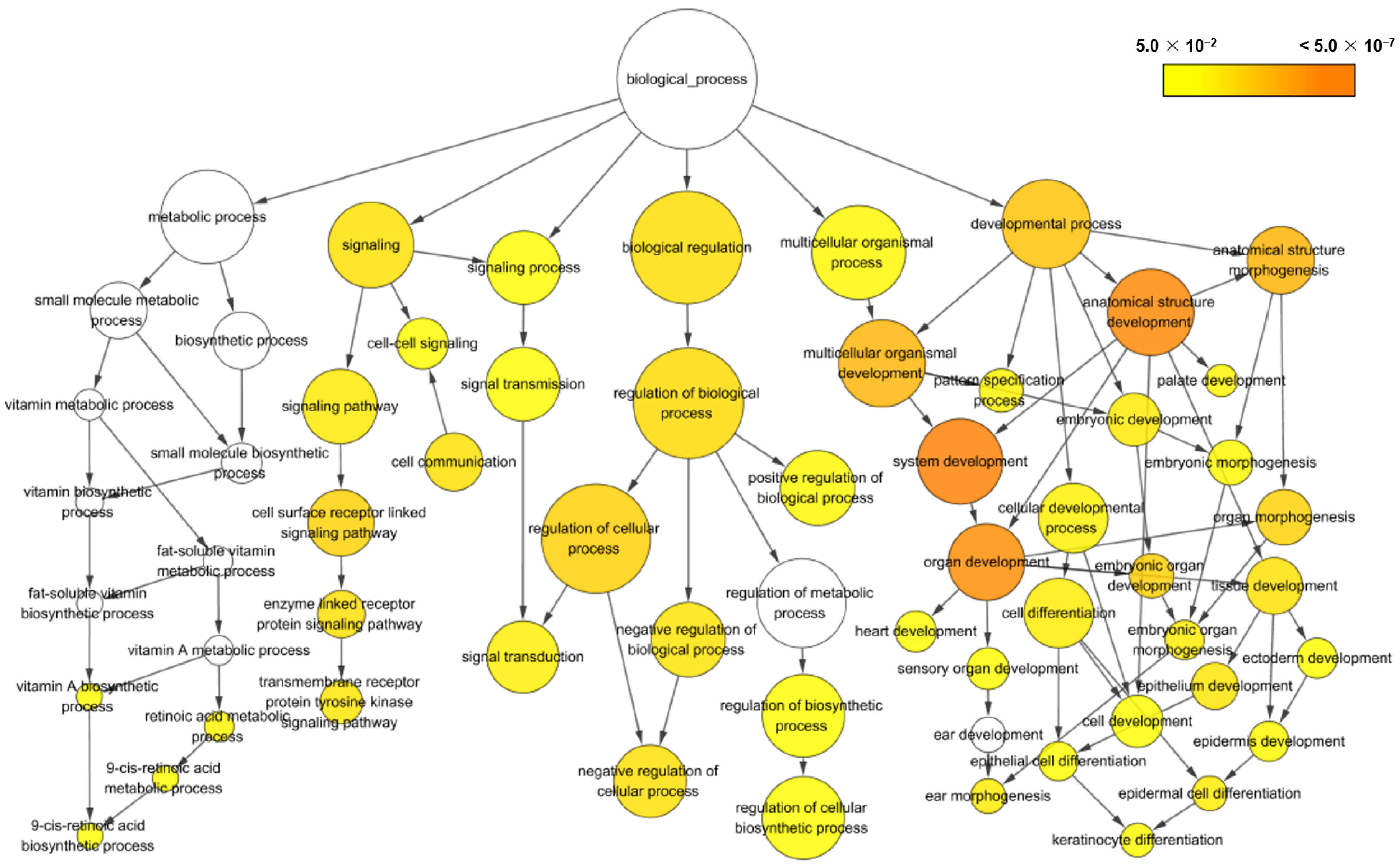
3.3. Gene Ontology (GO) Enrichment Analysis
3.4. Identification of DEGs Related to Cell Survival and Development
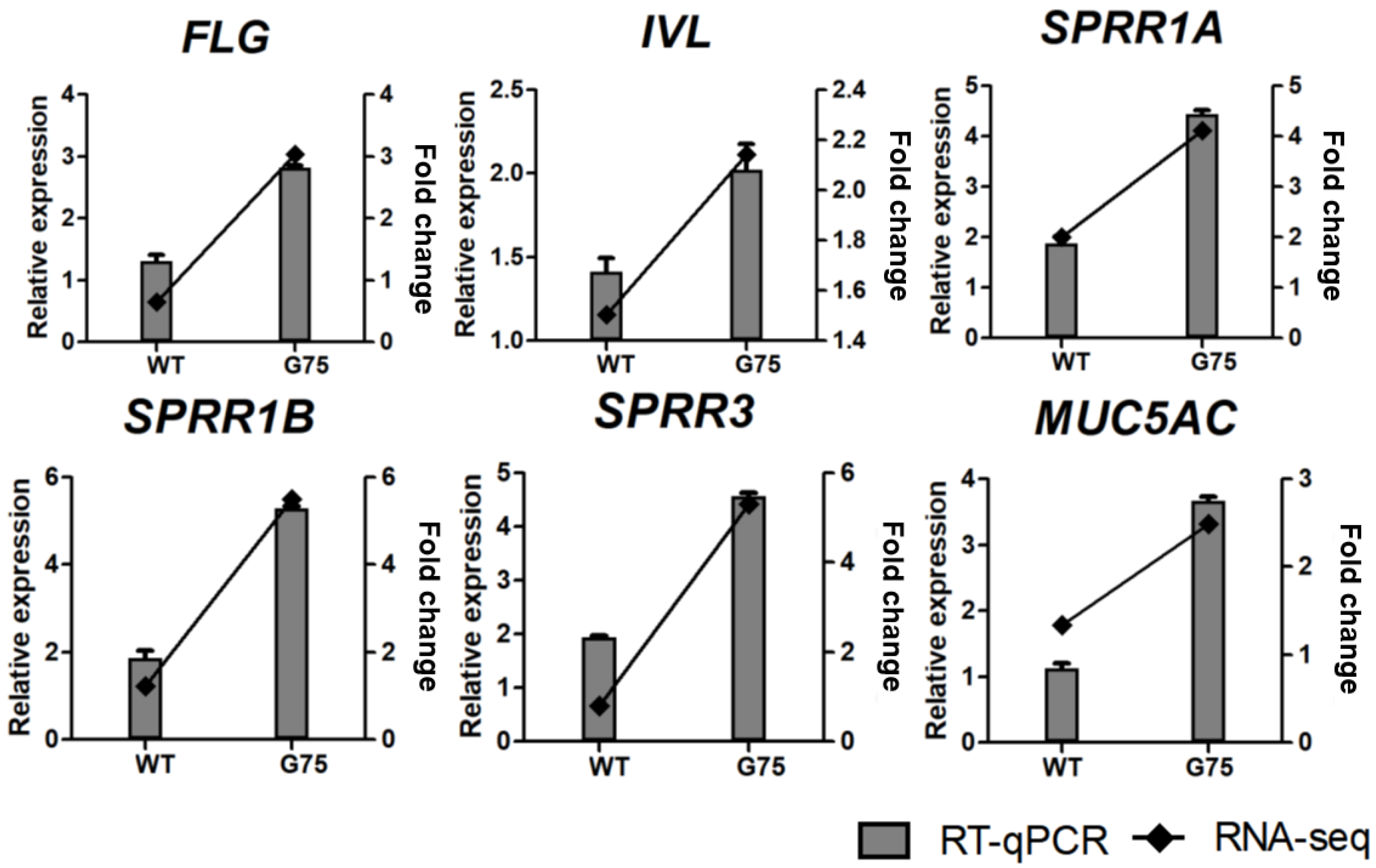
3.5. Validation of RNA-seq Data by RT-qPCR
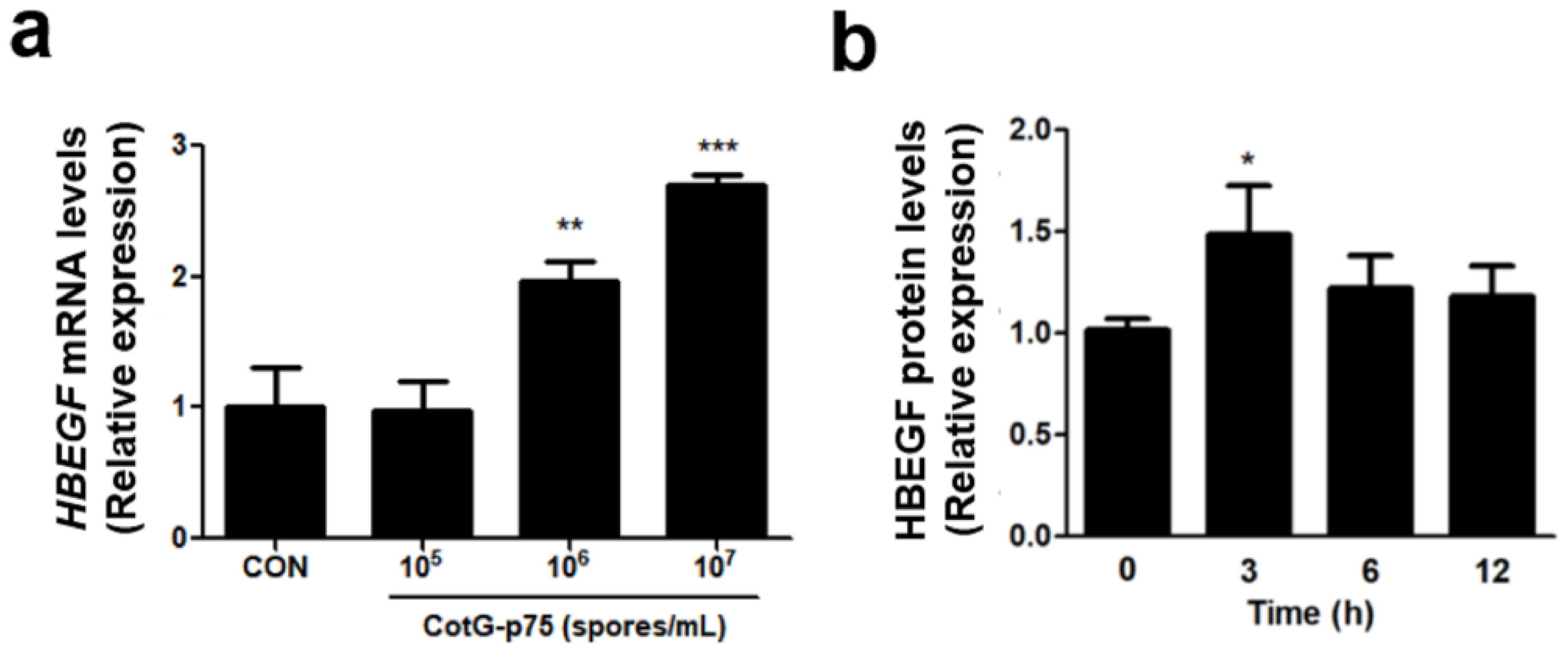
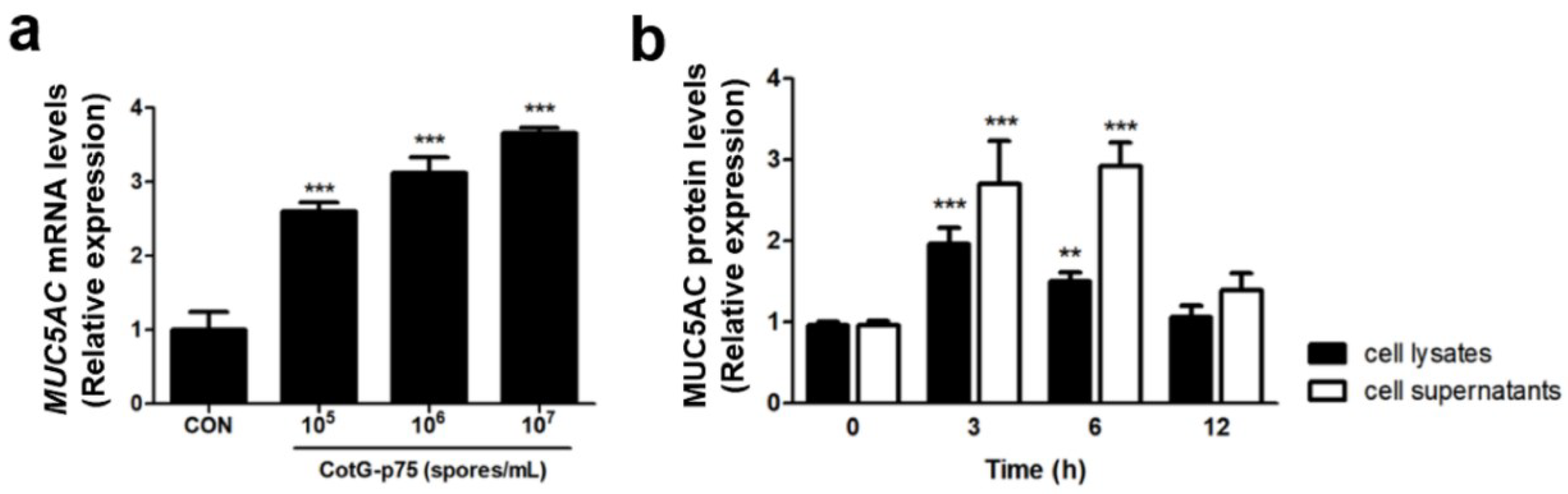
3.6. Effect of CotG-p75 on Heparin-Binding Epidermal Growth Factor (HBEGF) and MUC5AC Protein Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hotel, A.C.P.; Cordoba, A. Health and nutritional properties of probiotics in food including powder milk with live lactic acid bacteria. Prevention 2001, 5, 1–10. [Google Scholar]
- Singh, R.; Chandrashekharappa, S.; Bodduluri, S.R.; Baby, B.V.; Hegde, B.; Kotla, N.G.; Hiwale, A.A.; Saiyed, T.; Patel, P.; Vijay-Kumar, M. Enhancement of the gut barrier integrity by a microbial metabolite through the Nrf2 pathway. Nat. Commun. 2019, 10, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nataraj, B.H.; Ali, S.A.; Behare, P.V.; Yadav, H. Postbiotics-parabiotics: The new horizons in microbial biotherapy and functional foods. Microb. Cell Factories 2020, 19, 168. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, B.; Bressollier, P.; Urdaci, M.C. Exported proteins in probiotic bacteria: Adhesion to intestinal surfaces, host immunomodulation and molecular cross-talking with the host. FEMS Immunol. Med. Microbiol. 2008, 54, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, B.; Urdaci, M.C. Extracellular proteins from Lactobacillus plantarum BMCM12 prevent adhesion of enteropathogens to mucin. Curr. Microbiol. 2012, 64, 592–596. [Google Scholar] [CrossRef]
- Yan, F.; Cao, H.; Cover, T.L.; Washington, M.K.; Shi, Y.; Liu, L.; Chaturvedi, R.; Peek, R.M.; Wilson, K.T.; Polk, D.B. Colon-specific delivery of a probiotic-derived soluble protein ameliorates intestinal inflammation in mice through an EGFR-dependent mechanism. J. Clin. Investig. 2011, 121, 2242–2253. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, L.; Moore, D.J.; Shen, X.; Peek, R.; Acra, S.A.; Li, H.; Ren, X.; Polk, D.B.; Yan, F. An LGG-derived protein promotes IgA production through upregulation of APRIL expression in intestinal epithelial cells. Mucosal Immunol. 2017, 10, 373–384. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Polk, D.B. Lactobacillus rhamnosus GG: An updated strategy to use microbial products to promote health. Funct. Food Rev. 2012, 4, 77–84. [Google Scholar]
- Delgado, S.; Sánchez, B.; Margolles, A.; Ruas-Madiedo, P.; Ruiz, L. Molecules produced by probiotics and intestinal microorganisms with immunomodulatory activity. Nutrients 2020, 12, 391. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yu, Z.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Surface components and metabolites of probiotics for regulation of intestinal epithelial barrier. Microb. Cell Factories 2020, 19, 23. [Google Scholar] [CrossRef]
