Into the Thermus Mobilome: Presence, Diversity and Recent Activities of Insertion Sequences Across Thermus spp.

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
3. Results
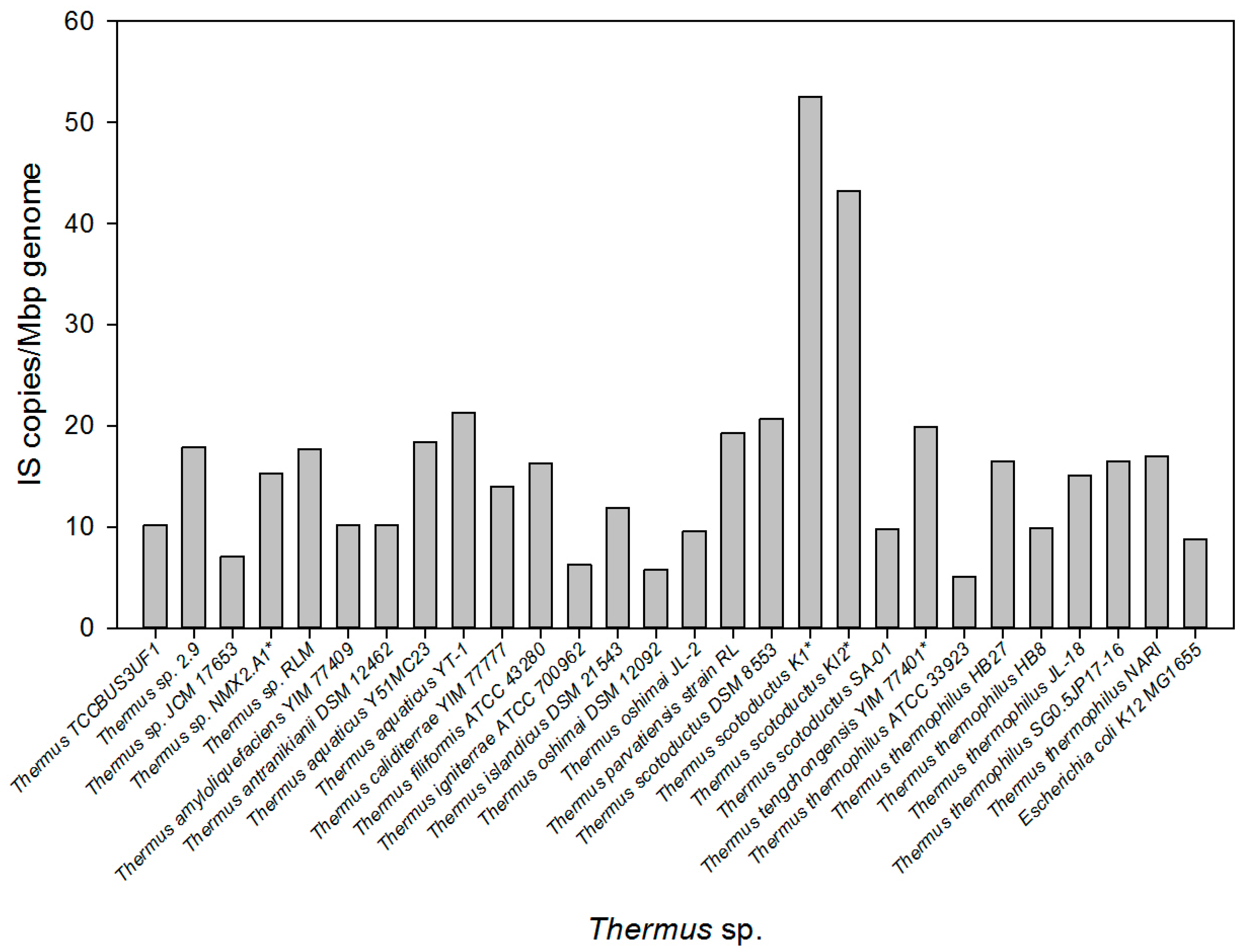
3.1. Incidence of ISs among Members of the Genus Thermus
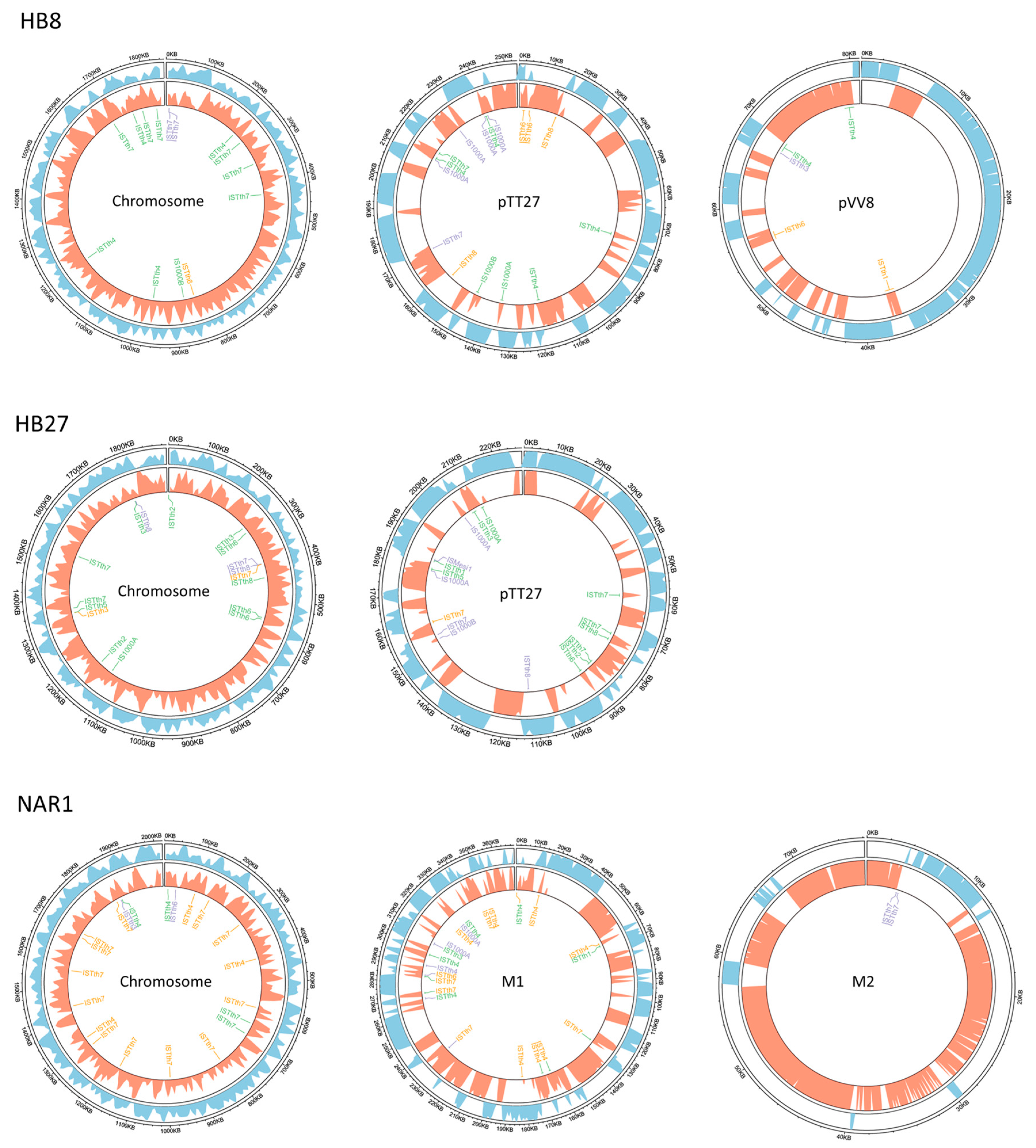
3.2. Diversity of ISs among Thermus spp. ISTth Families
3.3. Evidence of ISTth Propagation to Other Microorganisms
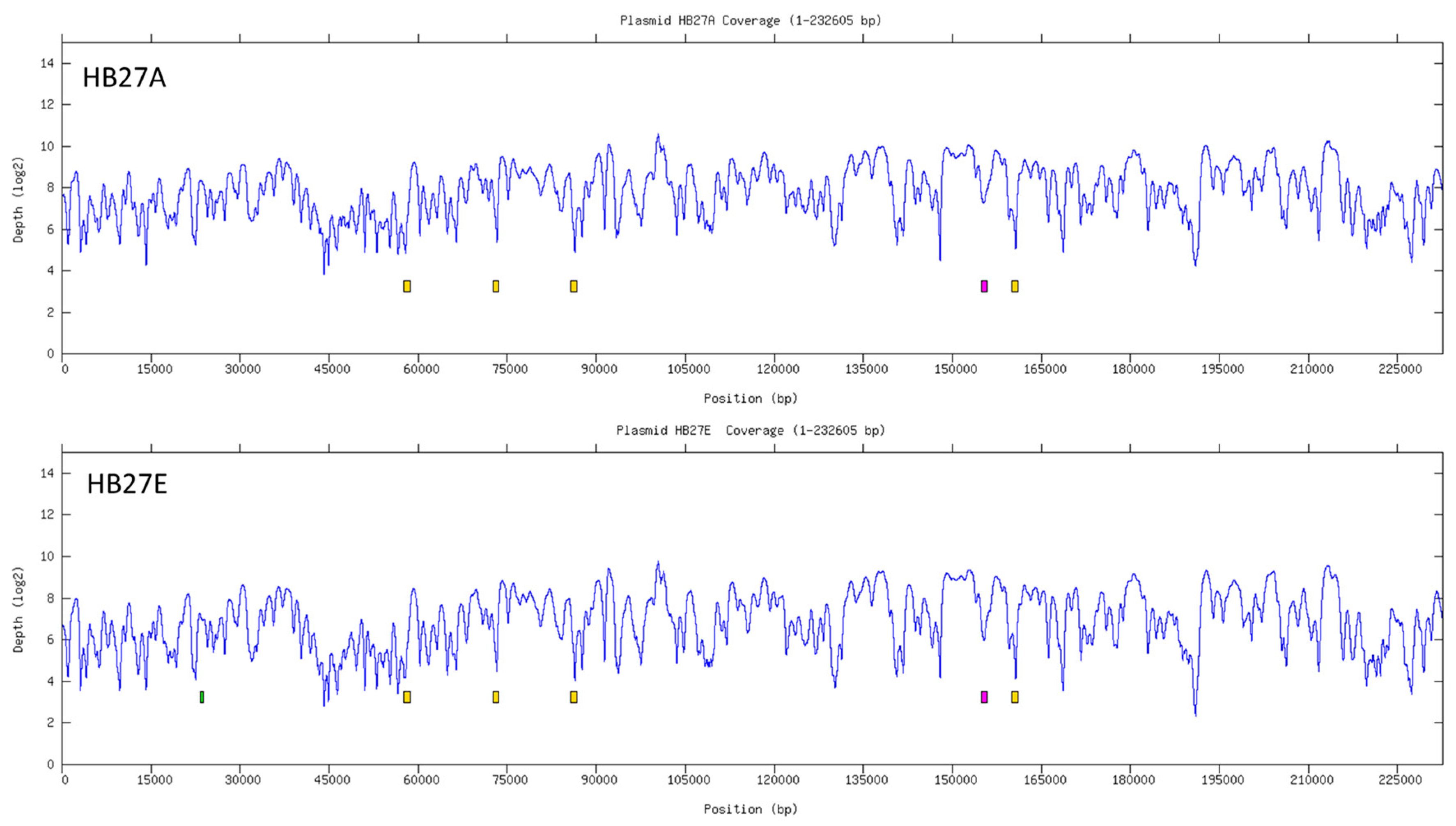
3.4. Distribution of Putatively Active vs. Inactive IS Copies across Selected T. thermophilus Genomes
3.5. Potentially Active Transposases
4. Discussion and Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gogarten, J.P.; Townsend, J.P. Horizontal gene transfer, genome innovation and evolution. Nat. Rev. Microbiol. 2005, 3, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Treangen, T.J.; Rocha, E.P.C. Horizontal Transfer, Not Duplication, Drives the Expansion of Protein Families in Prokaryotes. PLoS Genet. 2011, 7, e1001284. [Google Scholar] [CrossRef] [PubMed]
- Gogarten, J.P.; Doolittle, W.F.; Lawrence, J.G. Prokaryotic evolution in light of gene transfer. Mol. Biol. Evol. 2002, 19, 2226–2238. [Google Scholar] [CrossRef] [PubMed]
- Frost, L.S.; Leplae, R.; Summers, A.O.; Toussaint, A. Mobile genetic elements: The agents of open source evolution. Nat. Rev. Microbiol. 2005, 3, 722. [Google Scholar] [CrossRef] [PubMed]
- Lang, A.; Beatty, J.T.; Rice, P.A. Guest Editorial: Mobile Genetic Elements and Horizontal Gene Transfer in Prokaryotes; Elsevier: New York, NY, USA, 2017. [Google Scholar]
- Wozniak, R.A.; Waldor, M.K. Integrative and conjugative elements: Mosaic mobile genetic elements enabling dynamic lateral gene flow. Nat. Rev. Microbiol. 2010, 8, 552. [Google Scholar] [CrossRef]