- Yan, F.; Polk, D.B. Probiotic bacterium prevents cytokine-induced apoptosis in intestinal epithelial cells. J. Biol. Chem. 2002, 277, 50959–50965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, F.; Cao, H.; Cover, T.L.; Whitehead, R.; Washington, M.K.; Polk, D.B. Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology 2007, 132, 562–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bäuerl, C.; Pérez-Martínez, G.; Yan, F.; Polk, D.B.; Monedero, V. Functional analysis of the p40 and p75 proteins from Lactobacillus casei BL23. Microb. Physiol. 2010, 19, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Bäuerl, C.; Abitayeva, G.; Sosa-Carrillo, S.; Mencher-Beltrán, A.; Navarro-Lleó, N.; Coll-Marqués, J.M.; Zúñiga-Cabrera, M.; Shaikhin, S.; Pérez-Martinez, G. P40 and P75 are singular functional muramidases present in the Lactobacillus casei/paracasei/rhamnosus taxon. Front. Microbiol. 2019, 10, 1420. [Google Scholar] [CrossRef] [PubMed]
- Seth, A.; Yan, F.; Polk, D.B.; Rao, R. Probiotics ameliorate the hydrogen peroxide-induced epithelial barrier disruption by a PKC- and MAP kinase-dependent mechanism. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1060–G1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Cao, H.; Liu, L.; Wang, B.; Walker, W.A.; Acra, S.A.; Yan, F. Activation of epidermal growth factor receptor mediates mucin production stimulated by p40, a Lactobacillus rhamnosus GG-derived protein. J. Biol. Chem. 2014, 289, 20234–20244. [Google Scholar] [CrossRef] [Green Version]
- Isticato, R.; Cangiano, G.; Tran, H.T.; Ciabattini, A.; Medaglini, D.; Oggioni, M.R.; De Felice, M.; Pozzi, G.; Ricca, E. Surface display of recombinant proteins on Bacillus subtilis spores. J. Bacteriol. 2001, 183, 6294–6301. [Google Scholar] [CrossRef] [Green Version]
- Henriques, A.O.; Moran, J.; Charles, P. Structure, assembly, and function of the spore surface layers. Annu. Rev. Microbiol. 2007, 61, 555–588. [Google Scholar] [CrossRef]
- Jadamus, A.; Vahjen, W.; Simon, O. Growth behaviour of a spore forming probiotic strain in the gastrointestinal tract of broiler chicken and piglets. Arch. Anim. Nutr. 2001, 54, 1–17. [Google Scholar] [CrossRef]
- Hong, H.A.; Duc, L.H.; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef] [Green Version]
- Hoa, N.T.; Baccigalupi, L.; Huxham, A.; Smertenko, A.; Van, P.H.; Ammendola, S.; Ricca, E.; Cutting, S.M. Characterization of Bacillus species used for oral bacteriotherapy and bacterioprophylaxis of gastrointestinal disorders. Appl. Environ. Microbiol. 2000, 66, 5241–5247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cutting, S.M. Bacillus probiotics. Food Microbiol. 2011, 28, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Guoyan, Z.; Yingfeng, A.; Zabed, H.M.; Qi, G.; Yang, M.; Jiao, Y.; Li, W.; Wenjing, S.; Xianghui, Q. Bacillus subtilis spore surface display technology: A review of its development and applications. J. Microbiol. Biotechnol. 2019, 29, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; An, Y.; Yun, J.; Yang, M.; Magocha, T.A.; Zhu, J.; Xue, Y.; Qi, Y.; Hossain, Z.; Sun, W. Enhanced D-tagatose production by spore surface-displayed L-arabinose isomerase from isolated Lactobacillus brevis PC16 and biotransformation. Bioresour. Technol. 2018, 247, 940–946. [Google Scholar] [CrossRef] [PubMed]
- Hinc, K.; Isticato, R.; Dembek, M.; Karczewska, J.; Iwanicki, A.; Peszyńska-Sularz, G.; De Felice, M.; Obuchowski, M.; Ricca, E. Expression and display of UreA of Helicobacter acinonychis on the surface of Bacillus subtilis spores. Microb. Cell Factories 2010, 9, 2. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Yang, R.; Hua, X.; Zhang, W.; Zhao, W. An approach for lactulose production using the CotX-mediated spore-displayed β-galactosidase as a biocatalyst. J. Microbiol. Biotechnol. 2016, 26, 1267–1277. [Google Scholar] [CrossRef]
- Das, K.; Thomas, T.; Garnica, O.; Dhandayuthapani, S. Recombinant Bacillus subtilis spores for the delivery of Mycobacterium tuberculosis Ag85B-CFP10 secretory antigens. Tuberculosis 2016, 101, S18–S27. [Google Scholar] [CrossRef]
- Dai, X.; Liu, M.; Pan, K.; Yang, J. Surface display of OmpC of Salmonella serovar Pullorum on Bacillus subtilis spores. PLoS ONE 2018, 13, e0191627. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Feng, J.; Li, J.; Li, J.; Wang, Z.; Khalique, A.; Yang, M.; Ni, X.; Zeng, D.; Zhang, D. Surface display of antigen protein VP8* of porcine rotavirus on Bacillus Subtilis spores using CotB as a fusion partner. Molecules 2019, 24, 3793. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.; Feng, F.; Ge, Q.; Zhu, F.; Chen, L.; Lv, P.; Ma, S.; Yao, Q.; Chen, K. Display of quintuple glucagon-like peptide 1 (28–36) nonapeptide on Bacillus subtilis spore for oral administration in the treatment of type 2 diabetes. J. Appl. Microbiol. 2021, 130, 314–324. [Google Scholar] [CrossRef]
- Kang, S.J.; Jun, J.S.; Moon, J.A.; Hong, K.W. Surface display of p75, a Lactobacillus rhamnosus GG derived protein, on Bacillus subtilis spores and its antibacterial activity against Listeria monocytogenes. AMB Express 2020, 10, 139. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maere, S.; Heymans, K.; Kuiper, M. BiNGO: A Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinformatics 2005, 21, 3448–3449. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Von Mering, C.; Jensen, L.J.; Snel, B.; Hooper, S.D.; Krupp, M.; Foglierini, M.; Jouffre, N.; Huynen, M.A.; Bork, P. STRING: Known and predicted protein–protein associations, integrated and transferred across organisms. Nucleic Acids Res. 2005, 33, D433–D437. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Croft, M.; Siegel, R.M. Beyond TNF: TNF superfamily cytokines as targets for the treatment of rheumatic diseases. Nat. Rev. Rheumatol. 2017, 13, 217–233. [Google Scholar] [CrossRef] [Green Version]