- Shapiro, J. (Ed.) Mobile Genetic Elements; Elsevier: New York, NY, USA, 2012. [Google Scholar]
- Mahillon, J.; Chandler, M. Insertion sequences. Microbiol. Mol. Biol. Rev. 1998, 62, 725–774. [Google Scholar] [PubMed]
- Shintani, M. The behavior of mobile genetic elements (MGEs) in different environments. Biosci. Biotechnol. Biochem. 2017, 81, 854–862. [Google Scholar] [CrossRef] [PubMed]
- Van Elsas, J.D.; Bailey, M.J. The ecology of transfer of mobile genetic elements. FEMS Microbiol. Ecol. 2002, 42, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Syvanen, M. The evolutionary implications of mobile genetic elements. Annu. Rev. Genet. 1984, 18, 271–293. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef]
- Carraro, N.; Rivard, N.; Burrus, V.; Ceccarelli, D. Mobilizable genomic islands, different strategies for the dissemination of multidrug resistance and other adaptive traits. Mob. Genet. Elem. 2017, 7, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stokes, H.W.; Gillings, M.R. Gene flow, mobile genetic elements and the recruitment of antibiotic resistance genes into Gram-negative pathogens. FEMS Microbiol. Rev. 2011, 35, 790–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Top, E.M.; Springael, D. The role of mobile genetic elements in bacterial adaptation to xenobiotic organic compounds. Curr. Opin. Biotechnol. 2003, 14, 262–269. [Google Scholar] [CrossRef]
- Siguier, P.; Filée, J.; Chandler, M. Insertion sequences in prokaryotic genomes. Curr. Opin. Microbiol. 2006, 9, 526–531. [Google Scholar] [CrossRef]
- Siguier, P.; Pérochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic Acids Res. 2006, 34 (Suppl. S1), D32–D36. [Google Scholar] [CrossRef] [PubMed]