- Strater, J.; Wellisch, I.; Riedl, S.; Walczak, H.; Koretz, K.; Tandara, A.; Krammer, P.H.; Moller, P. CD95 (APO-1/Fas)-mediated apoptosis in colon epithelial cells: A possible role in ulcerative colitis. Gastroenterology 1997, 113, 160–167. [Google Scholar] [CrossRef]
- Janssen, E.M.; Droin, N.M.; Lemmens, E.E.; Pinkoski, M.J.; Bensinger, S.J.; Ehst, B.D.; Griffith, T.S.; Green, D.R.; Schoenberger, S.P. CD4+ T-cell help controls CD8+ T-cell memory via TRAIL-mediated activation-induced cell death. Nature 2005, 434, 88–93. [Google Scholar] [CrossRef]
- Begue, B.; Wajant, H.; Bambou, J.C.; Dubuquoy, L.; Siegmund, D.; Beaulieu, J.F.; Canioni, D.; Berrebi, D.; Brousse, N.; Desreumaux, P. Implication of TNF-related apoptosis-inducing ligand in inflammatory intestinal epithelial lesions. Gastroenterology 2006, 130, 1962–1974. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.R.; Thiele, D.L. Enhancement of MHC class I-stimulated alloresponses by TNF/TNF receptor (TNFR) 1 interactions and of MHC class II-stimulated alloresponses by TNF/TNFR2 interactions. Eur. J. Immunol. 2000, 30, 2900–2907. [Google Scholar] [CrossRef]
- Hamano, R.; Huang, J.; Yoshimura, T.; Oppenheim, J.J.; Chen, X. TNF optimally activatives regulatory T cells by inducing TNF receptor superfamily members TNFR2, 4-1BB and OX40. Eur. J. Immunol. 2011, 41, 2010–2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yarden, Y. The EGFR family and its ligands in human cancer: Signalling mechanisms and therapeutic opportunities. Eur. J. Cancer 2001, 37, 3–8. [Google Scholar] [CrossRef]
- Yarden, Y.; Sliwkowski, M.X. Untangling the ErbB signalling network. Nat. Rev. Mol. Cell Biol. 2001, 2, 127–137. [Google Scholar] [CrossRef]
- Dubé, P.E.; Yan, F.; Punit, S.; Girish, N.; McElroy, S.J.; Washington, M.K.; Polk, D.B. Epidermal growth factor receptor inhibits colitis-associated cancer in mice. J. Clin. Investig. 2012, 122, 2780–2792. [Google Scholar] [CrossRef]
- Nadel, J.A. Role of epidermal growth factor receptor activation in regulating mucin synthesis. Respir. Res. 2001, 2, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Liu, L.; Dempsey, P.J.; Tsai, Y.-H.; Raines, E.W.; Wilson, C.L.; Cao, H.; Cao, Z.; Liu, L.; Polk, D.B. A Lactobacillus rhamnosus GG-derived soluble protein, p40, stimulates ligand release from intestinal epithelial cells to transactivate epidermal growth factor receptor. J. Biol. Chem. 2013, 288, 30742–30751. [Google Scholar] [CrossRef] [Green Version]
- Marchiando, A.M.; Graham, W.V.; Turner, J.R. Epithelial barriers in homeostasis and disease. Annu. Rev. Pathol. Mech. Dis. 2010, 5, 119–144. [Google Scholar] [CrossRef]
- Kalinin, A.; Marekov, L.N.; Steinert, P.M. Assembly of the epidermal cornified cell envelope. J. Cell Sci. 2001, 114, 3069–3070. [Google Scholar] [CrossRef]
- Kypriotou, M.; Huber, M.; Hohl, D. The human epidermal differentiation complex: Cornified envelope precursors, S100 proteins and the ‘fused genes’ family. Exp. Dermatol. 2012, 21, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Furue, M. Regulation of filaggrin, loricrin, and involucrin by IL-4, IL-13, IL-17A, IL-22, AHR, and NRF2: Pathogenic implications in atopic dermatitis. Int. J. Mol. Sci. 2020, 21, 5382. [Google Scholar] [CrossRef]
- Kezic, S.; Jakasa, I. Filaggrin and skin barrier function. Ski. Barrier Funct. 2016, 49, 1–7. [Google Scholar]
- Thyssen, A.P.; Thyssen, E.P. Filaggrin Dysfunction and Its Association with Inflammatory Conditions of the GI Tract. In Filaggrin; Springer: Berlin/Heidelberg, Germany, 2014; pp. 327–331. [Google Scholar]
- Carregaro, F.; Stefanini, A.C.B.; Henrique, T.; Tajara, E.H. Study of small proline-rich proteins (SPRRs) in health and disease: A review of the literature. Arch. Dermatol. Res. 2013, 305, 857–866. [Google Scholar] [CrossRef] [PubMed]
- Rinnerthaler, M.; Duschl, J.; Steinbacher, P.; Salzmann, M.; Bischof, J.; Schuller, M.; Wimmer, H.; Peer, T.; Bauer, J.W.; Richter, K. Age-related changes in the composition of the cornified envelope in human skin. Exp. Dermatol. 2013, 22, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Wong, M.H.; Thelin, A.; Hansson, L.; Falk, P.G.; Gordon, J.I. Molecular analysis of commensal host-microbial relationships in the intestine. Science 2001, 291, 881–884. [Google Scholar] [CrossRef] [Green Version]
- Hooper, L.V.; Gordon, J.I. Commensal host-bacterial relationships in the gut. Science 2001, 292, 1115–1118. [Google Scholar] [CrossRef]
- Knight, P.A.; Pemberton, A.D.; Robertson, K.A.; Roy, D.J.; Wright, S.H.; Miller, H.R. Expression profiling reveals novel innate and inflammatory responses in the jejunal epithelial compartment during infection with Trichinella spiralis. Infect. Immun. 2004, 72, 6076–6086. [Google Scholar] [CrossRef] [Green Version]
- Stern, L.E.; Erwin, C.R.; Falcone, R.A., Jr.; Huang, F.S.; Kemp, C.J.; Williams, J.L.; Warner, B.W. cDNA microarray analysis of adapting bowel after intestinal resection. J. Pediatr. Surg. 2001, 36, 190–195. [Google Scholar] [CrossRef]
- Bergstrom, K.S.; Xia, L. Mucin-type O-glycans and their roles in intestinal homeostasis. Glycobiology 2013, 23, 1026–1037. [Google Scholar] [CrossRef]
- Johansson, M.E.; Hansson, G.C. Immunological aspects of intestinal mucus and mucins. Nat. Rev. Immunol. 2016, 16, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Forstner, J.; Forstner, G. Gastrointestinal mucus. In Physiology of the Gastrointestinal Tract; Johnson, L.R., Ed.; Raven Press: New York, NY, USA, 1994. [Google Scholar]