- Brochier-Armanet, C.; Forterre, P. Widespread distribution of archaeal reverse gyrase in thermophilic bacteria suggests a complex history of vertical inheritance and lateral gene transfers. Archaea 2006, 2, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Nelson, K.E.; Clayton, R.A.; Gill, S.R.; Gwinn, M.L.; Dodson, R.J.; Haft, D.H.; McDonald, L.; Utterback, T.R.; Malek, J.A.; Linher, K.D.; et al. Evidence for lateral gene transfer between Archaea and bacteria from genome sequence of Thermotoga maritima. Nature 1999, 399, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Akanuma, S.; Yokobori, S.-I.; Yamagishi, A. Comparative genomics of thermophilic bacteria and archaea. In Thermophilic Microbes in Environmental and Industrial Biotechnology; Springer: Dordrecht, The Netherlands, 2013; pp. 331–349. [Google Scholar]
- Aminov, R.I. Horizontal gene exchange in environmental microbiota. Front. Microbiol. 2011, 2, 158. [Google Scholar] [CrossRef] [PubMed]
- Lossouarn, J.; Dupont, S.; Gorlas, A.; Mercier, C.; Bienvenu, N.; Marguet, E.; Forterre, P.; Geslin, C. An abyssal mobilome: Viruses, plasmids and vesicles from deep-sea hydrothermal vents. Res. Microbiol. 2015, 166, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Krupovic, M.; Gonnet, M.; Hania, W.; Forterre, P.; Erauso, G. Insights into Dynamics of Mobile Genetic Elements in Hyperthermophilic Environments. PLoS ONE 2013, 8, e49044. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Gourbeyre, E.; Chandler, M. Bacterial insertion sequences: Their genomic impact and diversity. FEMS Microbiol. Rev. 2014, 38, 865–891. [Google Scholar] [CrossRef] [PubMed]