- Mack, D.R.; Michail, S.; Wei, S.; McDougall, L.; Hollingsworth, M.A. Probiotics inhibit enteropathogenic E. coli adherence in vitro by inducing intestinal mucin gene expression. Am. J. Physiol. Gastrointest. Liver Physiol. 1999, 276, G941–G950. [Google Scholar] [CrossRef] [PubMed]
- Mattar, A.; Teitelbaum, D.H.; Drongowski, R.; Yongyi, F.; Harmon, C.; Coran, A. Probiotics up-regulate MUC-2 mucin gene expression in a Caco-2 cell-culture model. Pediatr. Surg. Int. 2002, 18, 586–590. [Google Scholar]
- Mack, D.R.; Ahrne, S.; Hyde, L.; Wei, S.; Hollingsworth, M.A. Extracellular MUC3 mucin secretion follows adherence of Lactobacillus strains to intestinal epithelial cells in vitro. Gut 2003, 52, 827–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GO ID | GO Description | Adjusted p-Value a | Cluster Frequency b | Total Frequency c |
|---|---|---|---|---|
| GO:0048731 | system development | 2.04 × 10−8 | 103/387 (26.6%) | 2421/17,777 (13.6%) |
| GO:0048856 | anatomical structure development | 2.14 × 10−7 | 106/387 (27.3%) | 2655/17,777 (14.9%) |
| GO:0048513 | organ development | 3.23 × 10−7 | 80/387 (20.6%) | 1791/17,777 (10.0%) |
| GO:0009653 | anatomical structure morphogenesis | 1.12 × 10−5 | 58/387 (14.9%) | 1217/17,777 (6.8%) |
| GO:0007275 | multicellular organismal development | 1.16 × 10−5 | 108/387 (27.9%) | 2970/17,777 (16.7%) |
| GO:0032502 | developmental process | 3.53 × 10−5 | 113/387 (29.1%) | 3233/17,777 (18.1%) |
| GO:0048568 | embryonic organ development | 1.27 × 10−4 | 19/387 (4.9%) | 215/17,777 (1.2%) |
| GO:0050794 | regulation of cellular process | 1.35 × 10−4 | 183/387 (47.2%) | 6218/17,777 (34.9%) |
| GO:0007166 | cell surface receptor linked signaling pathway | 1.55 × 10−4 | 56/387 (14.4%) | 1279/17,777 (7.1%) |
| GO:0009887 | organ morphogenesis | 1.62 × 10−4 | 35/387 (9.0%) | 634/17,777 (3.5%) |
| GO:0050789 | regulation of biological process | 3.76 × 10−4 | 188/387 (48.5%) | 6548/17,777 (36.8%) |
| GO:0065007 | biological regulation | 5.21 × 10−4 | 196/387 (50.6%) | 6937/17,777 (39.0%) |
| GO:0007167 | enzyme linked receptor protein signaling pathway | 6.06 × 10−4 | 23/387 (5.9%) | 346/17,777 (1.9%) |
| GO:0048523 | negative regulation of cellular process | 6.13 × 10−4 | 70/387 (18.0%) | 1844/17,777 (10.3%) |
| GO:0023052 | signaling | 6.46 × 10−4 | 104/387 (26.8%) | 3130/17,777 (17.6%) |
| GO:0009888 | tissue development | 6.46 × 10−4 | 37/387 (9.5%) | 748/17,777 (4.2%) |
| GO:0007154 | cell communication | 6.58 × 10−4 | 39/387 (10.0%) | 812/17,777 (4.5%) |
| GO:0048519 | negative regulation of biological process | 8.37 × 10−4 | 74/387 (19.1%) | 2019/17,777 (11.3%) |
| GO:0060429 | epithelium development | 8.69 × 10−4 | 22/387 (5.6%) | 336/17,777 (1.8%) |
| GO:0007169 | transmembrane receptor protein tyrosine kinase signaling pathway | 8.69 × 10−4 | 60/387 (15.5%) | 1526/17,777 (8.5%) |