- Nelson, W.C.; Wollerman, L.; Bhaya, D.; Heidelberg, J.F. Analysis of insertion sequences in thermophilic cyanobacteria: Exploring the mechanisms of establishing, maintaining and withstanding high insertion sequence abundance. Appl. Environ. Microbiol. 2011, 77, 5458–5466. [Google Scholar] [CrossRef] [PubMed]
- Takami, H.; Takaki, Y.; Chee, G.J.; Nishi, S.; Shimamura, S.; Suzuki, H.; Matsui, S.; Uchiyama, I. Thermoadaptation trait revealed by the genome sequence of thermophilic Geobacillus kaustophilus. Nucleic Acids Res. 2004, 32, 6292–6303. [Google Scholar] [CrossRef] [PubMed]
- Filee, J.; Siguier, P.; Chandler, M. Insertion sequence diversity in archaea. Microbiol. Mol. Biol. Rev. 2007, 71, 121–157. [Google Scholar] [CrossRef] [PubMed]
- Omelchenko, M.V.; Wolf, Y.I.; Gaidamakova, E.K.; Matrosova, V.Y.; Vasilenko, A.; Zhai, M.; Daly, M.J.; Koonin, E.V.; Makarova, K.S. Comparative genomics of Thermus thermophilus and Deinococcus radiodurans: Divergent routes of adaptation to thermophily and radiation resistance. BMC Evol. Biol. 2005, 5, 57. [Google Scholar] [CrossRef] [PubMed]
- Madigan, M.T.; Martinko, J.M.; Parker, J. Brock Biology of Microorganisms; Prentice Hall: Upper Saddle River, NJ, USA, 1997; Volume 11. [Google Scholar]
- Blesa, A.; Averhoff, B.; Berenguer, J. Horizontal Gene Transfer in Thermus spp. Curr. Issues Mol. Biol. 2018, 29, 23–36. [Google Scholar] [CrossRef]
- Cava, F.; Hidalgo, A.; Berenguer, J. Thermus thermophilus as biological model. Extremophiles 2009, 13, 213. [Google Scholar] [CrossRef] [PubMed]
- Touchon, M.; Bobay, L.M.; Rocha, E.P. The chromosomal accommodation and domestication of mobile genetic elements. Curr. Opin. Microbiol. 2014, 22, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Ashby, M.; Bergquist, P. Cloning and sequence of IS1000, a putative insertion sequence from Thermus thermophilus HB8. Plasmid 1990, 24, 1–11. [Google Scholar] [CrossRef]
- Tabata, K.; Hoshino, T. Mapping of 61 genes on the refined physical map of the chromosome of Thermus thermophilus HB27 and comparison of genome organization with that of T. thermophilus HB8. Microbiology 1996, 142, 401–410. [Google Scholar] [CrossRef]
- Gregory, S.T.; Dahlberg, A.E. Transposition of an insertion sequence ISTth7 in the genome of the extreme thermophile Thermus thermophilus HB8. FEMS Microbiol. Lett. 2008, 289, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Cava, F.; Laptenko, O.; Borukhov, S.; Chahlafi, Z.; Blas-Galindo, E.; Gómez-Puertas, P.; Berenguer, J. Control of the respiratory metabolism of Thermus thermophilus by the nitrate respiration conjugative element NCE. Mol. Microbiol. 2007, 64, 630–646. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Arcos, S.; Fernández-Herrero, L.A.; Marín, I.; Berenguer, J. Anaerobic growth, a property horizontally transferred by an Hfr-like mechanism among extreme thermophiles. J. Bacteriol. 1998, 180, 3137–3143. [Google Scholar] [PubMed]
- Chin, C.S.; Alexander, D.H.; Marks, P.; Klammer, A.A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.E.; et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat. Methods 2013, 10, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Sommer, D.D.; Delcher, A.L.; Salzberg, S.L.; Pop, M. Minimus: A fast, lightweight genome assembler. BMC Bioinform. 2007, 8, 64. [Google Scholar] [CrossRef]
- PacBio Utilities. Available online: https://github.com/douglasgscofield/PacBio-utilities (accessed on 11 November 2018).