| GO:0009790 | embryonic development | 1.06 × 10−3 | 17/387 (4.3%) | 219/17,777 (1.2%) |
| GO:0023033 | signaling pathway | 1.39 × 10−3 | 31/387 (8.0%) | 602/17,777 (3.3%) |
| GO:0030154 | cell differentiation | 1.39 × 10−3 | 75/387 (19.3%) | 2099/17,777 (11.8%) |
| GO:0048562 | embryonic organ morphogenesis | 1.47 × 10−3 | 63/387 (16.2%) | 1668/17,777 (9.3%) |
| GO:0042471 | ear morphogenesis | 1.64 × 10−3 | 13/387 (3.3%) | 140/17,777 (0.7%) |
| GO:0009913 | epidermal cell differentiation | 1.64 × 10−3 | 9/387 (2.3%) | 66/17,777 (0.3%) |
| GO:0007165 | signal transduction | 2.02 × 10−3 | 9/387 (2.3%) | 68/17,777 (0.3%) |
| GO:0048869 | cellular developmental process | 2.09 × 10−3 | 68/387 (17.5%) | 1877/17,777 (10.5%) |
| GO:0007389 | pattern specification process | 2.78 × 10−3 | 63/387 (16.2%) | 1714/17,777 (9.6%) |
| GO:0042573 | retinoic acid metabolic process | 2.78 × 10−3 | 60/387 (15.5%) | 1607/17,777 (9.0%) |
| GO:0060021 | palate development | 3.25 × 10−3 | 18/387 (4.6%) | 271/17,777 (1.5%) |
| GO:0048598 | embryonic morphogenesis | 3.47 × 10−3 | 5/387 (1.2%) | 18/17,777 (0.1%) |
| GO:0048518 | positive regulation of biological process | 3.75 × 10−3 | 7/387 (1.8%) | 43/17,777 (0.2%) |
| GO:0032501 | multicellular organismal process | 5.09 × 10−3 | 20/387 (5.1%) | 336/17,777 (1.8%) |
| GO:0008544 | epidermis development | 5.09 × 10−3 | 75/387 (19.3%) | 2207/17,777 (12.4%) |
| GO:0030216 | keratinocyte differentiation | 5.50 × 10−3 | 129/387 (33.3%) | 4374/17,777 (24.6%) |
| GO:0030855 | epithelial cell differentiation | 5.50 × 10−3 | 14/387 (3.6%) | 186/17,777 (1.0%) |
| GO:0031326 | regulation of cellular biosynthetic process | 5.78 × 10−3 | 8/387 (2.0%) | 63/17,777 (0.3%) |
| GO:0007267 | cell–cell signaling | 6.60 × 10−3 | 17/387 (4.3%) | 265/17,777 (1.4%) |
| GO:0042904 | 9-cis-retinoic acid biosynthetic process | 7.01 × 10−3 | 13/387 (3.3%) | 168/17,777 (0.9%) |
| GO:0042905 | 9-cis-retinoic acid metabolic process | 7.10 × 10−3 | 95/387 (24.5%) | 3021/17,777 (16.9%) |
| GO:0035238 | vitamin A biosynthetic process | 7.10 × 10−3 | 8/387 (2.0%) | 66/17,777 (0.3%) |
| GO:0007507 | heart development | 7.10 × 10−3 | 28/387 (7.2%) | 587/17,777 (3.3%) |
| GO:0007423 | sensory organ development | 7.10 × 10−3 | 3/387 (0.7%) | 5/17,777 (0.0%) |
| GO:0009889 | regulation of biosynthetic process | 7.10 × 10−3 | 3/387 (0.7%) | 5/17,777 (0.0%) |
| GO:0023046 | signaling process | 7.10 × 10−3 | 3/387 (0.7%) | 5/17,777 (0.0%) |
| GO:0023060 | signal transmission | 7.10 × 10−3 | 3/387 (0.7%) | 5/17,777 (0.0%) |
| GO:0048468 | cell development | 7.10 × 10−3 | 3/387 (0.7%) | 5/17,777 (0.0%) |
| GO:0007398 | ectoderm development | 7.40 × 10−3 | 15/387 (3.8%) | 221/17,777 (1.2%) |
| Gene Symbol | Gene Name | Fold Change | p-Value |
|---|---|---|---|
| Epithelial Cell Differentiation and Development | |||