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. An Integrated Tool for Comprehensive Microbial Variant Detection and Genome Assembly Improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bionformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Felsenstein, J. PHYLIP (Phylogeny Inference Package) Distributed by the Author; Version 3.c; Department of Genome Sciences, University of Washington: Seattle, WA, USA, 2005. [Google Scholar]
- Touchon, M.; Rocha, E.P.C. Causes of Insertion Sequences Abundance in Prokaryotic Genomes. Mol. Biol. Evol. 2007, 24, 969–981. [Google Scholar] [CrossRef] [Green Version]
- Da Cunha, V.; Guerillot, R.; Brochet, M.; Glaser, P. Integrative and conjugative elements encoding DDE transposases. In Bacterial Integrative Mobile Genetic Elements; Roberts, A.P., Mullany, P., Eds.; Landes Bioscience: Austin, TX, USA, 2013; pp. 250–260. [Google Scholar]
- De la Cruz, F.; Davies, J. Horizontal gene transfer and the origin of species: Lessons from bacteria. Trends Microbiol. 2000, 8, 128–133. [Google Scholar] [CrossRef]
- Brüggemann, H.; Chen, C. Comparative genomics of Thermus thermophilus: Plasticity of the megaplasmid and its contribution to a thermophilic lifestyle. J. Biotechnol. 2006, 124, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, L.; Bricio, C.; Gómez, M.J.; Berenguer, J. Lateral transfer of the denitrification pathway genes among Thermus thermophilus strains. Appl. Environ. Microbiol. 2011, 77, 1352–1358. [Google Scholar] [CrossRef] [PubMed]
- Swarts, D.C.; Jore, M.M.; Westra, E.R.; Zhu, Y.; Janssen, J.H.; Snijders, A.P.; Wang, Y.; Patel, D.J.; Berenguer, J.; Brouns, S.J.J.; et al. DNA-guided DNA interference by a prokaryotic Argonaute. Nature 2014, 507, 258. [Google Scholar] [CrossRef]
- Blesa, A.; Baquedano, I.; Quintáns, N.G.; Mata, C.P.; Castón, J.R.; Berenguer, J. The transjugation machinery of Thermus thermophilus: Identification of TdtA, an ATPase involved in DNA donation. PLoS Genet. 2017, 13, e1006669. [Google Scholar] [CrossRef]
- Rankin, D.J.; Rocha, E.P.; Brown, S.P. What traits are carried on mobile genetic elements, and why? Heredity 2011, 106, 1. [Google Scholar] [CrossRef] [PubMed]
- Blount, Z.D.; Grogan, D.W. New insertion sequences of Sulfolobus: Functional properties and implications for genome evolution in hyperthermophilic archaea. Mol. Microbiol. 2005, 55, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Baliga, N.S.; Bonneau, R.; Facciotti, M.T.; Pan, M.; Glusman, G.; Deutsch, E.W.; Shannon, P.; Chiu, Y.; Weng, R.S.; Gan, R.R.; et al. Genome sequence of Haloarcula marismortui: A halophilic archaeon from the Dead Sea. Genome Res. 2004, 14, 2221–2234. [Google Scholar] [CrossRef]
- Takami, H.; Han, C.G.; Takaki, Y.; Ohtsubo, E. Identification and distribution of new insertion sequences in the genome of alkaliphilic Bacillus halodurans C-125. J. Bacteriol. 2001, 183, 4345–4356. [Google Scholar] [CrossRef] [PubMed]
- Brazelton, W.J.; Baross, J.A. Abundant transposases encoded by the metagenome of a hydrothermal chimney biofilm. ISME J. 2009, 3, 1420. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NAR1 | HB27 | HB8 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ch | M1 | M2 | Ch | pTT27 | Ch | pTT27 | pVV8 | |||||||||
| C | P | C | P | C | P | C | P | C | P | C | P | C | P | C | P | |
| ISTth1 | 1 | 1 | 1 | |||||||||||||
| ISTth2 | 2 | 1 | ||||||||||||||
| ISTth3 | 1 | 1 | 2 | 1 | 1 | 1 | ||||||||||
| ISTth4 | 5 | 11 | 4 | 4 | 2 | |||||||||||
| ISTth5 | 1 | 1 | ||||||||||||||
| ISTth6 | 1 | 1 | 3 | 1 | 1 | 1 | 1 | 1 | ||||||||
| ISTth7 | 13 | 5 | 2 | 2 | 2 | 4 | 1 | 6 | 2 | 1 | 2 | |||||
| ISTth8 | 3 | 1 | 1 | 2 | ||||||||||||
| IS1000A/B* | 2 | 1 | 8 | 1 | 34 | 1 | 14 | 3 | 39 | |||||||
| Name | Family | Sub-Group | Size Range (bp) | DRs (bp) | N° ORF | Frequency in Thermus spp. |
|---|---|---|---|---|---|---|
| ISTth1 | IS3 | IS150 | 1200–1600 | 3–4 | 2 | + |
| ISTth2 | IS4 | IS10 | 1200–1350 | 9 | 1 | + |
| ISTth3 | IS1634 | IS4 | 1500–2000 | 5–6 | 1 | +++ |
| ISTth4 | IS256 | - | 1200–1500 | 8–9 | 1 | +++ |
| ISTth5 | IS256 | 1200–1500 | 0 | 1 | + | |
| ISTth6 | IS630 | - | 1000–1400 | 2 | 1–2 | ++ |
| ISTth7 | IS5 | ISH1 | 900–1150 | 8 | 1 | +++ |
| ISTth8 | IS701 | - | 1400–1550 | 4 | 1 | ++ |
| IS1000A | IS110 | - | 1136–1558 | 2 | 1 | +++ |
| IS1000B | IS110 | 954–1558 | 0 | 1 | ++ | |
| IS421 | IS4 | IS231 | 1450–5400 | 10–12 | 1 | + |
| Start | End | Found in Reference | Found in HB27A | Found in HB27E |
|---|---|---|---|---|
| 259,293 | 259,789 | NO | YES | YES |
| 386,772 | 389,127 | NO | YES | YES |
| 562,354 | 562,716 | NO | YES | YES |
| 1,082,351 | 1,082,693 | NO | NO | YES |
| 1,133,885 | 1,134,345 | NO | YES | YES |
| 1,370,579 | 1,371,607 | YES | YES | YES |
| 1,533,343 | 1,534,271 | YES | YES | YES |
| 1,550,847 | 1,551,323 | NO | YES | YES |
| 1,655,564 | 1,655,957 | NO | YES | YES |
| Start | End | Found in Reference | Found in HB27A | Found in HB27E |
|---|---|---|---|---|
| 23,402 | 23,834 | NO | NO | YES |
| 57,597 | 58,625 | YES | YES | YES |
| 72,589 | 73,617 | YES | YES | YES |
| 85,677 | 86,705 | YES | YES | YES |
| 154,975 | 155,865 | NO | YES | YES |
| 159,958 | 160,986 | YES | YES | YES |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blesa, A.; Sánchez, M.; Sacristán-Horcajada, E.; González-de la Fuente, S.; Peiró, R.; Berenguer, J. Into the Thermus Mobilome: Presence, Diversity and Recent Activities of Insertion Sequences Across Thermus spp. Microorganisms 2019, 7, 25. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7010025
Blesa A, Sánchez M, Sacristán-Horcajada E, González-de la Fuente S, Peiró R, Berenguer J. Into the Thermus Mobilome: Presence, Diversity and Recent Activities of Insertion Sequences Across Thermus spp. Microorganisms. 2019; 7(1):25. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7010025
Chicago/Turabian StyleBlesa, Alba, Mercedes Sánchez, Eva Sacristán-Horcajada, Sandra González-de la Fuente, Ramón Peiró, and José Berenguer. 2019. "Into the Thermus Mobilome: Presence, Diversity and Recent Activities of Insertion Sequences Across Thermus spp." Microorganisms 7, no. 1: 25. https://0-doi-org.brum.beds.ac.uk/10.3390/microorganisms7010025