| ALDH1A3 | aldehyde dehydrogenase 1 family member A3 | 8.154 | 0.000 |
| GLI2 | GLI family zinc finger 2 | 5.524 | 0.000 |
| SPRR1B | small proline rich protein 1B | 5.493 | 0.000 |
| DHRS9 | dehydrogenase/reductase 9 | 5.330 | 0.000 |
| SPRR3 | small proline rich protein 3 | 5.304 | 0.000 |
| SPRR1A | small proline rich protein 1A | 4.118 | 0.001 |
| EREG | epiregulin | 3.221 | 0.000 |
| FLG | filaggrin | 3.037 | 0.031 |
| IVL | involucrin | 2.141 | 0.000 |
| WNT7B | Wnt family member 7B | 2.134 | 0.000 |
| SRF | serum response factor | 2.124 | 0.046 |
| MYC | MYC proto-oncogene, bHLH transcription factor | 2.120 | 0.000 |
| HOXA5 | homeobox A5 | −2.043 | 0.000 |
| FZD2 | frizzled class receptor 2 | −2.697 | 0.022 |
| PTCH1 | patched 1 | −2.987 | 0.000 |
| POU3F1 | POU class 3 homeobox 1 | −3.769 | 0.000 |
| Cell proliferation | |||
| GLI2 | GLI family zinc finger 2 | 5.524 | 0.000 |
| FASLG | Fas ligand | 4.559 | 0.002 |
| EPGN | epithelial mitogen | 4.132 | 0.013 |
| CDKN1A | cyclin dependent kinase inhibitor 1A | 3.420 | 0.000 |
| CSF2 | colony stimulating factor 2 | 3.225 | 0.000 |
| EREG | epiregulin | 3.221 | 0.000 |
| FOSL1 | FOS like 1, AP-1 transcription factor subunit | 3.063 | 0.000 |
| FLG | filaggrin | 3.037 | 0.031 |
| CXCL8 | C-X-C motif chemokine ligand 2 | 2.510 | 0.003 |
| PTGS2 | prostaglandin-endoperoxide synthase 2 | 2.424 | 0.000 |
| FGF19 | fibroblast growth factor 19 | 2.400 | 0.000 |
| MYC | N-myc downstream regulated 1 | 2.164 | 0.000 |
| SERPINE1 | serpin family E member 1 | 2.143 | 0.000 |
| HOXA5 | homeobox A5 | −2.043 | 0.000 |
| ERBB3 | erb-b2 receptor tyrosine kinase 3 | −2.140 | 0.000 |
| IRS2 | insulin receptor substrate 2 | −2.605 | 0.000 |
| FGFR2 | fibroblast growth factor receptor 2 | −2.613 | 0.000 |
| CBLB | Cbl proto-oncogene B | −3.391 | 0.000 |
| BCL6 | B-cell CLL/lymphoma 6 | −3.718 | 0.000 |
| BTG2 | BTG anti-proliferation factor 2 | −4.712 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, S.-J.; Kim, M.-J.; Son, D.-Y.; Kang, S.-S.; Hong, K.-W. Effects of Spore-Displayed p75 Protein from Lacticaseibacillus rhamnosus GG on the Transcriptional Response of HT-29 Cells. Microorganisms 2022, 10, 1276. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10071276
Kang S-J, Kim M-J, Son D-Y, Kang S-S, Hong K-W. Effects of Spore-Displayed p75 Protein from Lacticaseibacillus rhamnosus GG on the Transcriptional Response of HT-29 Cells. Microorganisms. 2022; 10(7):1276. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10071276
Chicago/Turabian StyleKang, Soo-Ji, Min-Joo Kim, Do-Yeong Son, Seok-Seong Kang, and Kwang-Won Hong. 2022. "Effects of Spore-Displayed p75 Protein from Lacticaseibacillus rhamnosus GG on the Transcriptional Response of HT-29 Cells" Microorganisms 10, no. 7: 1276. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms10071276